Geographical Distribution and Selection of European Honey Bees Resistant to Varroa destructor

 , , ,
, , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Review of Scientific Research
2.1. Why Should We Select Bees Resistant to the Mite?
2.2. Naturally Selected Populations and Their Known Mechanisms
2.2.1. Apis cerana
2.2.2. Africanized Honey Bees
2.2.3. African Honey Bees
2.2.4. A. m. capensis
2.2.5. Surviving Honey Bees Populations from France
2.2.6. Surviving Honey Bees from Norway
2.2.7. Surviving Honey Bees from the USA
2.2.8. Surviving Honey Bees from Russia
2.2.9. The Case of Wild Honey Bees in Europe
2.3. Artificial Selection
2.3.1. Gotland Bees
2.3.2. Kefuss Bees
2.3.3. Blacquière Bees
2.4. Genetic Selection on Chosen Characters
2.4.1. Hygienic Behavior
2.4.2. Suppressed Mite Reproduction (SMR)
2.4.3. Varroa Sensitive Hygiene (VSH)
2.4.4. Uncapping-Recapping of Varroa Infested Brood Cells
2.4.5. Mite Non-Reproduction (MNR)
2.4.6. Grooming Behavior
2.4.7. Attractiveness of the Brood
2.4.8. Mite Population Dynamics
2.4.9. Postcapping Stage
2.4.10. Brood Cell Size
2.4.11. Varroa Versus Virus Selection?
2.4.12. Genomic Analysis of Varroa Resistant Bees
2.5. Conclusions from the Literature
3. Survey on the Presence of Naturally Selected Resistant Honey Bee Populations and the State of Selection Programs on Varroa Resistance in Across the EU
4. Interviews with Experts in the Field (Beekeepers, Breeders, Researchers) to Obtain Information on Practical Experience with Selection for Varroa Resistant Bees
4.1. Naturally Selected Populations
4.2. Deliberately Genetically Selected Populations
5. Conclusions about Varroa Resistant Honey Bees in Europe
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Milani, N. The resistance of Varroa jacobsoni Oud. to acaricides. Apidologie 1999, 30, 229–234. [Google Scholar] [CrossRef]
- Büchler, R.; Berg, S.; Le Conte, Y. Breeding for resistance to Varroa destructor in Europe. Apidologie 2010, 41, 393–408. [Google Scholar] [CrossRef] [Green Version]
- Rinderer, T.E.; Harris, J.W.; Hunt, G.J.; de Guzman, L.I. Breeding for resistance to Varroa destructor in North America. Apidologie 2010, 41, 409–424. [Google Scholar] [CrossRef] [Green Version]
- Dietemann, V.; Pflugfelder, J.; Anderson, D.; Charriere, J.-D.; Chejanovsky, N.; Dainat, B.; de Miranda, J.; Delaplane, K.; Dillier, F.-X.; Fuch, S.; et al. Varroa destructor: Research avenues towards sustainable control. J. Apic. Res. 2012, 51, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Mondet, F.; Beaurepaire, A.; McAfee, A.; Locke, B.; Alaux, C.; Blanchard, S.; Danka, B.; Le Conte, Y. Honey bee survival mechanisms against the parasite Varroa destructor: A systematic review of phenotypic and genomic research efforts. Int. J. Parasitol. 2020, 50, 433–447. [Google Scholar] [CrossRef] [PubMed]
- Locke, B. Natural Varroa mite-surviving Apis mellifera honeybee populations. Apidologie 2016, 47, 467–482. [Google Scholar] [CrossRef] [Green Version]
- Dietemann, V.; Nazzi, F.; Martin, S.J.; Anderson, D.L.; Locke, B.; Delaplane, K.S.; Wauquiez, Q.; Tannahill, C.; Frey, E.; Ziegelmann, B.; et al. Standard methods for varroa research. J. Apic. Res. 2013, 52. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, K.; Sakai, T. Possible combination of three control measures for Varroasis. In Proceedings of the XXXth International Apicultural Congress, Nagoya, Japan, 10–16 October 1985; pp. 202–206. [Google Scholar]
- Oudemans, A.C. On a new genus and species of parasitic acari. Notes Leyden Mus. 1904, 24, 216–222. [Google Scholar]
- Anderson, D.L.; Trueman, J.W.H. Varroa jacobsoni (Acari: Varroidae) is more than one species. Exp. Appl. Acarol. 2000, 24, 165–189. [Google Scholar] [CrossRef]
- Rath, W. Co-adaptation of Apis cerana Fabr. and Varroa jacobsoni Oud. Apidologie 1999, 30, 97–110. [Google Scholar] [CrossRef] [Green Version]
- Rath, W. The key to Varroa—The drones of Apis cerana and thier cell cap. Am. Bee J. 1992, 132, 329–331. [Google Scholar]
- Rosenkranz, P.; Tewarson, N.C.; Singh, A.; Engels, W. Differential hygienic behaviour towards Varroa jacobsoni in capped worker brood of Apis cerana depends on alien scent adhering to the mites. J. Apic. Res. 1993, 32, 89–93. [Google Scholar] [CrossRef]
- Hanel, H.; Koeniger, N. Possible regulation of the reproduction of the honey bee mite Varroa jacobsoni by a host’s hormone Juvenile Hormone III. J. Insect Physiol. 1986, 32, 791–798. [Google Scholar] [CrossRef]
- Rosenkranz, P.; Rachinsky, A.; Strambi, A.; Strambi, C.; Ropstorf, P. Juvenile-hormone titer in capped worker brood of Apis mellifera and reproduction in the bee mite Varroa jacobsoni. Gen. Comp. Endocrinol. 1990, 78, 189–193. [Google Scholar] [CrossRef]
- Page, P.; Lin, Z.; Buawangpong, N.; Zheng, H.; Hu, F.; Neumann, P.; Chantawannakul, P.; Dietemann, V. Social apoptosis in honey bee superorganisms. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Z.; Qin, Y.; Page, P.; Wang, S.; Li, L.; Wen, Z.; Hu, F.; Neumann, P.; Zheng, H.; Dietemann, V. Reproduction of parasitic mites Varroa destructor in original and new honeybee hosts. Ecol. Evol. 2018, 8, 2135–2145. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.S.; Fang, Y.; Xu, S.; Ge, L. The resistance Mecanism of the Asian Honey Bee, Apis cerana Fabr., to an Ectoparasitic Mite, Varroa jacobsoni Oudemans. J. Invertebr. Pathol. 1987, 49, 54–60. [Google Scholar] [CrossRef]
- Tewarson, N.C.; Singh, A.; Engels, W. Reproduction of Varroa jacobsoni in colonies of Apis cerana indica under natural and experimental conditions. Apidologie 1992, 23, 161–171. [Google Scholar] [CrossRef] [Green Version]
- Boecking, O.; Spivak, M. Behavioral defenses of honey bees against Varroa jacobsoni Oud. Apidologie 1999, 30, 141–158. [Google Scholar] [CrossRef] [Green Version]
- Dejong, D.; Goncalves, L.S. The Varroa Problem in Brazil. Am. Bee J. 1981, 121, 186–189. [Google Scholar]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and control of Varroa destructor. J. Invertebr. Pathol. 2010, 103, S96–S119. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, F.E.; Torres, R.R.; Strapazzon, R.; Ramirez, S.A.; Guerra, J.C.V., Jr.; Koling, D.F.; Moretto, G. Changes in the reproductive ability of the mite Varroa destructor (Anderson e Trueman) in Africanized honey bees (Apis mellifera L.) (Hymenoptera: Apidae) colonies in southern Brazil. Neotrop. Entomol. 2007, 36, 949–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schafaschek, T.P.; Hickel, E.R.; de Oliveira, C.A.L.; de Toled, V.A.A. Infestation and Reproduction of Varroa destructor Anderson and Trueman and Hygienic Behavior in Colonies of Apis mellifera L. (Africanized Honeybee) with Queens of Different Genetic Origins. Sociobiology 2019, 66, 448–456. [Google Scholar] [CrossRef] [Green Version]
- Reyes-Quintana, M.; Espinosa-Montano, L.G.; Prieto-Merlos, D.; Koleoglu, G.; Petukhova, T.; Correa-Benitez, A.; Guzman-Novoa, E. Impact of Varroa destructor and deformed wing virus on emergence, cellular immunity, wing integrity and survivorship of Africanized honey bees in Mexico. J. Invertebr. Pathol. 2019, 164, 43–48. [Google Scholar] [CrossRef]
- Message, D.; Goncalves, L.S. Effect of the size of worker brood cells of Africanized honey bees on infestation and reproduction of the Ectoparasitic mite Varroa jacobsoni Oud. Apidologie 1995, 26, 381–386. [Google Scholar] [CrossRef] [Green Version]
- Ritter, W.; De Jong, D. Reproduction of Varroa jacobsoni O. in Europe, the middle East and tropical South America. Z. Angew. Entomol. J. Appl. Entomol. 1984, 98, 55–57. [Google Scholar] [CrossRef]
- Vandame, R. Africanised bees tolerance to varroa in Mexico: Mite infertility is not the only factor. In Proceedings of the XXXVth International Apicultural Congress, Antwerp, Belgium, 1–6 September 1997. [Google Scholar]
- GuzmanNovoa, E.; Sanchez, A.; Page, R.E.; Garcia, T. Susceptibility of European and africanized honeybees (Apis mellifera L) and their hybrids to Varroa jacobsoni Oud. Apidologie 1996, 27, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Guzman-Novoa, E.; Vandame, R.; Arechavaleta, M.E. Susceptibility of European and Africanized honey bees (Apis mellifera L.) to Varroa jacobsoni Oud. in Mexico. Apidologie 1999, 30, 173–182. [Google Scholar] [CrossRef]
- Arechavaleta-Velasco, M.E.; Guzman-Novoa, E. Relative effect of four characteristics that restrain the population growth of the mite Varroa destructor in honey bee (Apis mellifera) colonies. Apidologie 2001, 32, 157–174. [Google Scholar] [CrossRef] [Green Version]
- Moretto, G.; Goncalves, L.S.; Dejong, D. Heritability of africanized and european honey-bee defensive behavior against the mite Varroa jacobsoni. Rev. Bras. Genet. 1993, 16, 71–77. [Google Scholar]
- Aumeier, P. Bioassay for grooming effectiveness towards Varroa destructor mites in Africanized and Carniolan honey bees. Apidologie 2001, 32, 81–90. [Google Scholar] [CrossRef] [Green Version]
- Guzman-Novoa, E.; Emsen, B.; Unger, P.; Espinosa-Montano, L.G.; Petukhova, T. Genotypic variability and relationships between mite infestation levels, mite damage, grooming intensity, and removal of Varroa destructor mites in selected strains of worker honey bees (Apis mellifera L.). J. Invertebr. Pathol. 2012, 110, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Matheson, A. World Bee Health Report. Bee World 1993, 74, 176–212. [Google Scholar] [CrossRef]
- Allsopp, M. Varroa jacobsoni in South Africa. S. Afr. Bee J. 1997, 69, 73–82. [Google Scholar]
- Pirk, C.W.W.; Strauss, U.; Yusuf, A.A.; Demares, F.; Human, H. Honeybee health in Africa-a review. Apidologie 2016, 47, 276–300. [Google Scholar] [CrossRef] [Green Version]
- Dietemann, V.; Pirk, C.W.W.; Crewe, R. Is there a need for conservation of honeybees in Africa? Apidologie 2009, 40, 285–295. [Google Scholar] [CrossRef] [Green Version]
- Pirk, C.W.W.; Human, H.; Crewe, R.M.; vanEngelsdorp, D. A survey of managed honey bee colony losses in the Republic of South Africa-2009 to 2011. J. Apic. Res. 2014, 53, 35–42. [Google Scholar] [CrossRef]
- Strauss, U.; Human, H.; Gauthier, L.; Crewe, R.M.; Dietemann, V.; Pirk, C.W.W. Seasonal prevalence of pathogens and parasites in the savannah honeybee (Apis mellifera scutellata). J. Invertebr. Pathol. 2013, 114, 45–52. [Google Scholar] [CrossRef] [Green Version]
- Strauss, U.; Pirk, C.W.W.; Crewe, R.M.; Human, H.; Dietemann, V. Impact of Varroa destructor on honeybee (Apis mellifera scutellata) colony development in South Africa. Exp. Appl. Acarol. 2015, 65, 89–106. [Google Scholar] [CrossRef] [Green Version]
- Nganso, B.T.; Fombong, A.T.; Yusuf, A.A.; Pirk, C.W.W.; Stuhl, C.; Torto, B. Low fertility, fecundity and numbers of mated female offspring explain the lower reproductive success of the parasitic mite Varroa destructor in African honeybees. Parasitology 2018, 145, 1633–1639. [Google Scholar] [CrossRef]
- Moritz, R.F.A. Heritability of the postcapping stage in Apis mellifera and its relation to varroatosis resistance. J. Hered. 1985, 76, 267–270. [Google Scholar] [CrossRef]
- Moritz, R.F.A.; Mautz, D. Development of Varroa-jacobsoni in colonies of Apis mellifera capensis and Apis Mellifera Carn. Apidologie 1990, 21, 53–58. [Google Scholar] [CrossRef] [Green Version]
- Allsopp, M. Analysis of Varroa destructor Infestation of Southern African Honey Bee Populations. Master’s Thesis, University of Pretoria, Pretoria, South Africa, 2006. [Google Scholar]
- Cheruiyot, S.K.; Lattorff, H.M.G.; Kahuthia-Gathu, R.; Mbugi, J.P.; Muli, E. Varroa-specific hygienic behavior of Apis mellifera scutellata in Kenya. Apidologie 2018, 49, 439–449. [Google Scholar] [CrossRef] [Green Version]
- Gebremedhn, H.; Amssalu, B.; De Smet, L.; de Graaf, D.C. Factors restraining the population growth of Varroa destructor in Ethiopian honey bees (Apis mellifera simensis). PLoS ONE 2019, 14, e0223236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calis, J.; Boot, W.J.; Beetsma, J. Reproductive success of the Varroa mite in honeybee worker brood with differential development times. In Proceedings of the Section Experimental and Applied Entomology of the Netherlands Entomological Society; Nederlandse Entomologische Verniging (NEV): Amsterdam, The Netherlands, 1996; pp. 89–94. [Google Scholar]
- Martin, S.J.; Kryger, P. Reproduction of Varroa destructor in South African honey bees: Does cell space influence Varroa male survivorship? Apidologie 2002, 33, 51–61. [Google Scholar] [CrossRef] [Green Version]
- Le Conte, Y.; De Vaublanc, G.; Crauser, D.; Jeanne, F.; Rousselle, J.C.; Becard, J.M. Honey bee colonies that have survived Varroa destructor. Apidologie 2007, 38, 566–572. [Google Scholar] [CrossRef] [Green Version]
- Fries, I.; Aarhus, A.; Hansen, H.; Korpela, S. Comparison of Diagnostic Methods for Detection of Low Infestation Levels of Varroa-jacobsoni in Honey-Bee (Apis-Mellifera) Colonies. Exp. Appl. Acarol. 1991, 10, 279–287. [Google Scholar] [CrossRef]
- Locke, B.; Le Conte, Y.; Crauser, D.; Fries, I. Host adaptations reduce the reproductive success of Varroa destructor in two distinct European honey bee populations. Ecol. Evol. 2012, 2, 1144–1150. [Google Scholar] [CrossRef]
- Navajas, M.; Migeon, A.; Alaux, C.; Martin-Magniette, M.L.; Robinson, G.E.; Evans, J.D.; Cros-Arteil, S.; Crauser, D.; Le Conte, Y. Differential gene expression of the honey bee Apis mellifera associated with Varroa destructor infection. BMC Genom. 2008, 9, 301. [Google Scholar] [CrossRef] [Green Version]
- Navajas, M.; Le Conte, Y.; Solignac, M.; Cros-Arteil, S.; Cornuet, J.M. The complete sequence of the mitochondrial genome of the honeybee ectoparasite mite Varroa destructor (Acari: Mesostigmata). Mol. Biol. Evol. 2002, 19, 2313–2317. [Google Scholar] [CrossRef] [Green Version]
- Solignac, M.; Cornuet, J.M.; Vautrin, D.; Le Conte, Y.; Anderson, D.; Evans, J.; Cros-Arteil, S.; Navajas, M. The invasive Korea and Japan types of Varroa destructor, ectoparasitic mites of the Western honeybee (Apis mellifera), are two partly isolated clones. Proc. R. Soc. B Biol. Sci. 2005, 272, 411–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Solignac, M.; Vautrin, D.; Pizzo, A.; Navajas, M.; Le Conte, Y.; Cornuet, J.M. Characterization of microsatellite markers for the apicultural pest Varroa destructor (Acari: Varroidae) and its relatives. Mol. Ecol. Notes 2003, 3, 556–559. [Google Scholar] [CrossRef]
- Liu, Z.Q.; Tan, J.G.; Huang, Z.Y.; Dong, K. Effect of a fluvalinate-resistance-associated sodium channel mutation from varroa mites on cockroach sodium channel sensitivity to fluvalinate, a pyrethroid insecticide. Insect Biochem. Mol. Biol. 2006, 36, 885–889. [Google Scholar] [CrossRef] [PubMed]
- Beaurepaire, A.L.; Moro, A.; Mondet, F.; Le Conte, Y.; Neumann, P.; Locke, B. Population genetics of ectoparasitic mites suggest arms race with honeybee hosts. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Sumpter, D.J.T.; Martin, S.J. The dynamics of virus epidemics in Varroa-infested honey bee colonies. J. Anim. Ecol. 2004, 73, 51–63. [Google Scholar] [CrossRef]
- Oddie, M.A.Y.; Dahle, B.; Neumann, P. Norwegian honey bees surviving Varroa destructor mite infestations by means of natural selection. Peerj 2017, 5. [Google Scholar] [CrossRef] [Green Version]
- Oddie, M.A.Y.; Dahle, B.; Neumann, P. Reduced Postcapping Period in Honey Bees Surviving Varroa destructor by Means of Natural Selection. Insects 2018, 9, 149. [Google Scholar] [CrossRef] [Green Version]
- Oddie, M.A.Y.; Neumann, P.; Dahle, B. Cell size and Varroa destructor mite infestations in susceptible and naturally-surviving honeybee (Apis mellifera) colonies. Apidologie 2019, 50, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Oddie, M.; Büchler, R.; Dahle, B.; Kovacic, M.; Le Conte, Y.; Locke, B.; de Miranda, J.R.; Mondet, F.; Neumann, P. Rapid parallel evolution overcomes global honey bee parasite. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [Green Version]
- Seeley, T.D. Honey bees of the Arnot Forest: A population of feral colonies persisting with Varroa destructor in the northeastern United States. Apidologie 2007, 38, 19–29. [Google Scholar] [CrossRef]
- Loftus, J.C.; Smith, M.L.; Seeley, T.D. How Honey Bee Colonies Survive in the Wild: Testing the Importance of Small Nests and Frequent Swarming. PLoS ONE 2016, 11, e0150362. [Google Scholar] [CrossRef] [PubMed]
- Seeley, T.D. Life-history traits of wild honey bee colonies living in forests around Ithaca, NY, USA. Apidologie 2017, 48, 743–754. [Google Scholar] [CrossRef] [Green Version]
- Rinderer, T.E.; Kuznetsov, V.N.; Danka, R.G.; Delatte, G.T. An importation of potentially Varroa-resistant honey bees from far-eastern Russia. Am. Bee J. 1997, 137, 787–789. [Google Scholar]
- Tarpy, D.R.; Summers, J.; Keller, J.J. Comparison of parasitic mites in Russian-Hybrid and Italian honey bee (Hymenoptera: Apidae) colonies across three different locations in north Carolina. J. Econ. Entomol. 2007, 100, 258–266. [Google Scholar] [CrossRef]
- Bourgeois, A.L.; Rinderer, T.E. Genetic Characterization of Russian Honey Bee Stock Selected for Improved Resistance to Varroa destructor. J. Econ. Entomol. 2009, 102, 1233–1238. [Google Scholar] [CrossRef] [PubMed]
- de Guzman, L.I.; Rinderer, T.E.; Frake, A.M. Comparative reproduction of Varroa destructor in different types of Russian and Italian honey bee combs. Exp. Appl. Acarol. 2008, 44, 227–238. [Google Scholar] [CrossRef]
- Kirrane, M.J.; de Guzman, L.I.; Whelan, P.M.; Frake, A.M.; Rinderer, T.E. Evaluations of the Removal of Varroa destructor in Russian Honey Bee Colonies that Display Different Levels of Varroa Sensitive Hygienic Activities. J. Insect Behav. 2018, 31, 283–297. [Google Scholar] [CrossRef]
- Kohl, P.L.; Rutschmann, B. The neglected bee trees: European beech forests as a home for feral honey bee colonies. Peerj 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Dynes, T.L.; Berry, J.A.; Delaplane, K.S.; de Roode, J.C.; Brosi, B.J. Assessing virulence of Varroa destructor mites from different honey bee management regimes. Apidologie 2020, 51, 276–289. [Google Scholar] [CrossRef] [Green Version]
- Kefuss, J.; Vanpoucke, J.; De Lahitte, J.D.; Ritter, W. Varroa tolerance in France of intermissa bees from Tunisia and their naturally mated descendants: 1993–2004. Am. Bee J. 2004, 144, 563–568. [Google Scholar]
- Fries, I.; Imdorf, A.; Rosenkranz, P. Survival of mite infested (Varroa destructor) honey bee (Apis mellifera) colonies in a Nordic climate. Apidologie 2006, 37, 564–570. [Google Scholar] [CrossRef] [Green Version]
- Blacquiere, T.; Boot, W.; Calis, J.; Moro, A.; Neumann, P.; Panziera, D. Darwinian black box selection for resistance to settled invasive Varroa destructor parasites in honey bees. Biol. Invasions 2019, 21, 2519–2528. [Google Scholar] [CrossRef] [Green Version]
- Fries, I.; Hansen, H.; Imdorf, A.; Rosenkranz, P. Swarming in honey bees (Apis mellifera) and Varroa destructor population development in Sweden. Apidologie 2003, 34, 389–397. [Google Scholar] [CrossRef] [Green Version]
- Fries, I.; Bommarco, R. Possible host-parasite adaptations in honey bees infested by Varroa destructor mites. Apidologie 2007, 38, 525–533. [Google Scholar] [CrossRef]
- Locke, B.; Fries, I. Characteristics of honey bee colonies (Apis mellifera) in Sweden surviving Varroa destructor infestation. Apidologie 2011, 42, 533–542. [Google Scholar] [CrossRef] [Green Version]
- Thaduri, S.; Locke, B.; Granberg, F.; de Miranda, J.R. Temporal changes in the viromes of Swedish Varroa-resistant and Varroa-susceptible honeybee populations. PLoS ONE 2018, 13, e0206938. [Google Scholar] [CrossRef]
- Ritter, W. Development of varroa mite population in treated and untreated colonies in Tunisia. Apidologie 1990, 21, 368–370. [Google Scholar]
- Kefuss, J.; Vanpoucke, J.; Bolt, M.; Kefuss, C. Selection for resistance to Varroa destructor under commercial beekeeping conditions. J. Apic. Res. 2015, 54, 563–576. [Google Scholar] [CrossRef] [Green Version]
- Conlon, B.H.; Aurori, A.; Giurgiu, A.-I.; Kefuss, J.; Dezmirean, D.S.; Moritz, R.F.A.; Routtu, J. A gene for resistance to the Varroa mite (Acari) in honey bee (Apis mellifera) pupae. Mol. Ecol. 2019, 28, 2958–2966. [Google Scholar] [CrossRef]
- Kruitwagen, A.; van Langevelde, F.; van Dooremalen, C.; Blacquiere, T. Naturally selected honey bee (Apis mellifera) colonies resistant to Varroa destructor do not groom more intensively. J. Apic. Res. 2017, 56, 354–365. [Google Scholar] [CrossRef] [Green Version]
- Panziera, D.; van Langevelde, F.; Blacquiere, T. Varroa sensitive hygiene contributes to naturally selected varroa resistance in honey bees. J. Apic. Res. 2017, 56, 635–642. [Google Scholar] [CrossRef] [Green Version]
- Blacquière, T.; Panziera, D. A Plea for Use of Honey Bees’ Natural Resilience in Beekeeping. Bee World 2018, 95, 34–38. [Google Scholar] [CrossRef] [Green Version]
- Büchler, R. Varroa Tolerance in Honey Bees—Occurrence, Characters and Breeding. Bee World 1994, 75, 54–70. [Google Scholar] [CrossRef]
- Mondet, F.; Kim, S.H.; de Miranda, J.R.; Beslay, D.; Le Conte, Y.; Mercer, A.R. Specific Cues Associated With Honey Bee Social Defence against Varroa destructor Infested Brood. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef]
- Winston, M.L. The Biology of the Honey Bee; Havard University Press: Cambridge, UK, 1987. [Google Scholar]
- Morfin, N.; Espinosa-Montano, L.G.; Guzman-Novoa, E. A direct assay to assess self-grooming behavior in honey bees (Apis mellifera L.). Apidologie 2020, 51, 892–897. [Google Scholar] [CrossRef]
- Kraus, B.; Velthuis, H.H.W. High humidity in the honey bee (Apis mellifera L) brood nest limits reproduction of the parasitic mite Varroa jacobsoni Oud. Naturwissenschaften 1997, 84, 217–218. [Google Scholar] [CrossRef] [Green Version]
- Villa, J.D.; Danka, R.G.; Harris, J.W. Simplified methods of evaluating colonies for levels of Varroa Sensitive Hygiene (VSH). J. Apic. Res. 2009, 48, 162–167. [Google Scholar] [CrossRef]
- Harbo, J.R.; Harris, J.W. Suppressed mite reproduction explained by the behaviour of adult bees. J. Apic. Res. 2005, 44, 21–23. [Google Scholar] [CrossRef]
- Büchler, R.; Andonov, S.; Bienefeld, K.; Costa, C.; Hatjina, F.; Kezic, N.; Kryger, P.; Spivak, M.; Uzunov, A.; Wilde, J. Standard methods for rearing and selection of Apis mellifera queens. J. Apic. Res. 2013, 52. [Google Scholar] [CrossRef] [Green Version]
- Rothenbuhler, W.C. Behaviour genetics of nest cleaning in honey bees. I. Responses of 4 inbred lines to disease-killd brood. Anim. Behav. 1964, 12, 578–583. [Google Scholar] [CrossRef]
- Momot, J.P.; Rothenbuhler, W.C. Behaviour Genetics of Nest Cleaning in Honeybees. VI. Interactions of Age and Genotype of Bees, and Nectar Flow. J. Apic. Res. 1971, 10, 11–21. [Google Scholar] [CrossRef]
- Spivak, M.; Gilliam, M. Hygienic behaviour of honey bees and its application for control of brood diseases and varroa. Part I. Hygienic behaviour and resistance to American foulbrood. Bee World 1998, 79, 124–134. [Google Scholar] [CrossRef]
- Newton, D.C.; Ostasiewski, N.J. A simplified bioassay for behavioral resistance to American foulbrood in honey bees (Apis mellifera L.). Am. Bee J. 1986, 126, 278–281. [Google Scholar]
- Spivak, M.; Downey, D.L. Field assays for hygienic behavior in honey bees (Hymenoptera: Apidae). J. Econ. Entomol. 1998, 91, 64–70. [Google Scholar] [CrossRef] [Green Version]
- Espinosa-Montano, L.G.; Guzman-Novoa, E.; Sanchez-Albarran, A.; Montaldo, H.H.; Correa-Benitez, A. Comparative study of three assays to evaluate hygienic behavior in honey bee (Apis mellifera L.) colonies. Vet. Mex. 2008, 39, 39–54. [Google Scholar]
- Hooffmann, S. Untersuchungsmethoden und Analyse der quantitativ genetischen Basis unterschiedlicher Varroatose-Anfälligkeit von Bienenvölkern der Carnica-Rasse (Apis mellifera carnica Pollmann). Ph.D. Thesis, Rheinische Friedrich-Wilhelms-Universität, Bonn, Germany, 1996. [Google Scholar]
- Facchini, E.; Bijma, P.; Pagnacco, G.; Rizzi, R.; Brascamp, E.W. Hygienic behaviour in honeybees: A comparison of two recording methods and estimation of genetic parameters. Apidologie 2019, 50, 163–172. [Google Scholar] [CrossRef] [Green Version]
- Spivak, M. Honey bee hygienic behavior and defense against Varroa jacobsoni. Apidologie 1996, 27, 245–260. [Google Scholar] [CrossRef] [Green Version]
- Thakur, R.K.; Bienefeld, K.; Keller, R. Varroa defense behavior in A. mellifera carnica. Am. Bee J. 1997, 137, 143–148. [Google Scholar]
- Masterman, R.; Ross, R.; Mesce, K.; Spivak, M. Olfactory and behavioral response thresholds to odors of diseased brood differ between hygienic and non-hygienic honey bees (Apis mellifera L.). J. Comp. Physiol. A Sens. Neural Behav. Physiol. 2001, 187, 441–452. [Google Scholar]
- Swanson, J.A.I.; Torto, B.; Kells, S.A.; Mesce, K.A.; Tumlinson, J.H.; Spivak, M. Odorants that Induce Hygienic Behavior in Honeybees: Identification of Volatile Compounds in Chalkbrood-Infected Honeybee Larvae. J. Chem. Ecol. 2009, 35, 1108–1116. [Google Scholar] [CrossRef]
- Guarna, M.M.; Melathopoulos, A.P.; Huxter, E.; Iovinella, I.; Parker, R.; Stoynov, N.; Tam, A.; Moon, K.-M.; Chan, Q.W.T.; Pelosi, P.; et al. A search for protein biomarkers links olfactory signal transduction to social immunity. BMC Genom. 2015, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondet, F.; Alaux, C.; Severac, D.; Rohmer, M.; Mercer, A.R.; Le Conte, Y. Antennae hold a key to Varroa-sensitive hygiene behaviour in honey bees. Sci. Rep. 2015, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spivak, M.; Reuter, G.S. Honey bee hygienic behavior. Am. Bee J. 1998, 138, 283–286. [Google Scholar]
- Ibrahim, A.; Reuter, G.S.; Spivak, M. Field trial of honey bee colonies bred for mechanisms of resistance against Varroa destructor. Apidologie 2007, 38, 67–76. [Google Scholar] [CrossRef] [Green Version]
- Palacio, M.A.; Figini, E.E.; Ruffinengo, S.R.; Rodriguez, E.M.; del Hoyo, M.L.; Bedascarrasbure, E.L. Changes in a population of Apis mellifera L. selected for hygienic behaviour and its relation to brood disease tolerance. Apidologie 2000, 31, 471–478. [Google Scholar] [CrossRef]
- Bigio, G.; Al Toufailia, H.; Hughes, W.O.H.; Ratnieks, F.L.W. The effect of one generation of controlled mating on the expression of hygienic behaviour in honey bees. J. Apic. Res. 2014, 53, 563–568. [Google Scholar] [CrossRef] [Green Version]
- Harbo, J.R.; Harris, J.W. Selecting honey bees for resistance to Varroa jacobsoni. Apidologie 1999, 30, 183–196. [Google Scholar] [CrossRef]
- Harris, J.W.; Harbo, J.R.; Villa, J.D.; Danka, R.G. Variable population growth of Varroa destructor (Mesostigmata: Varroidae) in colonies of honey bees (Hymenoptera: Apidae) during a 10-year period. Environ. Entomol. 2003, 32, 1305–1312. [Google Scholar] [CrossRef]
- Harbo, J.R.; Hoopingarner, R.A. Resistance to varroa expressed by honey bees in the USA. Am. Bee J. 1995, 135, 827. [Google Scholar]
- Harris, J.W.; Harbo, J.R. Changes in reproduction of Varroa destructor after honey bee queens were exchanged between resistant and susceptible colonies. Apidologie 2000, 31, 689–699. [Google Scholar] [CrossRef]
- Harbo, J.R.; Harris, J.W. Heritability in honey bees (Hymenoptera: Apidae) of characteristics associated with resistance to Varroa jacobsoni (Mesostigmata: Varroidae). J. Econ. Entomol. 1999, 92, 261–265. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, A.; Spivak, M. The relationship between hygienic behavior and suppression of mite reproduction as honey bee (Apis mellifera) mechanisms of resistance to Varroa destructor. Apidologie 2006, 37, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.W.; Danka, R.G.; Villa, J.D. Honey Bees (Hymenoptera: Apidae) With the Trait of Varroa Sensitive Hygiene Remove Brood With All Reproductive Stages of Varroa Mites (Mesostigmata: Varroidae). Ann. Entomol. Soc. Am. 2010, 103, 146–152. [Google Scholar] [CrossRef]
- Harris, J.W. Bees with Varroa Sensitive Hygiene preferentially remove mite infested pupae aged ≤ five days post capping. J. Apic. Res. 2007, 46, 134–139. [Google Scholar] [CrossRef]
- Villa, J.D.; Danka, R.G.; Harris, J.W. Repeatability of measurements of removal of mite-infested brood to assess Varroa Sensitive Hygiene. J. Apic. Res. 2018, 57, 329. [Google Scholar] [CrossRef]
- Danka, R.G.; Harris, J.W.; Villa, J.D. Expression of Varroa Sensitive Hygiene (VSH) in Commercial VSH Honey Bees (Hymenoptera: Apidae). J. Econ. Entomol. 2011, 104, 745–749. [Google Scholar] [CrossRef] [PubMed]
- Rinderer, T.E.; Danka, R.G.; Johnson, S.; Bourgeois, A.L.; Frake, A.M.; Villa, J.D.; De Guzman, L.I.; Harris, J.W. Functionality of Varroa-Resistant Honey Bees (Hymenoptera: Apidae) When Used for Western US Honey Production and Almond Pollination. J. Econ. Entomol. 2014, 107, 523–530. [Google Scholar] [CrossRef] [Green Version]
- Danka, R.G.; Harris, J.W.; Dodds, G.E. Selection of VSH- derived "Pol-line" honey bees and evaluation of their Varroa -resistance characteristics. Apidologie 2016, 47, 483–490. [Google Scholar] [CrossRef]
- Correa-Marques, M.H.; De Jong, D. Uncapping of worker bee brood, a component of the hygienic behavior of Africanized honey bees against the mite Varroa jacobsoni Oudemans. Apidologie 1998, 29, 283–289. [Google Scholar] [CrossRef] [Green Version]
- Kirrane, M.J.; de Guzman, L.I.; Holloway, B.; Frake, A.M.; Rinderer, T.E.; Whelan, P.M. Phenotypic and Genetic Analyses of the Varroa Sensitive Hygienic Trait in Russian Honey Bee (Hymenoptera: Apidae) Colonies. PLoS ONE 2015, 10, e0116672. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.J.; Hawkins, G.P.; Brettell, L.E.; Reece, N.; Correia-Oliveira, M.E.; Allsopp, M.H. Varroa destructor reproduction and cell re-capping in mite-resistant Apis mellifera populations. Apidologie 2020, 51, 369–381. [Google Scholar] [CrossRef] [Green Version]
- Buchegger, M.; Buechler, R.; Fuerst-Waltl, B.; Kovacic, M.; Willam, A. Relationships between resistance characteristics of honey bees (Apis mellifera) against Varroa mites (Varroa destructor). J. Cent. Eur. Agric. 2018, 19, 954–958. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.W.; Danka, R.G.; Villa, J.D. Changes in Infestation, Cell Cap Condition, and Reproductive Status of Varroa destructor (Mesostigmata: Varroidae) in Brood Exposed to Honey Bees With Varroa Sensitive Hygiene. Ann. Entomol. Soc. Am. 2012, 105, 512–518. [Google Scholar] [CrossRef]
- Kovačić, M.; Puškadija, Z.; Dražić, M.M.; Uzunov, A.; Meixner, M.D.; Büchler, R. Effects of selection and local adaptation on resilience and economic suitability in Apis mellifera carnica. Apidologie 2020, 51, 1062–1073. [Google Scholar] [CrossRef]
- Büchler, R.; Kovačić, M.; Buchegger, M.; Puškadija, Z.; Hoppe, A.; Brascamp, E.W. Evaluation of Traits for the Selection of Apis mellifera for Resistance against Varroa destructor. Insects 2020, 11, 618. [Google Scholar] [CrossRef] [PubMed]
- Noel, A.; Le Conte, Y.; Mondet, F. Varroa destructor: How does it harm Apis mellifera honey bees and what can be done about it? Emerg. Top. Life Sci. 2020, 4, 45–57. [Google Scholar] [CrossRef] [PubMed]
- Ruttner, F.; Hänel, H. Active defence against Varroa mites in a Carniolan strain of honey bee (Apis mellifera carnica Pollman). Apidologie 1992, 23, 173–187. [Google Scholar] [CrossRef]
- Büchler, R.; Drescher, W.; Tornier, I. Grooming behaviour of Apis cerana, Apis mellifera and Apis dorsata and its effects on the parasitic mites Varroa jacobsoni and Tropilaelaps clareae. Exp. Appl. Acarol. 1992, 16, 313–319. [Google Scholar] [CrossRef]
- Bozic, J.; Valentincic, T. Quantitative analysis of social grooming behavior of the honey bee Apis mellifera carnica. Apidologie 1995, 26, 141–147. [Google Scholar] [CrossRef] [Green Version]
- Rivera-Marchand, B.; Oskay, D.; Giray, T. Gentle Africanized bees on an oceanic island. Evol. Appl. 2012, 5, 746–756. [Google Scholar] [CrossRef]
- Fries, I.; Wei, H.Z.; Shi, W.; Chen, S.J. Grooming behavior and damaged mites (Varroa jacobsoni) in Apis cerana cerana and Apis mellifera ligustica. Apidologie 1996, 27, 3–11. [Google Scholar] [CrossRef]
- Webster, T.C.; Thacker, E.M.; Vorisek, F.E. Live Varroa jacobsoni (Mesostigmata: Varroidae) fallen from honey bee (Hymenoptera: Apidae) colonies. J. Econ. Entomol. 2000, 93, 1596–1601. [Google Scholar] [CrossRef] [PubMed]
- Rosenkranz, P.; Fries, I.; Boecking, O.; Sturmer, M. Damaged Varroa mites in the debris of honey bee (Apis mellifera L) colonies with and without hatching brood. Apidologie 1997, 28, 427–437. [Google Scholar] [CrossRef] [Green Version]
- Bienefeld, K.; Zautke, F.; Pronin, D.; Mazeed, A. Recording the proportion of damaged Varroa jacobsoni Oud. in the debris of honey bee colonies (Apis mellifera). Apidologie 1999, 30, 249–256. [Google Scholar] [CrossRef] [Green Version]
- Szabo, T.I.; Walker, C.R.T. Damage to dead Varroa jacobsoni caused by the larvae of Galleria mellonella. Am. Bee J. 1995, 135, 421–422. [Google Scholar]
- Davis, A.R. Regular dorsal dimples on Varroa destructor—Damage symptoms or developmental origin? Apidologie 2009, 40, 151–162. [Google Scholar] [CrossRef] [Green Version]
- Kovacic, M.; Puskadija, Z.; Drazic, M.M. Grooming behavior in relation to varroa (Varroa destructor) infestation level of Carniolan honey bee colonies (Apis mellifera carnica). J. Cent. Eur. Agric. 2018, 19, 959–964. [Google Scholar] [CrossRef]
- Dadoun, N.; Nait-Mouloud, M.; Mohammedi, A.; Sadeddine Zennouche, O. Differences in grooming behavior between susceptible and resistant honey bee colonies after 13 years of natural selection. Apidologie 2020. [Google Scholar] [CrossRef]
- Büchler, R. Design and success of a German breeding program for Varroa tolerance. Am. Bee J. 2000, 140, 662–665. [Google Scholar]
- Ehrhardt, K.; Reinsh, N.; Büchler, R.; Garrido, C.; Bienefeld, K. Genetic parameters for varroa resistance in the honey bee. In Proceedings of the 40th International Apicultural Congress, Melbourne, Australia, 9–14 September 2007; pp. 145–146. [Google Scholar]
- Hamiduzzaman, M.M.; Emsen, B.; Hunt, G.J.; Subramanyam, S.; Williams, C.E.; Tsuruda, J.M.; Guzman-Novoa, E. Differential Gene Expression Associated with Honey Bee Grooming Behavior in Response to Varroa Mites. Behav. Genet. 2017, 47, 335–344. [Google Scholar] [CrossRef] [Green Version]
- Arechavaleta-Velasco, M.E.; Alcala-Escamilla, K.; Robles-Rios, C.; Tsuruda, J.M.; Hunt, G.J. Fine-Scale Linkage Mapping Reveals a Small Set of Candidate Genes Influencing Honey Bee Grooming Behavior in Response to Varroa Mites. PLoS ONE 2012, 7, e47269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Currie, R.W.; Tahmasbi, G.H. The ability of high- and low-grooming lines of honey bees to remove the parasitic mite Varroa destructor is affected by environmental conditions. Can. J. Zool. 2008, 86, 1059–1067. [Google Scholar] [CrossRef]
- Andino, G.K.; Hunt, G.J. A scientific note on a new assay to measure honeybee mite-grooming behavior. Apidologie 2011, 42, 481–484. [Google Scholar] [CrossRef] [Green Version]
- Le Conte, Y.; Arnold, G.; Trouiller, J.; Masson, C.; Chappe, B.; Ourisson, G. Attraction of the parasitic mite Varroa to the drone larvae of honeybees by simple aliphatic esters. Science 1989, 245, 638–639. [Google Scholar] [CrossRef] [PubMed]
- Nazzi, F.; Le Conte, Y. Ecology of Varroa destructor, the Major Ectoparasite of the Western Honey Bee, Apis mellifera. In Annual Review of Entomology; Berenbaum, M.R., Ed.; Annual Reviews: Palo Alto, CA, USA, 2016; Volume 61, pp. 417–432. [Google Scholar]
- Trouiller, J.; Arnold, G.; Le Conte, Y.; Masson, C.; Chappe, B. Temporal pheromonal and kairomonal secretion in the brood of honeybees. Naturwissenschaften 1991, 78, 368–370. [Google Scholar] [CrossRef]
- Aumeier, P.; Rosenkranz, P. Brood attractivity and Varroa infestation: A comparison of Africanized and European bees. Apidologie 1997, 28, 182–184. [Google Scholar]
- Aumeier, P.; Rosenkranz, P.; Gonçalves, L.S. Defense mechanisms of honey bees against varroosis and brood diseases: Comparison between Apis mellifera carnica and africanized bees in Brazil. Apidologie 1996, 27, 286–288. [Google Scholar]
- Fuchs, S. Untersuchungen zur quantitativen Abschätzung des Befalls von Bienenvölkern mit Varroa jacobsoni Oudemans und zur Verteilung des Parasiten im Bienenvolk. Apidologie 1985, 16, 343–368. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.V.; Moon, R.D.; Burkness, E.C.; Hutchison, W.D.; Spivak, M. Practical Sampling Plans for Varroa destructor (Acari: Varroidae) in Apis mellifera (Hymenoptera: Apidae) Colonies and Apiaries. J. Econ. Entomol. 2010, 103, 1039–1050. [Google Scholar] [CrossRef]
- Moritz, F.A.; Hanel, H. Restricted development of the parasitic mite Varroa jacobsoni Oud. in the Cape honeybee Apis mellifera capensis Esch. Z. Angew. Entomol. 1984, 97, 91–95. [Google Scholar] [CrossRef]
- Büchler, R.; Drescher, W. Variance and heritability of the capped developmental stage in european Apis mellifera L. and its correlation with decreased Varroa jacobsoni Oud. infestation. J. Apic. Res. 1990, 29, 172–176. [Google Scholar] [CrossRef]
- Le Conte, Y.; Bruchou, C.; Benhamouda, K.; Gauthier, C.; Cornuet, J.M. Heritability of the queen brood post-capping stage duration in Apis mellifera mellifera L. Apidologie 1994, 25, 513–519. [Google Scholar] [CrossRef] [Green Version]
- Schousboe, C. Undersogelse af forseglingsperiodens længde hos honningbiyngel (Apis mellifera L.) med henblik på forbedring af resistens mod Varroa-miden (Varroa jacobsoni Oud.). Tidsskr. Planteavl. 1986, 90, 293–299. [Google Scholar]
- Moritz, R.F.A.; Jordan, M. Selection of resistance against Varroa jacobsoni across Caste and Sex in the Honeybee (Apis mellifera L, Hymenoptera, Apidae). Exp. Appl. Acarol. 1992, 16, 345–353. [Google Scholar] [CrossRef]
- Siuda, M.; Wilde, J. The parental effect on the progeny brood post-capping stage duration. Pszczenicze Zesz. Nauk. 1996, 40, 7–14. [Google Scholar]
- Wilde, J.; Koeniger, N. Selektion auf Verkürzung der Zellverdecklungsdauer (ZVD) der Arbeiterinnenbrut von Apis mellifera carnica. Ann. Univ. Mariae Curie Sklodowska Lub. Pol. 1992, 47, 133–136. [Google Scholar]
- Siuda, M.; Wilde, J. Varroa resistance of honey bees with a shorter postcapping stage duration. Pszcelnicze Zesyty Nauk. 1998, 42, 75–76. [Google Scholar]
- Siuda, M.; Wilde, J.; Koeniger, N. Further research on honey bee breeding with short post-capping periods. Pszcelnicze Zesyty Nauk. 1996, 40, 135–143. [Google Scholar]
- Lusby, D.A. Small cell size foundation for mite control. Am. Bee J. 1996, 136, 468–470. [Google Scholar]
- Lusby, D.A. More on small cell foundation for mite control. Am. Bee J. 1997, 137, 411–412. [Google Scholar]
- Saucy, F. On the natural cell size of European honey bees: A "fatal error" or distortion of historical data? J. Apic. Res. 2014, 53, 327–336. [Google Scholar] [CrossRef] [Green Version]
- Ball, B.V.; Allen, M.F. The prevalence of pathogens in honey bee (Apis mellifera) colonies infested with the parasite mite Varroa jacobsoni. Ann. Appl. Biol. 1988, 113, 237–244. [Google Scholar] [CrossRef]
- Bailey, L.; Ball, B.V. Honey Bee Pathology; Academic Press: Cambridge, MA, USA, 1991; 193p, ISBN 978-148328-8093. [Google Scholar]
- Grozinger, C.M.; Flenniken, M.L. Bee Viruses: Ecology, Pathogenicity, and Impacts. In Annual Review of Entomology; Douglas, A.E., Ed.; Annual Reviews: Palo Alto, CA, USA, 2019; Volume 64, pp. 205–226. [Google Scholar]
- Martin, S.J.; Highfield, A.C.; Brettell, L.; Villalobos, E.M.; Budge, G.E.; Powell, M.; Nikaido, S.; Schroeder, D.C. Global Honey Bee Viral Landscape Altered by a Parasitic Mite. Science 2012, 336, 1304–1306. [Google Scholar] [CrossRef] [PubMed]
- Mordecai, G.J.; Wilfert, L.; Martin, S.J.; Jones, I.M.; Schroeder, D.C. Diversity in a honey bee pathogen: First report of a third master variant of the Deformed Wing Virus quasispecies. ISME J. 2016, 10, 1264–1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gisder, S.; Mockel, N.; Eisenhardt, D.; Genersch, E. In vivo evolution of viral virulence: Switching of deformed wing virus between hosts results in virulence changes and sequence shifts. Environ. Microbiol. 2018, 20, 4612–4628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalmon, A.; Desbiez, C.; Coulon, M.; Thomasson, M.; Le Conte, Y.; Alaux, C.; Vallon, J.; Moury, B. Evidence for positive selection and recombination hotspots in Deformed wing virus (DWV). Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Mondet, F.; de Miranda, J.R.; Kretzschmar, A.; Le Conte, Y.; Mercer, A.R. On the Front Line: Quantitative Virus Dynamics in Honeybee (Apis mellifera L.) Colonies along a New Expansion Front of the Parasite Varroa destructor. PLoS Pathog. 2014, 10. [Google Scholar] [CrossRef] [Green Version]
- Mordecai, G.J.; Brettell, L.E.; Martin, S.J.; Dixon, D.; Jones, I.M.; Schroeder, D.C. Superinfection exclusion and the long-term survival of honey bees in Varroa-infested colonies. ISME J. 2016, 10, 1182–1191. [Google Scholar] [CrossRef]
- Locke, B.; Forsgren, E.; de Miranda, J.R. Increased Tolerance and Resistance to Virus Infections: A Possible Factor in the Survival of Varroa destructor-Resistant Honey Bees (Apis mellifera). PLoS ONE 2014, 9, e99998. [Google Scholar] [CrossRef] [Green Version]
- Brettell, L.E.; Mordecai, G.J.; Schroeder, D.C.; Jones, I.M.; da Silva, J.R.; Vicente-Rubiano, M.; Martin, S.J. A Comparison of Deformed Wing Virus in Deformed and Asymptomatic Honey Bees. Insects 2017, 8, 28. [Google Scholar] [CrossRef] [Green Version]
- Lattorff, H.M.G.; Buchholz, J.; Fries, I.; Moritz, R.F.A. A selective sweep in a Varroa destructor resistant honeybee (Apis mellifera) population. Infect. Genet. Evol. 2015, 31, 169–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behrens, D.; Huang, Q.; Gessner, C.; Rosenkranz, P.; Frey, E.; Locke, B.; Moritz, R.F.A.; Kraus, F.B. Three QTL in the honey bee Apis mellifera L. suppress reproduction of the parasitic mite Varroa destructor. Ecol. Evol. 2011, 1, 8. [Google Scholar] [CrossRef]
- Conlon, B.H.; Frey, E.; Rosenkranz, P.; Locke, B.; Moritz, R.F.A.; Routtu, J. The role of epistatic interactions underpinning resistance to parasitic Varroa mites in haploid honey bee (Apis mellifera) drones. J. Evol. Biol. 2018, 31, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Broeckx, B.J.G.; De Smet, L.; Blacquiere, T.; Maebe, K.; Khalenkow, M.; Van Poucke, M.; Dahle, B.; Neumann, P.; Nguyen, K.B.; Smagghe, G.; et al. Honey bee predisposition of resistance to ubiquitous mite infestations. Sci. Rep. 2019, 9, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.C.; Du, Z.P.G.; Bernstein, R.; Meyer, M.; Hoppe, A.; Schilling, E.; Ableitner, M.; Juling, K.; Dick, R.; Strauss, A.S.; et al. Tool for genomic selection and breeding to evolutionary adaptation: Development of a 100K single nucleotide polymorphism array for the honey bee. Ecol. Evol. 2020, 10, 6246–6256. [Google Scholar] [CrossRef] [PubMed]
- Meixner, M.D.; Büchler, R.; Costa, C.; Francis, R.M.; Hatjina, F.; Kryger, P.; Uzunov, A.; Carreck, N.L. Honey bee genotypes and the environment. J. Apic. Res. 2014, 53, 183–187. [Google Scholar] [CrossRef] [Green Version]
- Büchler, R.; Costa, C.; Hatjina, F.; Andonov, S.; Meixner, M.D.; Le Conte, Y.; Uzunov, A.; Berg, S.; Bienkowska, M.; Bouga, M.; et al. The influence of genetic origin and its interaction with environmental effects on the survival of Apis mellifera L. colonies in Europe. J. Apic. Res. 2014, 53, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Francis, R.M.; Amiri, E.; Meixner, M.D.; Kryger, P.; Gajda, A.; Andonov, S.; Uzunov, A.; Topolska, G.; Charistos, L.; Costa, C.; et al. Effect of genotype and environment on parasite and pathogen levels in one apiary—A case study. J. Apic. Res. 2014, 53, 230–232. [Google Scholar] [CrossRef]
- Meixner, M.D.; Francis, R.M.; Gajda, A.; Kryger, P.; Andonov, S.; Uzunov, A.; Topolska, G.; Costa, C.; Amiri, E.; Berg, S.; et al. Occurrence of parasites and pathogens in honey bee colonies used in a European genotype-environment interactions experiment. J. Apic. Res. 2014, 53, 215–229. [Google Scholar] [CrossRef]
- Guichard, M.; Neuditschko, M.; Fried, P.; Soland, G.; Dainat, B. A future resistance breeding strategy against Varroa destructor in a small population of the dark honey bee. J. Apic. Res. 2019, 58, 814–823. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
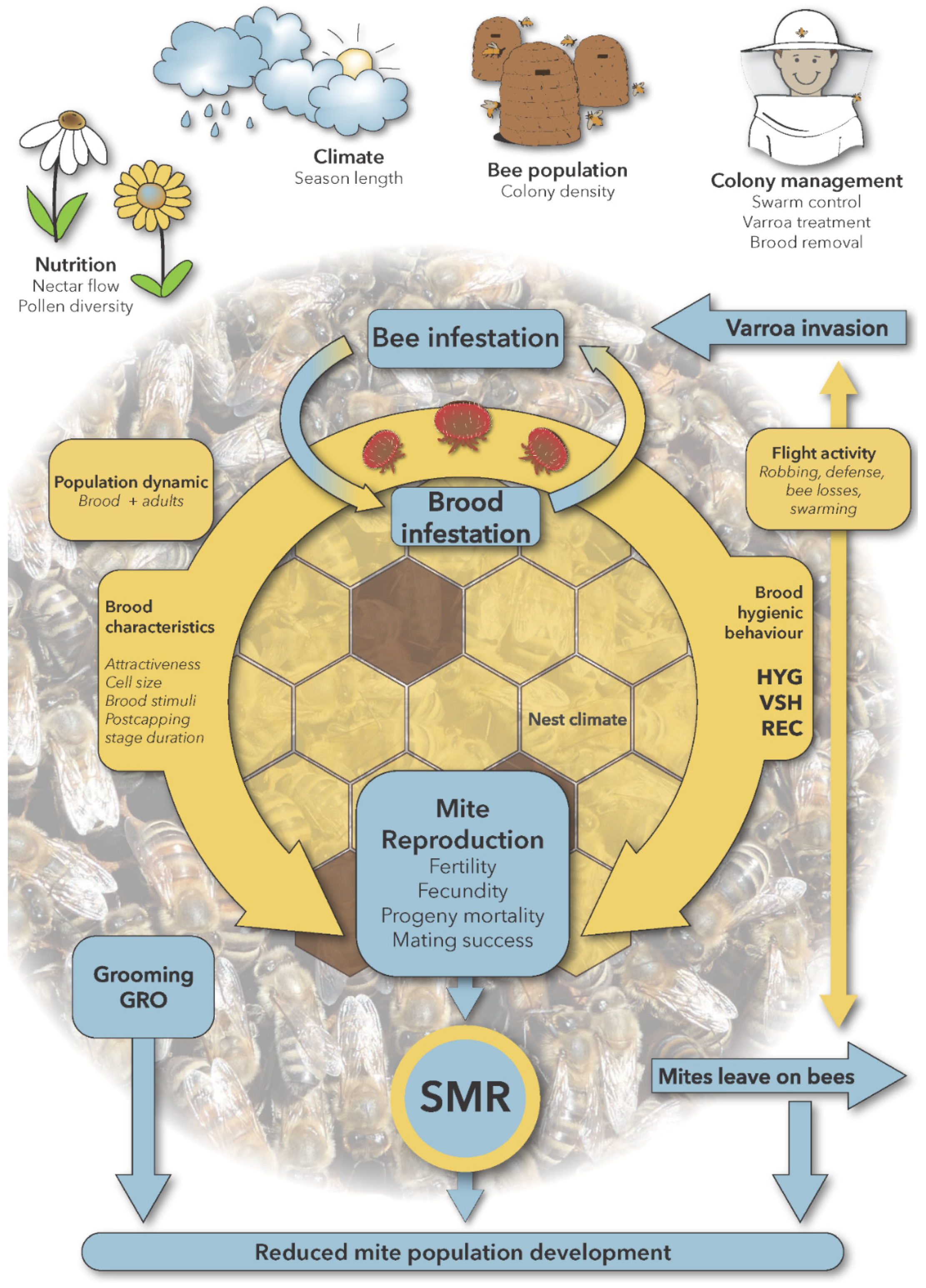
| Character/Parameter | Abbr. | Measures | Description and Remarks | References |
|---|---|---|---|---|
| Bee infestation | Mites/g of bees | Proportion of phoretic mites on adult bees | Rosenkranz et al. 2010 [22] | |
| Mites leave on bees | Bees per day | Loss of bees inside and outside the hive due to mortality and drifting, attached mites are lost | Büchler et al. 2010 [2] | |
| Brood attractiveness | Relative brood infestation of different brood samples under uniform infestation pressure, expressed also as the ratio of mites on bees to mites in brood cells | Rosenkranz et al. 2010 [22] | ||
| Brood dynamic | Change over time in the number of worker and drone brood cells of a colony | Rosenkranz et al. 2010 [22] | ||
| Brood infestation | (adult) mites per brood cell | Proportion of mites in brood cells | Rosenkranz et al. 2010 [22] | |
| Brood stimuli | Response causing agents, which are produced by the brood | Mondet et al., 2016 [88] | ||
| Cell size | Diameter of cells [mm] | smaller cells may affect mite infestation and reproduction | Winston, 1987 [89] | |
| Defense | Scores | Aggressive behavior of worker bees in order to protect the colony | Winston, 1987 [89] | |
| Fecundity | Number of offspring per individual | Potential for reproduction, e.g. max. number of offspring per reproductive cycle | Rosenkranz et al. 2010 [22] | |
| Fertility | Presence of offspring | Capability to produce offspring | Rosenkranz et al. 2010 [22] | |
| Flight activity | Returning bees/min | Flying of worker bees, infested bees or attached mites can be lost in the field | Rosenkranz et al. 2010 [22] | |
| Grooming behavior | GRO | Mite injuries and mite removal | Worker bees detect, remove, damage or destroy the mite from themselves or other workers | Morfin et al., 2020 [90] Guzman-Novoa et al., 2012 [34] |
| Hygienic behavior | HYG | Freeze-killed or pinkilled brood | General hygiene behavior towards diseased/infested brood cells | Dietemann et al., 2013 [7] |
| Varroa invasion | Mites/interval | Mites enter a colony, transported by worker bees | Rosenkranz et al. 2010 [22] | |
| Mite mating success | Rate of successfully impregnated female mites | Rosenkranz et al. 2010 [22] | ||
| Mite mortality | Dead mites on bottom board | Death rate of mites | Dietemann et al., 2013 [7] | |
| Mite reproduction | MR | see with SMR Rate of reproductive mites from single infested worker brood cells | Production of varroa offspring | Mondet et al., 2020 [5] |
| Nest climate | Temperature and humidity level and variation in the brood nest, affects postcapping stage duration and mite reproduction | Kraus et al., 1997 [91] | ||
| Population dynamic | Change over time in the number and ratio of worker and drone brood and bees in a colony, influenced by season, swarming, etc. | Büchler et al. 2010 [2] | ||
| Postcapping stage duration | PCD | Hours between cell capping and emergence | Time span between closing of brood cell and emergence of adult bee. A prolonged postcapping period of the brood cell should reduce the number of mature adult daughter mites. However, a shorter developmental time of the brood probably permits one brood cycle more per season. | Rosenkranz et al. 2010 [22] |
| Progeny mortality | Dead offspring in cells | Death rate of mite offspring | Villa et al., 2009 [92] | |
| Recapping | REC | Inner side of cell cap without cocoon | Opening and closing (recapping) of sealed brood cells, may be non-infested or infested | Oddie et al. 2018 [63] |
| Reduced mite population development | MPD | Comparison of infestation levels during time | Attenuated increase in the number of mites per colony; is affected by mite reproduction, population dynamics, grooming and flight activity (varroa invasion, mites that leave on bees) | Büchler et al., 2010 [2] |
| Robbing | Worker bees steal honey from foreign colonies, phoretic mites may leave or enter a colony | Winston, 1987 [89] | ||
| Suppressed mite reproduction | SMR | Rate of non-reproductive mites from single infested worker brood cells | Heritable trait of the honey bee that negatively influences varroa reproduction | Harbo and Harris, 2005 [93] |
| Swarming | Scores | Reproduction of the colony: the old queen leaves with about half of the worker bees, brood interruption and split of mite population reduces infestation level | Fries et al., 2003 [77], Büchler et al., 2013 [94] | |
| Varroa sensitive hygiene | VSH | Rate of removal from all infested worker brood cells | Uncapping and removal of Varroa infested brood cells | Villa et al., 2009 [92] |
| Country | Naturally Selected Resistant Populations Present | If Yes, How Many | Proven Examples of Survivor-Stock | If Yes, How Many | Are There Selection Programs for Varroa Resistance | If Yes, are Queens Commercially Available? | If Yes, at What Price |
|---|---|---|---|---|---|---|---|
| Austria | none known | No | yes, two | Yes, but not marketed as ‘resistant’ | no data | ||
| Belgium | none known | no | yes, starting: three | no | |||
| Bulgaria | anecdotal reports | no data available. | no; but selection on hygienic behavior carried out | ||||
| Croatia | anecdotal reports | no | yes | no | |||
| Cyprus | none known | no | no | ||||
| Czech Republic | none known | no | no, but selection on hygienic behavior | ||||
| Denmark | none known | no | Just initiated. No sale, no price. | ||||
| Estonia | none known | no | no | ||||
| Finland | none known | no | starting | ||||
| France | yes | 3: Avignon, Sarthe, Tarn | yes, several initiatives | no | |||
| Germany | none known | anecdotal reports | yes, several initiatives, at least three | Yes, but not marketed as ‘resistant’ | EUR 50–EUR 75 | ||
| Greece | anecdotal reports | anecdotal reports | few-only individual initiatives | no | |||
| Hungary | no | no | yes, based on VSH | no | |||
| Ireland | anecdotal reports | no | yes | no | |||
| Italy | yes | 4: Liguria (A.m.m.), Eolie (A.m.s.), Vicenza | Gorgona (A.m.l.), | yes (one public and two private breeders (Ligustica, and Carnica-mix). | no | ||
| Latvia | none known | no | no | ||||
| Lithuania | yes | local hybridized bees of A.m mellifera near Belarus border | anecdotal reports | no | |||
| Luxembourg | |||||||
| Malta | none known | no | starting (Smartbees project) | no | |||
| Netherlands | yes | 3: Lelystad, Tiengemeten and at Laren | anecdotal reports | yes, 4 in total | no | ||
| Poland | none known | no | yes | no | |||
| Portugal | none known | no | no | ||||
| Romania | none known | no | starting (Smartbees project) | no | |||
| Slovakia | none known | no | yes, to some extent | yes | EUR 20 | ||
| Slovenia | none known | no | yes | no | |||
| Spain | none known | no | starting (Smartbees project) | no | |||
| Sweden | yes | southern Gotland (the Bond bees) | anecdotal reports. | yes, one based on VSH, and some other small projects | no | ||
| United Kingdom | anecdotal reports | Lleyn peninsula | possibly… | yes, one University program (hygienic) | sometimes, small scale | no data | |
| Israel | none known | no | no | ||||
| Macedonia | none known | no | no | ||||
| Norway | yes | one in southeast Norway, untreated since 1997 | no | few initiatives (Smartbees) | very limited amounts | EUR 75 | |
| Switzerland | none known | anecdotal reports | no | ||||
| Turkey | yes | Marmara Island | no | yes, based on Marmara Island stock | yes | EUR 15–EUR 20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Le Conte, Y.; Meixner, M.D.; Brandt, A.; Carreck, N.L.; Costa, C.; Mondet, F.; Büchler, R. Geographical Distribution and Selection of European Honey Bees Resistant to Varroa destructor. Insects 2020, 11, 873. https://doi.org/10.3390/insects11120873
Le Conte Y, Meixner MD, Brandt A, Carreck NL, Costa C, Mondet F, Büchler R. Geographical Distribution and Selection of European Honey Bees Resistant to Varroa destructor. Insects. 2020; 11(12):873. https://doi.org/10.3390/insects11120873
Chicago/Turabian StyleLe Conte, Yves, Marina D. Meixner, Annely Brandt, Norman L. Carreck, Cecilia Costa, Fanny Mondet, and Ralph Büchler. 2020. "Geographical Distribution and Selection of European Honey Bees Resistant to Varroa destructor" Insects 11, no. 12: 873. https://doi.org/10.3390/insects11120873
APA StyleLe Conte, Y., Meixner, M. D., Brandt, A., Carreck, N. L., Costa, C., Mondet, F., & Büchler, R. (2020). Geographical Distribution and Selection of European Honey Bees Resistant to Varroa destructor. Insects, 11(12), 873. https://doi.org/10.3390/insects11120873