Propolis Consumption Reduces Nosema ceranae Infection of European Honey Bees (Apis mellifera)

 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sources of Honey Bees and Propolis
2.2. Propolis Extract Preparation
2.3. Pathogen Preparation
2.4. Survival Bioassay Set-Up
2.5. Food Choice Test
2.6. Scan Bees Sampling and Food Consumption Test
2.7. Quantifying N. ceranae Infection: DNA Extraction and qPCR
2.8. Chemical Analyses of Propolis
2.9. Statistical Methods
3. Results
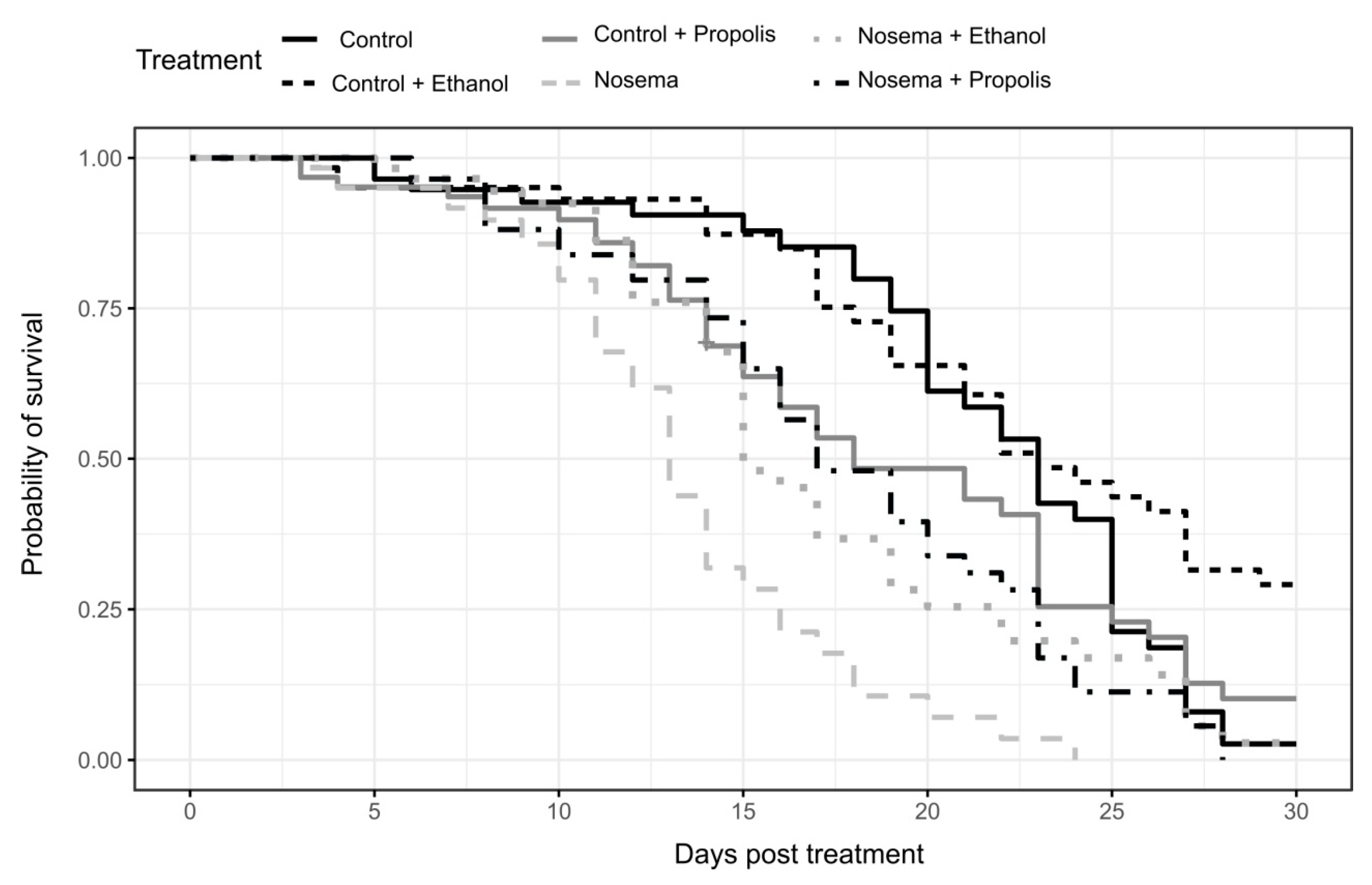
3.1. Survival Bioassay
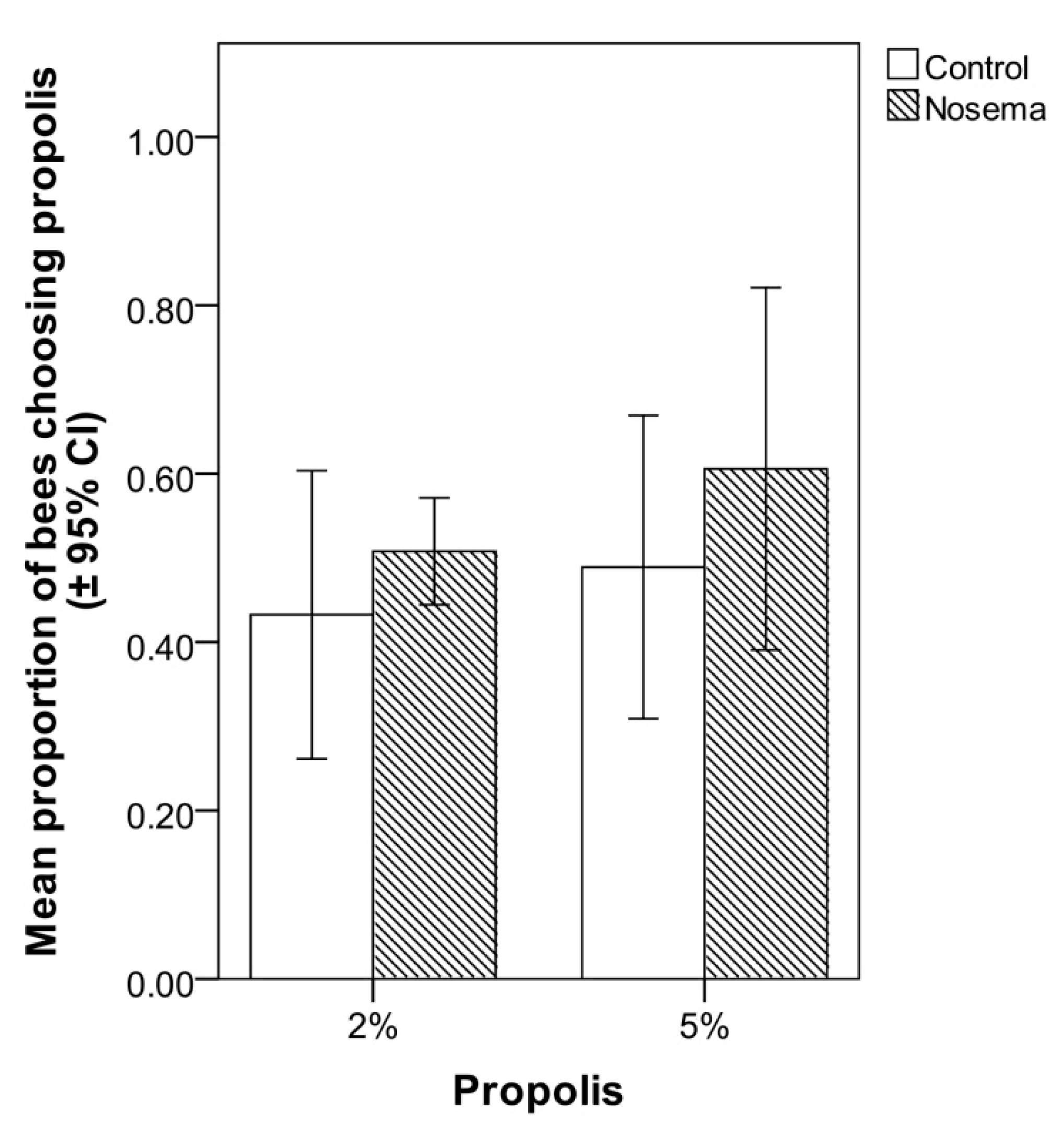
3.2. Food Choice Test
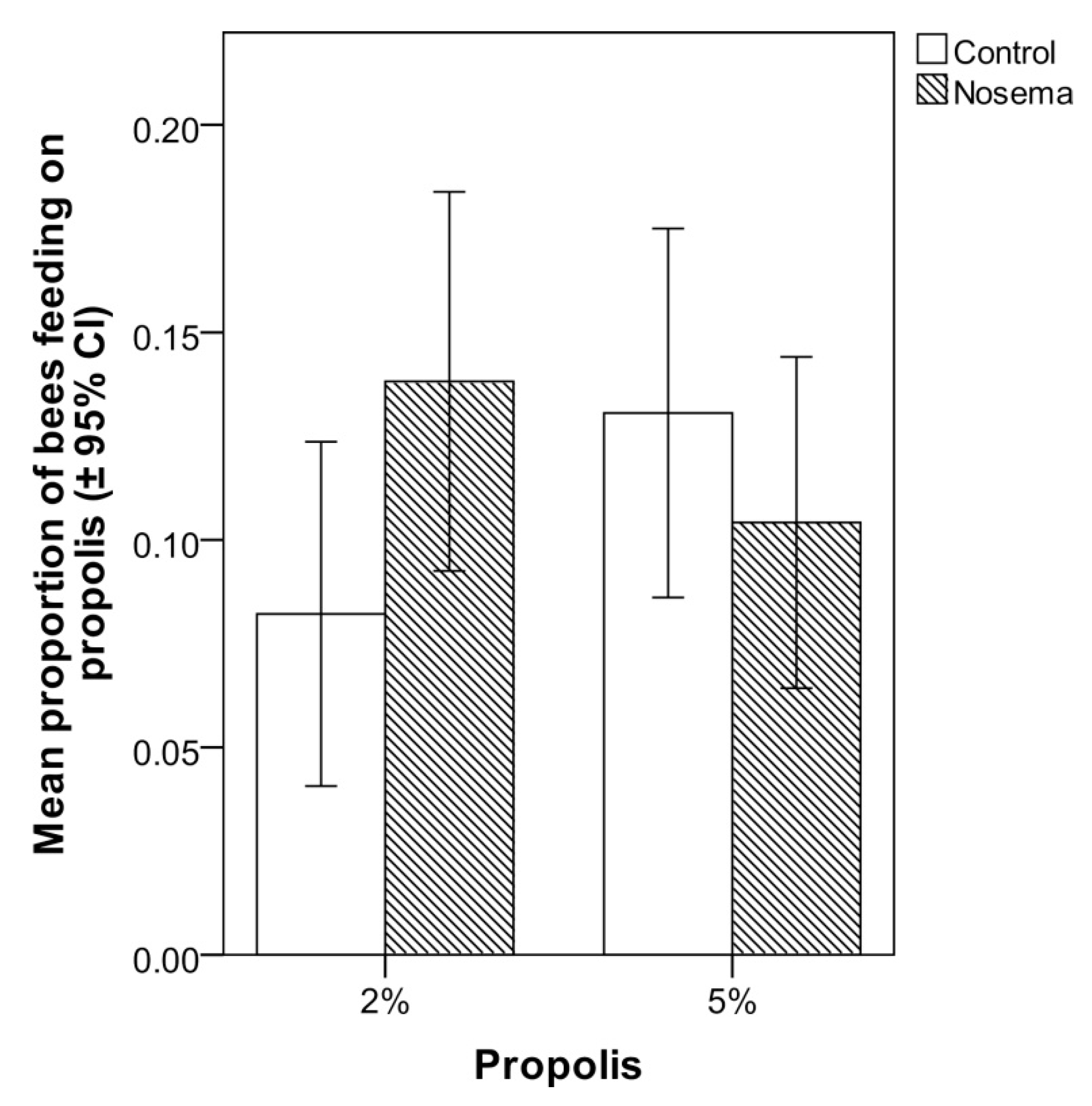
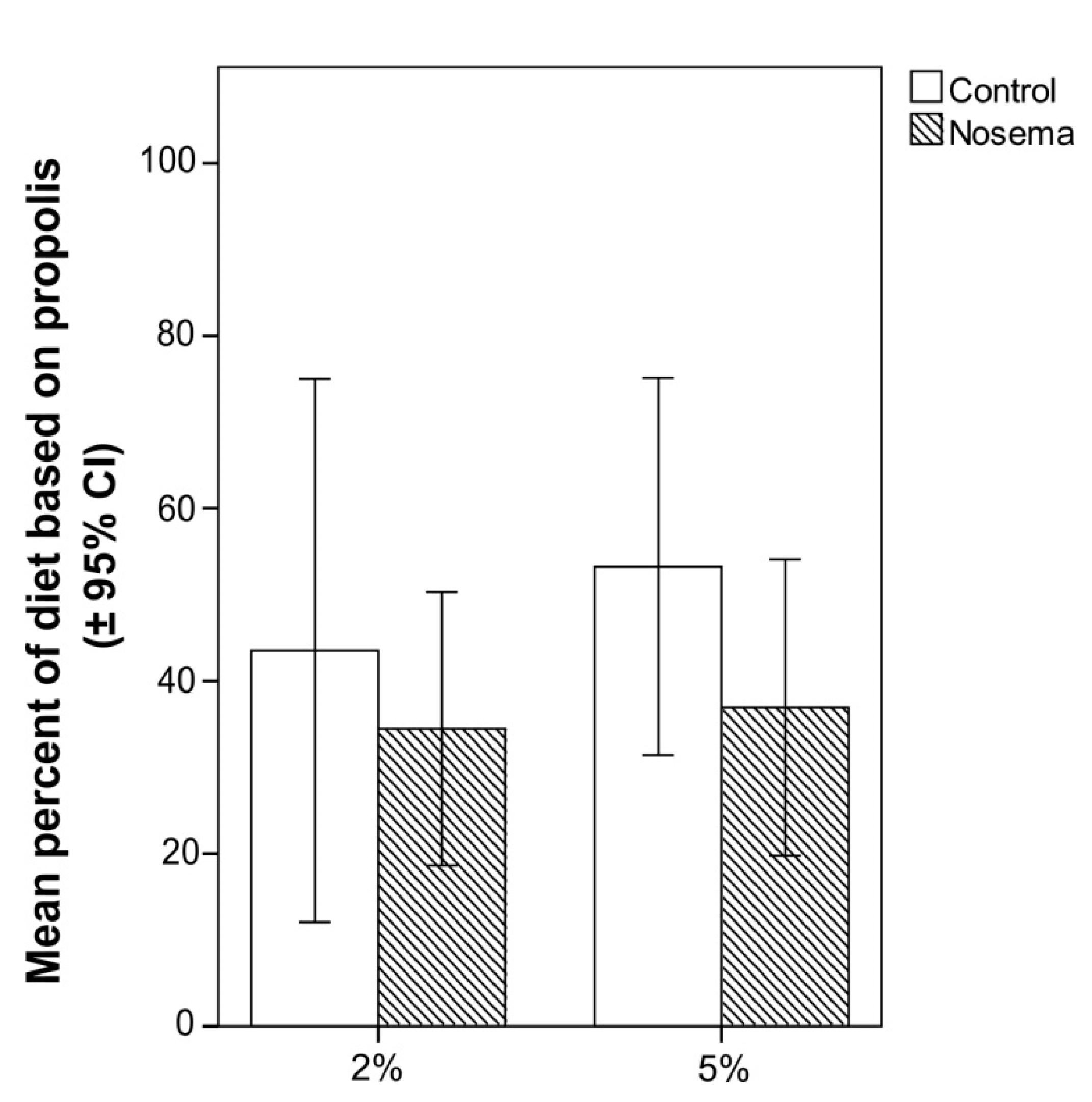
3.3. Scan Sampling and Food Consumption
3.4. Chemical Analysis of Propolis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fries, I.; Feng, F.; da Silva, A.; Slemenda, S.B.; Pieniazek, N.J. Nosema ceranae n. sp. (Microspora, Nosematidae), morphological and molecular characterization of a microsporidian parasite of the Asian honey bees Apis cerana (Hymenoptera, Apidae). Eur. J. Protistol. 1996, 32, 356–365. [Google Scholar] [CrossRef]
- Goblirsch, M. Nosema ceranae disease of the honey bee (Apis mellifera). Apidologie 2018, 49, 131–150. [Google Scholar] [CrossRef] [Green Version]
- Higes, M.; Martín, R.; Meana, A. Nosema ceranae, a new microsporidian parasite in honeybees in Europe. J. Invertebr. Pathol. 2006, 92, 93–95. [Google Scholar] [CrossRef] [PubMed]
- Ferroglio, E.; Zanet, S.; Peraldo, N.; Tachis, E.; Trisciuoglio, A.; Laurino, D.; Porporato, M. Nosema ceranae has been infecting honey bees Apis mellifera in Italy since at least 1993. J. Apic. Res. 2013, 52, 60–61. [Google Scholar] [CrossRef]
- Klee, J.; Besana, A.M.; Genersch, E.; Gisder, S.; Nanetti, A.; Tam, D.Q.; Chinh, T.X.; Puerta, F.; Ruz, J.M.; Kryger, P.; et al. Widespread dispersal of the microsporidian Nosema ceranae, an emergent pathogen of the western honey bee, Apis mellifera. J. Invertebr. Pathol. 2007, 96, 1–10. [Google Scholar] [CrossRef]
- Chen, Y.; Evans, J.D.; Smith, I.B.; Petti, J.S. Nosema ceranae is a long-present and wide-spread microsporidian infection of the European honey bee (Apis mellifera) in the United States. J. Invertebr. Pathol. 2008, 97, 186–188. [Google Scholar] [CrossRef]
- Porrini, C.; Mutinelli, F.; Bortolotti, L.; Granato, A.; Laurenson, L.; Roberts, K.; Gallina, A.; Silvester, N.; Medrzycki, P.; Renzi, T.; et al. The status of honey bee health in Italy: results from the Nationwide Bee Monitoring Network. PLoS ONE 2016, 1, e0155411. [Google Scholar] [CrossRef]
- Natsopoulou, M.E.; McMahon, D.P.; Doublet, W.; Bryden, J.; Paxton, R.J. Interspecific competition in honeybee intracellular gut parasites is asymmetric and favours the spread of an emerging infectious disease. Proc. R. Soc. B 2015, 282, 20141896. [Google Scholar] [CrossRef] [Green Version]
- Kralj, J.; Fuchs, S. Nosema sp. influences flight behavior of infected honey bee (Apis mellifera) foragers. Apidologie 2010, 41, 21–28. [Google Scholar] [CrossRef] [Green Version]
- Goblirsch, M.; Huang, Z.Y.; Spivak, M. Physiological and behavioral changes in honey bees (Apis mellifera) induced by Nosema ceranae infection. PLoS ONE 2013, 8, e58165. [Google Scholar] [CrossRef]
- Mayack, C.; Natsopoulou, M.E.; McMahon, D.P. Nosema ceranae alters a highly conserved hormonal stress pathway in honeybees. Insect. Mol. Biol. 2015, 24, 662–670. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, Y.; Cook, S.C. Chronic Nosema ceranae infection inflicts comprehensive and persistent immune suppression and accelerated lipid loss in host Apis mellifera honey bees. Int. J. Parasitol. 2018, 48, 433–444. [Google Scholar] [CrossRef] [PubMed]
- Botías, C.; Martín-Hernández, R.; Barrios, L.; Meana, A.; Higes, M. Nosema spp. infection and its negative effects on honey bees (Apis mellifera iberiensis) at the colony level. Vet. Res. 2013, 44, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higes, M.; Martín-Hernández, R.; Botías, C.; Bailón, E.G.; González-Porto, A.V.; Barrios, L.; del Nozal, M.J.; Bernal, J.L.; Jiménez, J.J. How natural infection by Nosema ceranae causes honeybee colony collapse. Environ. Microbiol. 2008, 10, 2659–2669. [Google Scholar] [CrossRef] [PubMed]
- Higes, M.; Martín-Hernández, R.; Garrido-Bailón, E.; González-Porto, A.V.; García-Palencia, P.; Meana, A.; Del Nozal, M.J.; Mayo, R.; Bernal, J.L. Honeybee colony collapse due to Nosema ceranae in professional apiaries. Environ. Microbiol. Rep. 2009, 1, 110–113. [Google Scholar] [CrossRef]
- Bromenshenk, J.J.; Henderson, C.B.; Wick, C.H.; Stanford, M.F.; Zulich, A.W.; Jabbour, R.E.; Deshpande, S.V.; Mc Cubbin, P.E.; Seccomb, R.A.; Welch, P.M.; et al. Iridovirus and microsporidian linked to honey bee colony decline. PLoS ONE 2010, 5, e13181. [Google Scholar] [CrossRef]
- Fries, I. Nosema ceranae in European honey bees (Apis mellifera). J. Invertebr. Pathol. 2010, 103, 73–79. [Google Scholar] [CrossRef]
- Sulborska, A.; Horecka, B.; Cebrat, M.; Kowalczyk, M.; Skrzypek, T.H.; Kazimierczak, W.; Trytek, M.; Borsuk, G. Microsporidia Nosema spp.—Obligate bee parasites are transmitted by air. Sci. Rep. 2019, 9, 14376. [Google Scholar] [CrossRef]
- van den Heever, J.P.; Thompson, T.S.; Curtis, J.M.; Ibrahim, A.; Pernal, S. Fumagillin: an overview of recent scientific advances and their significance for apiculture. J. Agric. Food. Chem. 2014, 62, 2728–2737. [Google Scholar] [CrossRef]
- Huang, W.F.; Solter, L.F.; Yau, P.M.; Imai, B.S. Nosema ceranae escapes fumagillin control in honey bees. PLoS Pathog. 2013, 9, e1003185. [Google Scholar] [CrossRef] [Green Version]
- Giacobino, A.; Rivero, R.; Molineri, A.I.; Bulacio Cagnolo, N.; Merke, J.; Orellano, E.; Salto, C.; Signorini, M. Fumagillin control of Nosema ceranae (Microsporidia: Nosematidae) infection in honey bee (Hymenoptera: Apidae) colonies in Argentina. Vet. Ital. 2016, 52, 145–151. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, Y.; Diaz-Cetti, S.; Ramallo, G.; Santos, E.; Porrini, M.; Invernizzi, C. Nosema ceranae winter control: study of the effectiveness of different fumagillin treatments and consequences on the strength of honey bee (Hymenoptera: Apidae) colonies. J. Econ. Entomol. 2017, 110, 1–5. [Google Scholar] [CrossRef] [PubMed]
- MRL, Commission regulation EU 2010 n. 37/2010 of 22 December 2009. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A32010R0037 (accessed on 10 December 2019).
- Maistrello, L.; Lodesani, M.; Costa, C.; Leonardi, F.; Marani, G.; Caldon, M.; Mutinelli, F.; Granato, A. Screening of natural compounds for the control of nosema disease in honeybees (Apis mellifera). Apidologie 2008, 39, 436–445. [Google Scholar] [CrossRef] [Green Version]
- Porrini, M.P.; Fernández, N.J.; Garrido, P.M.; Gende, L.B.; Medici, S.K.; Eguaras, M.J. In vivo evaluation of antiparasitic activity of plant extracts on Nosema ceranae (Microsporidia). Apidologie 2011, 42, 700–707. [Google Scholar] [CrossRef] [Green Version]
- Porrini, M.P.; Garrido, P.M.; Gende, L.B.; Rossini, C.; Hermidac, L.; Marcángelia, J.A.; Eguaras, M.J. Oral administration of essential oils and main components: study on honey bee survival and Nosema ceranae development. J. Apic. Res. 2017, 56, 616–624. [Google Scholar] [CrossRef]
- Burnham, A.J. Scientific advances in controlling Nosema ceranae (Microsporidia) infections in honey bees (Apis mellifera). Front. Vet. Sci. 2019, 6, 79. [Google Scholar] [CrossRef] [Green Version]
- Costa, C.; Lodesani, M.; Maistrello, L. Effect of thymol and resveratrol administered with candy or syrup on the development of Nosema ceranae and on the longevity of honeybees (Apis mellifera L.) in laboratory conditions. Apidologie 2010, 41, 141–150. [Google Scholar] [CrossRef] [Green Version]
- Damiani, N.; Fernández, N.J.; Porrini, M.P.; Gende, L.B.; Álvarez, E.; Buffa, F.; Brasesco, C.; Maggi, M.D.; Marcangeli, J.A.; Eguaras, M.J. Laurel leaf extracts for honeybee pest and disease management: antimicrobial, microsporicidal, and acaricidal activity. Parasitol. Res. 2014, 113, 701–709. [Google Scholar] [CrossRef]
- Bravo, J.; Carbonell, V.; Sepúlveda, B.; Delporte, C.; Valdovinos, C.E.; Martín-Hernández, R.; Higes, M. Antifungal activity of the essential oil obtained from Cryptocarya alba against infection in honey bees by Nosema ceranae. J. Invertebr. Pathol. 2017, 149, 141–147. [Google Scholar] [CrossRef]
- Erler, S.; Moritz, R.F. Pharmacophagy and pharmacophory: mechanisms of self-medication and disease prevention in the honeybee colony (Apis mellifera). Apidologie 2016, 47, 389–411. [Google Scholar] [CrossRef] [Green Version]
- Seeley, T.D.; Morse, R.A. The nest of the honey bee (Apis mellifera L.). Insectes Soc. 1976, 23, 495–512. [Google Scholar] [CrossRef]
- Castella, G.; Chapuisat, M.; Christe, P. Prophylaxis with resin in wood ants. Anim. Behav. 2008, 75, 1591–1596. [Google Scholar] [CrossRef] [Green Version]
- Leonhardt, S.D.; Bluthgen, N. A sticky affair: resin collection by Bornean stingless bees. Biotropica 2009, 41, 730–736. [Google Scholar] [CrossRef]
- Simone-Finstrom, M.; Spivak, M. Propolis and bee health: the natural history and significance of resin use by honey bees. Apidologie 2010, 41, 295–311. [Google Scholar] [CrossRef] [Green Version]
- Kadhim, M.J.; Los, A.; Olszewski, K.; Borsuk, G. Propolis in livestock nutrition. Entomol. Ornithol. Herpetol. 2018, 7, 207. [Google Scholar] [CrossRef]
- Küstenmacher, M. Propolis. Ber. Deutshen Pharmacol. Ges. 1911, 21, 65–92. [Google Scholar] [CrossRef]
- Ghisalberti, E.L. Propolis: a review. Bee World 1979, 60, 59–84. [Google Scholar] [CrossRef]
- Simone, M.; Evans, J.D.; Spivak, M. Resin collection and social immunity in honey bees. Evolution 2009, 63, 3016–3022. [Google Scholar] [CrossRef]
- Cotter, S.C.; Kruuk, L.E.; Wilson, K. Costs of resistance: genetic correlations and potential trade-offs in an insect immune system. J. Evol. Biol. 2004, 17, 421–429. [Google Scholar] [CrossRef] [Green Version]
- Nicodemo, D.; Malheiros, E.B.; De Jong, D.; Couto, R.H.N. Honey bee lines selected for high propolis production also have superior hygienic behavior and increased honey and pollen stores. Genet. Mol. Res. 2013, 12, 6931–6938. [Google Scholar] [CrossRef]
- Borba, R.S.; Klyczck, K.K.; Mogen, K.L.; Spivak, M. Seasonal benefits of a natural propolis envelope to honey bee immunity and colony health. J. Exp. Biol. 2015, 218, 3689–3699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pusceddu, M.; Floris, I.; Mura, A.; Theodorou, P.; Cirotto, G.; Piluzza, S.; Bullitta, S.; Angioni, A.; Satta, A. The effects of raw propolis on Varroa-infested honey bee (Apis mellifera) workers. Parasitol. Res. 2018, 117, 3527–3535. [Google Scholar] [CrossRef] [PubMed]
- Antúnez, K.; Harriet, J.; Gende, L.; Maggi, M.; Eguaras, M.; Zunino, P. Efficacy of natural propolis extract in the control of American Foulbrood. Vet. Microbiol. 2008, 131, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Borba, R.S.; Spivak, M. Propolis envelope in Apis mellifera colonies supports honey bees against the pathogen, Paenibacillus larvae. Sci. Rep. 2017, 7, 11429. [Google Scholar] [CrossRef] [Green Version]
- Simone-Finstrom, M.D.; Spivak, M. Increased resin collection after parasite challenge: A case of self-medication in honey bees? PLoS ONE 2012, 7, e34601. [Google Scholar] [CrossRef] [Green Version]
- Pusceddu, M.; Piluzza, S.; Theodorou, P.; Buffa, F.; Ruiu, L.; Bullitta, S.; Floris, I.; Satta, A. Resin foraging dynamics in Varroa destructor-infested hives: A case of medication of kin? Insect Sci. 2019, 26, 297–310. [Google Scholar] [CrossRef]
- Drescher, N.; Klein, A.M.; Neumann, P.; Yañez, O.; Leonhardt, S. Inside honeybee hives: impact of natural propolis on the ectoparasitic mite Varroa destructor and viruses. Insects 2017, 8, 15. [Google Scholar] [CrossRef] [Green Version]
- Gherman, B.I.; Denner, A.; Bobiş, O.; Dezmirean, D.S.; Mărghitaş, L.A.; Schluens, H.; Moritz, R.F.; Erler, S. Pathogen-associated self-medication behavior in the honeybee Apis mellifera. Behav. Ecol. Sociobiol. 2014, 68, 1777–1784. [Google Scholar] [CrossRef] [Green Version]
- Turcatto, A.P.; Lourenço, A.P.; De Jong, D. Propolis consumption ramps up the immune response in honey bees infected with bacteria. Apidologie 2018, 49, 287–296. [Google Scholar] [CrossRef] [Green Version]
- Damiani, N.; Maggi, M.D.; Gende, L.B.; Faverin, C.; Eguaras, M.J.; Marcangeli, J.A. Evaluation of the toxicity of a propolis extract on Varroa destructor (Acari: Varroidae) and Apis mellifera (Hymenoptera: Apidae). J. Apic. Res. 2010, 49, 257–264. [Google Scholar] [CrossRef]
- Damiani, N.; Porrini, M.P.; Lancia, J.P.; Álvarez, E.; Garrido, P.M.; Domínguez, E.; Gende, L.B.; Eguaras, M.J. Effect of propolis oral intake on physiological condition of young worker honey bees, Apis mellifera L. J. Apic. Sci. 2017, 61, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Johnson, R.; Mao, W.; Pollock, H.; Niu, G.; Schuler, M.; Berenbaum, M. Ecologically appropriate xenobiotics induce cytochrome P450s in Apis mellifera. PLoS ONE 2012, 7, e31051. [Google Scholar] [CrossRef] [PubMed]
- Suwannapong, G.; Maksong, S.; Benbow, M.E. Stingless bee propolis effects on experimental infection of Apis florea with Nosema ceranae. J. Agr. Sci. Technol. A 2011, 1, 818–825. [Google Scholar]
- Suwannapong, G.; Maksong, S.; Phainchajoen, M.; Benbow, M.E.; Mayack, C. Survival and health improvement of Nosema infected Apis florea (Hymenoptera: Apidae) bees after treatment with propolis extract. J. Asia Pac. Entomol. 2018, 21, 437–444. [Google Scholar] [CrossRef]
- Yemor, T.; Phiancharoen, M.; Benbow, M.E.; Suwannapomg, G. Effects of stingless bee propolis on Nosema ceranae infected Asian honey bees, Apis cerana. J. Apic. Res. 2015, 54, 468–473. [Google Scholar] [CrossRef]
- Arismendi, N.; Vargas, M.; López, M.D.; Barría, Y.; Zapata, N. Promising antimicrobial activity against the honey bee parasite Nosema ceranae by methanolic extracts from Chilean native plants and propolis. J. Apic. Res. 2018, 57, 522–535. [Google Scholar] [CrossRef]
- da Silva, J.F.M.; de Souza, M.C.; Matta, S.R.; de Andrade, M.R.; Vidal, F.V.N. Correlation analysis between phenolic levels of Brazilian propolis extracts and their antimicrobial and antioxidant activities. Food. Chem. 2006, 99, 431–435. [Google Scholar] [CrossRef]
- Mihai, C.; Mărghitaş, L.; Dezmirean, D.; Chirilă, F.; Moritz, R.; Schlüns, H. Interactions among flavonoide of propolis affect antibacterial activity against the honeybee pathogen Paenibacillus larvae. J. Invertebr. Pathol. 2012, 110, 68–72. [Google Scholar] [CrossRef]
- Siripatrawan, U.; Vitchayakitti, W.; Sanguandeekul, R. Antioxidant and antimicrobial properties of Thai propolis extracted using ethanol aqueous solution. Int. J. Food. Sci. Technol. 2013, 48, 22–27. [Google Scholar] [CrossRef]
- Bagella, S.; Urbani, M. La flora degli affioramenti calcarei miocenici della Sardegna settentrionale. Giornale Botanico Italiano 1994, 128, 1–370. [Google Scholar] [CrossRef] [Green Version]
- Biondi, E.; Filigheddu, R.S.; Farris, E. II Paesaggio vegetale della Nurra (Sardegna nord-occidentale). Pavia Società Italiana di Fitosociologia 2001, 38, 3–105. Available online: http://eprints.uniss.it/3178/ (accessed on 10 December 2019).
- Fries, I.; Chauzat, M.P.; Chen, Y.P.; Doublet, V.; Genersch, E.; Gisder, S.; Higes, M.; McMahon, D.P.; Martín-Hernández, R.; Natsopoulou, M.E.; et al. Standard methods for Nosema research. J. Apic. Res. 2013, 52, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Damiani, N.; Fernández, N.J.; Maldonado, L.M.; Álvarez, A.R.; Eguaras, M.J.; Marcangeli, J.A. Bioactivity of propolis from different geographical origins on Varroa destructor (Acari: Varroidae). Parasitol. Res. 2010, 107, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Malone, L.A.; Stefanovic, D. Comparison of the responses of two races of honeybees to infection with Nosema apis Zander. Apidologie 1999, 30, 375–382. [Google Scholar] [CrossRef] [Green Version]
- Forsgren, E.; Fries, I. Comparative virulence of Nosema ceranae and Nosema apis in individual European honey bees. Vet. Parasitol. 2010, 170, 212–217. [Google Scholar] [CrossRef]
- Williams, G.R.; Alaux, C.; Costa, C.; Csáki, T.; Doublet, V.; Eisenhardt, D.; Fries, I.; Kuhn, R.; McMahon, D.P.; Medrzycki, P.; et al. Standard methods for maintaining adult Apis mellifera in cages under in vitro laboratory conditions. J. Apic. Res. 2013, 52, 1–36. [Google Scholar] [CrossRef] [Green Version]
- Higes, M.; García-Palencia, P.; Martín-Hernández, R.; Meana, A. Experimental infection of Apis mellifera honeybees with Nosema ceranae (microsporidia). J. Invertebr. Pathol. 2007, 94, 211–217. [Google Scholar] [CrossRef]
- van Engelsdorp, D.; Evans, J.D.; Saegerman, C.; Mullin, C.; Haubruge, E.; Nguyen, B.K.; Frazier, M.; Frazier, J.; Cox-Foster, D.; Chen, Y.; et al. Colony Collapse Disorder: a descriptive study. PLoS ONE 2009, 4, e6481. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphor-molybdic-phosphotungstic acid reagents. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Therneau, T.M. Coxme: Mixed Effects Cox Models. R Package Version 2.2-10. 2018. Available online: https://CRAN.R-project.org/package=coxme (accessed on 10 December 2019).
- Therneau, T.M.; Grambsch, P.M. Modeling Survival Data: Extending the Cox Model; Springer: New York, NY, USA, 2000; ISBN 0-387-98784-3. [Google Scholar]
- Kassambara, A.; Kosinski, M. Survminer: Drawing survival curves using ‘ggplot2’. R package version 0.4.3. 2018. Available online: https://CRAN.R-project.org/package=survminer (accessed on 10 December 2019).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 10 December 2019).
- Sridevi, V.; Chandana Lakshmi, M.V.V.; Manasa, M.; Sravani, M. Metabolic pathways for the biodegradation of phenol. Int. J. Eng. Sci. Adv. Technol. 2012, 2, 695–705. Available online: www.ijesat.org (accessed on 10 December 2019).
- Suwannapong, G.; Maksong, S.; Seanbualuang, P.; Benbow, M.E. Experimental infection of red dwarf honeybee, Apis florea, with Nosema ceranae. J. Asia Pac. Entom. 2010, 13, 361–364. [Google Scholar] [CrossRef]
- Ptaszynska, A.A.; Borsuk, G.; Mulenko, W.; Olszewski, K. Impact of ethanol on Nosema spp. infected bees. Med. Veter. 2013, 69, 736–740. [Google Scholar]
- Maze, I.S.; Wright, G.A.; Mustard, J.A. Acute ethanol ingestion produces dose-dependent effects on motor behavior in the honey bee (Apis mellifera). J. Insect. Physiol. 2006, 52, 1243–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonnell, G.; Russell, A.D. Antiseptics and disinfectants: activity, action, and resistance. Clin. Microbiol. Rev. 1999, 12, 147–179. [Google Scholar] [CrossRef] [Green Version]
- Sequeira, S.O.; Phillips, A.J.; Cabrita, E.J.; Macedo, M.F. Ethanol as an antifungal treatment for paper: short-term and long-term effects. Stud. Conserv. 2017, 62, 33–42. [Google Scholar] [CrossRef]
- Lindenfelser, L. Antimicrobial activity of propolis. Am. Bee. J. 1967, 90–131. [Google Scholar]
- Wilson, M.B.; Brinkman, D.; Spivak, M.; Gardner, G.; Cohen, J.D. Regional variation in composition and antimicrobial activity of U.S. propolis against Paenibacillus larvae and Ascosphaera apis. J. Invertebr. Pathol. 2015, 124, 44e50. [Google Scholar] [CrossRef]
- Bankova, V.S.; Popov, S.S.; Marekov, N.L. A study on flavonoids of propolis. J. Nat. Prods. 1983, 46, 71–474. [Google Scholar] [CrossRef]
- Bilikova, K.; Popova, M.; Trusheva, B.; Bankova, V. New anti-Paenibacillus larvae substances purified from propolis. Apidologie 2012, 44, 278–285. [Google Scholar] [CrossRef] [Green Version]
- Voight, K.; Rademacher, E. Effect of the propolis components, cinnamic acid and pinocembrin, on Apis mellifera and Ascophaera apis. J. Apic. Sci. 2015, 59, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Wilson, M.B.; Pawlus, A.D.; Brinkman, D.; Gardner, G.; Hegeman, A.D.; Spivak, M.; Cohen, J.D. 3-Acyl dihydroflavonols from poplar resins collected by honey bees are active against the bee pathogens Paenibacillus larvae and Ascosphaera apis. Phytochemistry 2017, 138, 83–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Park, J.K.; Lee, J.K. Evaluation of antimicrosporidian activity of plant extracts on Nosema ceranae. J. Apic. Sci. 2016, 60, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Mocan, A.; Crisan, G.; Vlase, L.; Crisan, O.; Vodnar, D.C.; Raita, O.; Gheldiu, A.M.; Toiu, A.; Oprean, R.; Tilea, I. Comparative studies on polyphenolic composition, antioxidant and antimicrobial activities of Schisandra chinensis leaves and fruits. Molecules 2014, 19, 15162–15179. [Google Scholar] [CrossRef] [Green Version]
- Cushnie, T.P.T.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef]
- Kujumgiev, A.; Tsvetkova, I.; Serkedjieva, Y.; Bankova, V.; Christov, R.; Popov, S. Antibacterial, antifungal and antiviral activity of propolis of different geographic origin. J. Ethnopharmacol. 1999, 64, 235–240. [Google Scholar] [CrossRef]
- Ptaszyńska, A.A.; Trytek, M.; Borsuk, G.; Buczek, K.; Rybicka-Jasińska, K.; Gryko, D. Porphyrins inactivate Nosema spp. microsporidia. Sci. Rep. 2018, 8, 5523. [Google Scholar] [CrossRef]
- Spivak, M.; Goblirsch, M.; Simone-Finstrom, M. Social-medication in bees: the line between individual and social regulation. Curr. Opin. Insect Sci. 2019, 33, 49–55. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | mg/g ± RSD% x |
|---|---|
| Caffeic acid y | 31.04 ± 5.01 |
| Caffeic acid derivatives z | 113.17 ± 12.35 |
| Paracumaric acid y | 7.12 ± 3.68 |
| Ferulic acid y | 7.62 ± 5.25 |
| Ferulic acid derivatives z | 122.70 ± 14.68 |
| Narirutin y | 1.37 ± 6.51 |
| Cinnamic acid derivatives z | 82.04 ± 10.78 |
| Quercetin y | 10.83 ± 3.45 |
| Quercetin derivatives z | 238.75 ± 6.59 |
| Luteolin y | 3.00 ± 5.68 |
| Naringin y | 59.01 ± 10.45 |
| Naringin derivatives z | 21.99 ± 1.56 |
| Kaempherol y | 34.01 ± 5.78 |
| OH-Flavone derivatives z | 17.21 ± 2.56 |
| Galangina y | 15.05 ± 2.48 |
| Rosmarinic acid derivatives z | 44.84 ± 13.98 |
| Ellagic acid derivatives z | 125.44 ± 4.65 |
| Quinic acid y | 1.39 ± 8.95 |
| Quinic acid derivatives z | 4.01 ± 6.01 |
| Total Phenols * | 119.77 ± 16.05 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mura, A.; Pusceddu, M.; Theodorou, P.; Angioni, A.; Floris, I.; Paxton, R.J.; Satta, A. Propolis Consumption Reduces Nosema ceranae Infection of European Honey Bees (Apis mellifera). Insects 2020, 11, 124. https://doi.org/10.3390/insects11020124
Mura A, Pusceddu M, Theodorou P, Angioni A, Floris I, Paxton RJ, Satta A. Propolis Consumption Reduces Nosema ceranae Infection of European Honey Bees (Apis mellifera). Insects. 2020; 11(2):124. https://doi.org/10.3390/insects11020124
Chicago/Turabian StyleMura, Alessandra, Michelina Pusceddu, Panagiotis Theodorou, Alberto Angioni, Ignazio Floris, Robert J. Paxton, and Alberto Satta. 2020. "Propolis Consumption Reduces Nosema ceranae Infection of European Honey Bees (Apis mellifera)" Insects 11, no. 2: 124. https://doi.org/10.3390/insects11020124
APA StyleMura, A., Pusceddu, M., Theodorou, P., Angioni, A., Floris, I., Paxton, R. J., & Satta, A. (2020). Propolis Consumption Reduces Nosema ceranae Infection of European Honey Bees (Apis mellifera). Insects, 11(2), 124. https://doi.org/10.3390/insects11020124