Using RNA Interference to Reveal the Function of Chromatin Remodeling Factor ISWI in Temperature Tolerance in Bemisia tabaci Middle East–Asia Minor 1 Cryptic Species


Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Total RNA Isolation and Reverse Transcription
2.3. Cloning and Sequencing of BtISWI
2.4. Bioinformatics Analysis
2.5. Real-Time Quantitative PCR
2.6. Temperature Stress Treatment
2.7. RNA Interference
2.8. Thermal Tolerance Test after dsRNA Feeding
2.9. Temperature Preference Behavioral Assay
2.10. Statistical Analysis
3. Results
3.1. Characterization of BtISWI
3.2. Expression Profiles of BtISWI in Response to Thermal Stress
3.3. BtISWI mRNA Expression Level after Feeding dsRNA
3.4. The Role of BtISWI in Temperature Stress
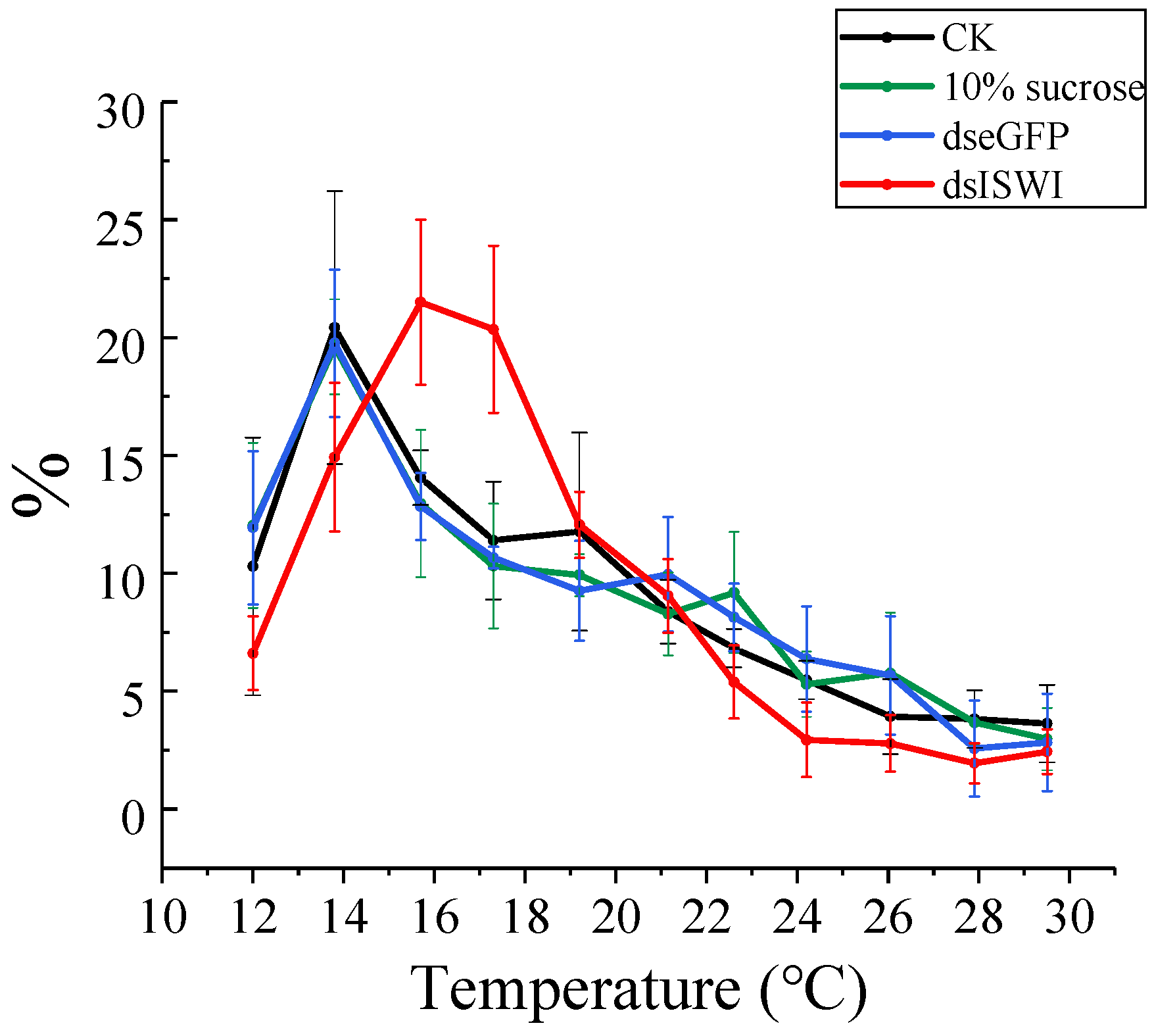
3.5. The Choice of Preferred Temperature
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- An, P.N.T.; Shimaji, K.; Tanaka, R.; Yoshida, H.; Kimura, H.; Fukusaki, E.; Yamaguchi, M. Epigenetic regulation of starvation-induced autophagy in Drosophila by histone methyltransferase G9a. Sci. Rep. 2017, 7, 7343. [Google Scholar] [CrossRef] [Green Version]
- Stankiewicz, A.M.; Swiergiel, A.H.; Lisowski, P. Epigenetics of stress adaptations in the brain. Brain Res. Bull. 2013, 98, 76–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skinner, M.K.; Manikkam, M.; Guerrero-Bosagna, C. Epigenetic transgenerational actions of environmental factors in disease etiology. Trends Endocrinol. Metab. 2010, 21, 214–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dukes, J.S. Responses of invasive species to a changing climate and atmosphere. In Fifty Years of Invasion Ecology: The Legacy of Charles Elton; Richardson, D.M., Ed.; Wiley-Blackwell: Oxford, UK, 2011; pp. 345–357. [Google Scholar]
- Richardson, D.M.; Pyšek, P. Fifty years of invasion ecology-the legacy of Charles Elton. Divers. Distrib. 2008, 14, 161–168. [Google Scholar] [CrossRef]
- Ward, N.L.; Masters, G.J. Linking climate change and species invasion: An illustration using insect herbivores. Glob. Chang. Biol. 2007, 13, 1605–1615. [Google Scholar] [CrossRef]
- Zhan, A.B.; Briski, E.; Bock, D.G.; Ghabooli, S.; MacIsaac, H.J. Ascidians as models for studying invasion success. Mar. Biol. 2015, 162, 2449–2470. [Google Scholar] [CrossRef]
- Estoup, A.; Ravigné, V.; Hufbauer, R.; Vitalis, R.; Gautier, M.; Facon, B. Is there a genetic paradox of biological invasion? Annu. Rev. Ecol. Evol. Syst. 2016, 47, 51–72. [Google Scholar] [CrossRef]
- Hawes, N.A.; Tremblay, L.A.; Pochon, X.; Dunphy, B.; Fidler, A.E.; Smith, K.F. Effects of temperature and salinity stress on DNA methylation in a highly invasive marine invertebrate, the colonial ascidian Didemnum vexillum. PeerJ 2018, 6, e5003. [Google Scholar] [CrossRef] [Green Version]
- Pérez, J.E.; Nirchio, M.; Alfonsi, C.; Muñoz, C. The biology of invasions: The genetic adaptation paradox. Biol. Invasions 2006, 8, 1115–1121. [Google Scholar] [CrossRef]
- Prentis, P.J.; Wilson, J.R.; Dormontt, E.E.; Richardson, D.M.; Lowe, A.J. Adaptive evolution in invasive species. Trends Plant Sci. 2008, 13, 288–294. [Google Scholar] [CrossRef]
- Corona, D.F.; Tamkun, J.W. Multiple roles for ISWI in transcription, chromosome organization and DNA replication. BBA-Gene Struct. Expr. 2004, 1677, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Hopfner, K.-P.; Gerhold, C.-B.; Lakomek, K.; Wollmann, P. Swi2/Snf2 remodelers: Hybrid views on hybrid molecular machines. Curr. Opin. Struct. Biol. 2012, 22, 225–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cairns, B.R.; Kim, Y.-J.; Sayre, M.H.; Laurent, B.C.; Kornberg, R.D. A multisubunit complex containing the SWI1/ADR6, SWI2/SNF2, SWI3, SNF5, and SNF6 gene products isolated from yeast. Proc. Natl. Acad. Sci. USA 1994, 91, 1950–1954. [Google Scholar] [CrossRef] [Green Version]
- Ito, T.; Bulger, M.; Pazin, M.J.; Kobayashi, R.; Kadonaga, J.T. ACF, an ISWI-containing and ATP-utilizing chromatin assembly and remodeling factor. Cell 1997, 90, 145–155. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Mizuguchi, G.; Hamiche, A.; Wu, C. A chromatin remodelling complex involved in transcription and DNA processing. Nature 2000, 406, 541. [Google Scholar] [CrossRef]
- Erdel, F.; Rippe, K. Binding kinetics of human ISWI chromatin-remodelers to DNA repair sites elucidate their target location mechanism. Nucleus 2011, 2, 19873–19878. [Google Scholar] [CrossRef] [Green Version]
- Bouazoune, K.; Brehm, A. ATP-dependent chromatin remodeling complexes in Drosophila. Chromosome Res. 2006, 14, 433–449. [Google Scholar] [CrossRef]
- Toto, M.; D’Angelo, G.; Corona, D.F. Regulation of ISWI chromatin remodelling activity. Chromosoma 2014, 123, 91–102. [Google Scholar] [CrossRef] [Green Version]
- Aydin, Ö.Z.; Marteijn, J.A.; Ribeiro-Silva, C.; Rodríguez López, A.; Wijgers, N.; Smeenk, G.; van Attikum, H.; Poot, R.A.; Vermeulen, W.; Lans, H. Human ISWI complexes are targeted by SMARCA5 ATPase and SLIDE domains to help resolve lesion-stalled transcription. Nucleic Acids Res. 2014, 42, 8473–8485. [Google Scholar] [CrossRef]
- Badenhorst, P.; Voas, M.; Rebay, I.; Wu, C. Biological functions of the ISWI chromatin remodeling complex NURF. Genes Dev. 2002, 16, 3186–3198. [Google Scholar] [CrossRef] [Green Version]
- Tsukiyama, T.; Becker, P.B.; Wu, C. ATP-dependent nucleosome disruption at a heat-shock promoter mediated by binding of GAGA transcription factor. Nature 1994, 367, 525. [Google Scholar] [CrossRef] [Green Version]
- Erkina, T.Y.; Zou, Y.; Freeling, S.; Vorobyev, V.; Erkine, A.M. Functional interplay between chromatin remodeling complexes RSC, SWI/SNF and ISWI in regulation of yeast heat shock genes. Nucleic Acids Res. 2009, 38, 1441–1449. [Google Scholar] [CrossRef] [Green Version]
- Buszewicz, D.; Archacki, R.; Palusiński, A.; Kotliński, M.; Fogtman, A.; Iwanicka-Nowicka, R.; Sosnowska, K.; Kuciński, J.; Pupel, P.; Olędzki, J.; et al. HD2C histone deacetylase and a SWI/SNF chromatin remodelling complex interact and both are involved in mediating the heat stress response in Arabidopsis. Plant Cell Environ. 2016, 39, 2108–2122. [Google Scholar] [CrossRef]
- Goldmark, J.P.; Fazzio, T.G.; Estep, P.W.; Church, G.M.; Tsukiyama, T. The Isw2 chromatin remodeling complex represses early meiotic genes upon recruitment by Ume6p. Cell 2000, 103, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Wagner, D. Chromatin regulation of plant development. Curr. Opin. Plant Biol. 2003, 6, 20–28. [Google Scholar] [CrossRef]
- Collins, N.; Poot, R.A.; Kukimoto, I.; García-Jiménez, C.; Dellaire, G.; Varga-Weisz, P.D. An ACF1-ISWI chromatin-remodeling complex is required for DNA replication through heterochromatin. Nat. Genet. 2002, 32, 627. [Google Scholar] [CrossRef]
- Brown, J.K.; Frohlich, D.; Rosell, R.C. The sweetpotato or silverleaf whiteflies: Biotypes of Bemisia tabaci or a species complex? Annu. Rev. Entomol. 1995, 40, 511–534. [Google Scholar] [CrossRef]
- De Barro, P.J.; Liu, S.S.; Boykin, L.M.; Dinsdale, A.B. Bemisia tabaci: A statement of species status. Annu. Rev. Entomol. 2011, 56, 1–19. [Google Scholar] [CrossRef]
- Liu, S.S.; Colvin, J.; De Barro, P.J. Species concepts as applied to the whitefly Bemisia tabaci systematics: How many species are there? J. Integr. Agric. 2012, 11, 176–186. [Google Scholar] [CrossRef]
- Wang, X.W.; Yang, N.W. The Whitefly Bemisia tabaci (Gennadius). In Biological Invasions and Its Management in China; Wan, F.H., Jiang, M.X., Zhan, A.B., Eds.; Springer: Dordrecht, The Netherlands, 2017; pp. 159–182. [Google Scholar]
- Broadbent, A.B.; Foottit, R.G.; Murphy, G.D. Sweetpotato whitefly Bemisia tabaci (Gennadius) (Homoptera: Aleyrodidae), a potential insect pest in Canada. Can. Entomol. 1989, 121, 1027–1028. [Google Scholar] [CrossRef]
- Lee, C.E. Evolutionary genetics of invasive species. Trends Ecol. Evol. 2002, 17, 386–391. [Google Scholar] [CrossRef]
- Wan, F.H.; Zhang, G.F.; Liu, S.S.; Luo, C.; Chu, D.; Zhang, Y.J.; Zang, L.S.; Jiu, M.; Lü, Z.C.; Cui, X.H. Invasive mechanism and management strategy of Bemisia tabaci (Gennadius) biotype B: Progress report of 973 Program on invasive alien species in China. Sci. China Ser. C Life Sci. 2009, 52, 88–95. [Google Scholar] [CrossRef]
- Yu, H.; Wan, F.H.; Guo, J.Y. Different thermal tolerance and hsp gene expression in invasive and indigenous sibling species of Bemisia tabaci. Biol. Invasions 2012, 14, 1587–1595. [Google Scholar] [CrossRef]
- Lü, Z.C.; Wan, F.H. Differential gene expression in whitefly (Bemisia tabaci) B-biotype females and males under heat-shock condition. Comp. Biochem. Physiol. Part D Genom. Proteom. 2008, 3, 257–262. [Google Scholar] [CrossRef]
- Mahadav, A.; Kontsedalov, S.; Czosnek, H.; Ghanim, M. Thermotolerance and gene expression following heat stress in the whitefly Bemisia tabaci B and Q biotypes. Insect Biochem. Mol. Biol. 2009, 39, 668–676. [Google Scholar] [CrossRef]
- Lü, Z.C.; Li, Q.; Liu, W.X.; Wan, F.H. Transient receptor potential is essential for high temperature tolerance in invasive Bemisia tabaci Middle East Asia minor 1 cryptic species. PLoS ONE 2014, 9, e108428. [Google Scholar] [CrossRef]
- Dai, T.M.; Lü, Z.C.; Liu, W.X.; Wan, F.H.; Hong, X.Y. The homology gene BtDnmt1 is Essential for Temperature Tolerance in Invasive Bemisia tabaci Mediterranean Cryptic Species. Sci. Rep. 2017, 7, 3040. [Google Scholar] [CrossRef] [Green Version]
- Dai, T.M.; Wang, Y.S.; Liu, W.X.; Lü, Z.C.; Wan, F.H. Thermal Discrimination and Transgenerational Temperature Response in Bemisia tabaci Mediterranean (Hemiptera: Aleyrodidae): Putative Involvement of the Thermo-Sensitive Receptor BtTRPA. Environ. Entomol. 2018, 47, 204–209. [Google Scholar] [CrossRef]
- Lü, Z.C.; Wang, Y.M.; Zhu, S.G.; Yu, H.; Guo, J.Y.; Wan, F.H. Trade-offs between survival, longevity, and reproduction, and variation of survival tolerance in Mediterranean Bemisia tabaci after temperature stress. J. Insect Sci. (Oxford UK) 2014, 14, 124. [Google Scholar] [CrossRef]
- Dai, T.M.; Lü, Z.C.; Wang, Y.S.; Liu, W.X.; Hong, X.Y.; Wan, F.H. Molecular characterizations of DNA methyltransferase 3 and its roles in temperature tolerance in the whitefly, Bemisia tabaci Mediterranean. Insect Mol. Biol. 2018, 27, 123–132. [Google Scholar] [CrossRef]
- Chen, W.; Hasegawa, D.K.; Kaur, N.; Kliot, A.; Pinheiro, P.V.; Luan, J.; Stensmyr, M.C.; Zheng, Y.; Liu, W.; Sun, H. The draft genome of whitefly Bemisia tabaci MEAM1, a global crop pest, provides novel insights into virus transmission, host adaptation, and insecticide resistance. BMC Biol. 2016, 14, 110. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Whelan, S.; Goldman, N. A general empirical model of protein evolution derived from multiple protein families using a maximum-likelihood approach. Mol. Biol. Evol. 2001, 18, 691–699. [Google Scholar] [CrossRef] [Green Version]
- Dai, T.M.; Lü, Z.C.; Liu, W.X.; Wan, F.H. Selection and validation of reference genes for qRT-PCR analysis during biological invasions: The thermal adaptability of Bemisia tabaci MED. PLoS ONE 2017, 12, e0173821. [Google Scholar] [CrossRef]
- Bowler, K.; Terblanche, J.S. Insect thermal tolerance: What is the role of ontogeny, ageing and senescence? Biol. Rev. 2008, 83, 339–355. [Google Scholar] [CrossRef]
- Lü, Z.C.; Wan, F.H. Using double-stranded RNA to explore the role of heat shock protein genes in heat tolerance in Bemisia tabaci (Gennadius). J. Exp. Biol. 2011, 214, 764–769. [Google Scholar] [CrossRef] [Green Version]
- Bubli, O.A.; Imasheva, A.G.; Loeschcke, V. Selection for knockdown resistance to heat in Drosophila melanogaster at high and low larval densities. Evolution 1998, 52, 619–625. [Google Scholar] [CrossRef]
- Ma, F.Z.; Lü, Z.C.; Wang, R.; Wan, F.H. Heritability and evolutionary potential in thermal tolerance traits in the invasive Mediterranean cryptic species of Bemisia tabaci (Hemiptera: Aleyrodidae). PLoS ONE 2014, 9, e103279. [Google Scholar] [CrossRef]
- Mitchell, K.A.; Hoffmann, A.A. Thermal ramping rate influences evolutionary potential and species differences for upper thermal limits in Drosophila. Funct. Ecol. 2010, 24, 694–700. [Google Scholar] [CrossRef]
- Gibert, P.; Moreteau, B.; Pétavy, G.; Karan, D.; David, J.R. Chill-coma tolerance, a major climatic adaptation among Drosophila species. Evolution 2001, 55, 1063–1068. [Google Scholar] [CrossRef]
- Head, L.M.; Tang, X.; Hayley, S.E.; Goda, T.; Umezaki, Y.; Chang, E.C.; Leslie, J.R.; Fujiwara, M.; Garrity, P.A.; Hamada, F.N. The influence of light on temperature preference in Drosophila. Curr. Biol. 2015, 25, 1063–1068. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Mathew, R.P.; Mishra, R.K.; Pandey, S.M. ISWI Chromatin Remodeling Complexes: Composition and Regulation perspectives. J. Sci. Res. 2018, 62, 133–145. [Google Scholar]
- Grüne, T.; Brzeski, J.; Eberharter, A.; Clapier, C.R.; Corona, D.F.; Becker, P.B.; Müller, C.W. Crystal structure and functional analysis of a nucleosome recognition module of the remodeling factor ISWI. Mol. Cell 2003, 12, 449–460. [Google Scholar] [CrossRef]
- Mueller-Planitz, F.; Klinker, H.; Ludwigsen, J.; Becker, P.B. The ATPase domain of ISWI is an autonomous nucleosome remodeling machine. Nat. Struct. Mol. Biol. 2013, 20, 82. [Google Scholar] [CrossRef]
- De Barro, P.J.; Driver, F.; Trueman, J.W.; Curran, J. Phylogenetic relationships of world populations of Bemisia tabaci (Gennadius) using ribosomal ITS1. Mol. Phylogenet. Evol. 2000, 16, 29–36. [Google Scholar] [CrossRef]
- Cui, X.H.; Wan, F.H.; Xie, M.; Liu, T. Effects of heat shock on survival and reproduction of two whitefly species, Trialeurodes vaporariorum and Bemisia tabaci biotype B. J. Insect Sci. (Oxford, UK) 2008, 8, 24. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Valencia, V.; González-Obando, R.; Díaz, F. Rapid responses to a strong experimental selection for heat hardening in the invasive whitefly Bemisia tabaci MEAM 1. Entomol. Exp. Appl. 2016, 160, 147–155. [Google Scholar] [CrossRef]
- Yu, H.; Wan, F.H. Cloning and expression of heat shock protein genes in two whitefly species in response to thermal stress. J. Appl. Entomol. 2009, 133, 602–614. [Google Scholar] [CrossRef]
- Tsukiyama, T.; Daniel, C.; Tamkun, J.; Wu, C. ISWI, a member of the SWl2/SNF2 ATPase family, encodes the 140 kDa subunit of the nucleosome remodeling factor. Cell 1995, 83, 1021–1026. [Google Scholar] [CrossRef] [Green Version]
- Shivaswamy, S.; Iyer, V.R. Stress-dependent dynamics of global chromatin remodeling in yeast: Dual role for SWI/SNF in the heat shock stress response. Mol. Cell. Biol. 2008, 28, 2221–2234. [Google Scholar] [CrossRef] [Green Version]
- Huey, R.; Crill, W.; Kingsolver, J.; Weber, K. A method for rapid measurement of heat or cold resistance of small insects. Funct. Ecol. 1992, 6, 489–494. [Google Scholar] [CrossRef] [Green Version]
- Goda, T.; Leslie, J.R.; Hamada, F.N. Design and analysis of temperature preference behavior and its circadian rhythm in Drosophila. JoVE-J. Vis. Exp. 2014, 83, e51097. [Google Scholar] [CrossRef]
- Kaneko, H.; Head, L.M.; Ling, J.; Tang, X.; Liu, Y.; Hardin, P.E.; Emery, P.; Hamada, F.N. Circadian rhythm of temperature preference and its neural control in Drosophila. Curr. Biol. 2012, 22, 1851–1857. [Google Scholar] [CrossRef] [Green Version]
- Fu, T.; Hull, J.; Yang, T.; Wang, G.R. Identification and functional characterization of four transient receptor potential ankyrin 1 variants in Apolygus lucorum (Meyer-Dür). Insect Mol. Biol. 2016, 25, 370–384. [Google Scholar] [CrossRef]
- Lee, Y.; Lee, Y.; Lee, J.; Bang, S.; Hyun, S.; Kang, J.; Hong, S.T.; Bae, E.; Kaang, B.-K.; Kim, J. Pyrexia is a new thermal transient receptor potential channel endowing tolerance to high temperatures in Drosophila melanogaster. Nat. Genet. 2005, 37, 305. [Google Scholar] [CrossRef]
- Kinzner, M.T.; Kinzner, M.-C.; Kaufmann, R.; Hoffmann, A.A.; Arthofer, W.; Schlick-Steiner, B.C.; Steiner, F.M. Is temperature preference in the laboratory ecologically relevant for the field? The case of Drosophila nigrosparsa. Glob. Ecol. Conserv. 2019, 18, e00638. [Google Scholar] [CrossRef]
- Dillon, M.E.; Liu, R.; Wang, G.; Huey, R.B. Disentangling thermal preference and the thermal dependence of movement in ectotherms. J. Therm. Biol. 2012, 37, 631–639. [Google Scholar] [CrossRef]
- Huey, R.B.; Hertz, P.E.; Sinervo, B. Behavioral drive versus behavioral inertia in evolution: A null model approach. Am. Nat. 2003, 161, 357–366. [Google Scholar] [CrossRef] [Green Version]
- MacLean, H.J.; Sørensen, J.G.; Kristensen, T.N.; Loeschcke, V.; Beedholm, K.; Kellermann, V.; Overgaard, J. Evolution and plasticity of thermal performance: An analysis of variation in thermal tolerance and fitness in 22 Drosophila species. Philos. Trans. R. Soc. B 2019, 374, 20180548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stratman, R.; Markow, T. Resistance to thermal stress in desert Drosophila. Funct. Ecol. 1998, 12, 965–970. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Primer Sequence (5′ to 3′) |
|---|---|
| Primers for full-length gene amplification | |
| ISWI-F1 | GCTTCAGCCAATATGGCGACT |
| ISWI-R1 | CGGCAGCAGCTTATCAAGAATG |
| ISWI-F2 | GACCTTTCTTGTTGCGTCGTCTC |
| ISWI-R2 | GGCGCTTGATACCTCTCCATTT |
| ISWI-F3 | ACTATTTCCGCAAGTCAGTGGGC |
| ISWI-R3 | GACGACGCATCTCAAGGCTAAA |
| Primers for real-time quantitative PCR | |
| ISWI-QF | GCAGGTTAGATGGTCAAACTCCCC |
| ISWI-QR | TTTTCCTCAACAGTATTTTCGGTG |
| EF1-α-F | TAGCCTTGTGCCAATTTCCG |
| EF1-α-R | CCTTCAGCATTACCGTCC |
| β-tub-F | TGTCAGGAGTAACGACGTGTTTG |
| β-tub-R | TTCGGGAACGGTAAGTGCTC |
| Primers for dsRNA synthesis | |
| ISWI-DF | TAATACGACTCACTATAGGGCTCCGATTCACCCTCT |
| ISWI-DR | TAATACGACTCACTATAGGGGTCCCAGTCTCCAGGC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, S.-X.; Wang, X.-D.; Shen, X.-N.; Liang, L.; Liu, W.-X.; Wan, F.-H.; Lü, Z.-C. Using RNA Interference to Reveal the Function of Chromatin Remodeling Factor ISWI in Temperature Tolerance in Bemisia tabaci Middle East–Asia Minor 1 Cryptic Species. Insects 2020, 11, 113. https://doi.org/10.3390/insects11020113
Ji S-X, Wang X-D, Shen X-N, Liang L, Liu W-X, Wan F-H, Lü Z-C. Using RNA Interference to Reveal the Function of Chromatin Remodeling Factor ISWI in Temperature Tolerance in Bemisia tabaci Middle East–Asia Minor 1 Cryptic Species. Insects. 2020; 11(2):113. https://doi.org/10.3390/insects11020113
Chicago/Turabian StyleJi, Shun-Xia, Xiao-Di Wang, Xiao-Na Shen, Lin Liang, Wan-Xue Liu, Fang-Hao Wan, and Zhi-Chuang Lü. 2020. "Using RNA Interference to Reveal the Function of Chromatin Remodeling Factor ISWI in Temperature Tolerance in Bemisia tabaci Middle East–Asia Minor 1 Cryptic Species" Insects 11, no. 2: 113. https://doi.org/10.3390/insects11020113
APA StyleJi, S. -X., Wang, X. -D., Shen, X. -N., Liang, L., Liu, W. -X., Wan, F. -H., & Lü, Z. -C. (2020). Using RNA Interference to Reveal the Function of Chromatin Remodeling Factor ISWI in Temperature Tolerance in Bemisia tabaci Middle East–Asia Minor 1 Cryptic Species. Insects, 11(2), 113. https://doi.org/10.3390/insects11020113