Larvicidal, Ovicidal, Synergistic, and Repellent Activities of Sophora alopecuroides and Its Dominant Constituents Against Aedes albopictus

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Insect Collection and Rearing
2.2. Plant Materials
Botanical Formation
2.3. Larvicidal, Ovicidal, and Repellent Activities
Larvicidal and Ovicidal Activity of the S. alopecuroides Extract and Its Constituents
2.4. Evaluation of the Synergistic/Antagonistic Relation between the Constituents of S. alopecuroides Extract
2.5. Adult Repellent Activity of the S. alopecuroides Extract
2.5.1. Preparation of Adult Females
2.5.2. Bioassay
2.6. Statistical Analysis
3. Results
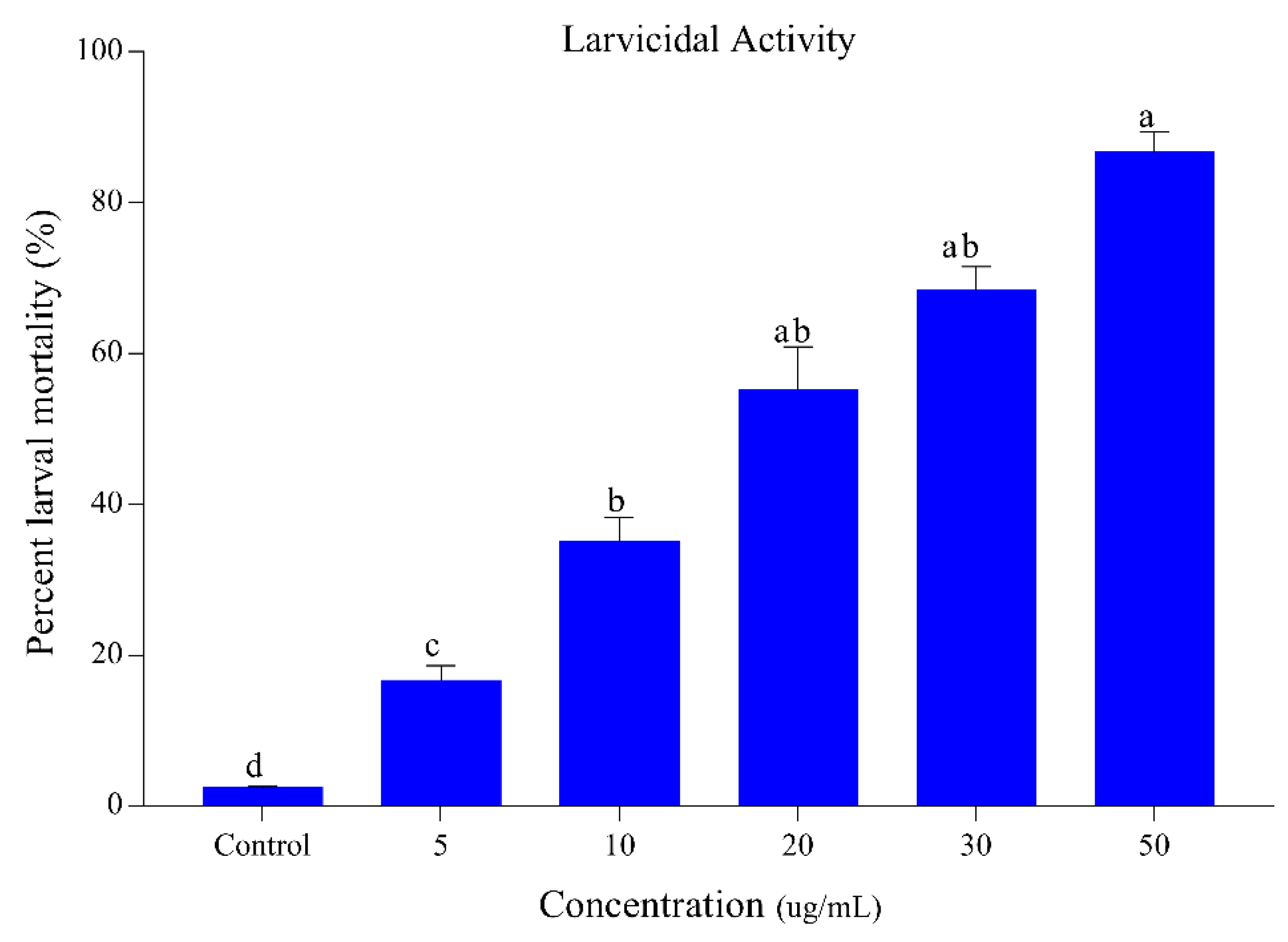
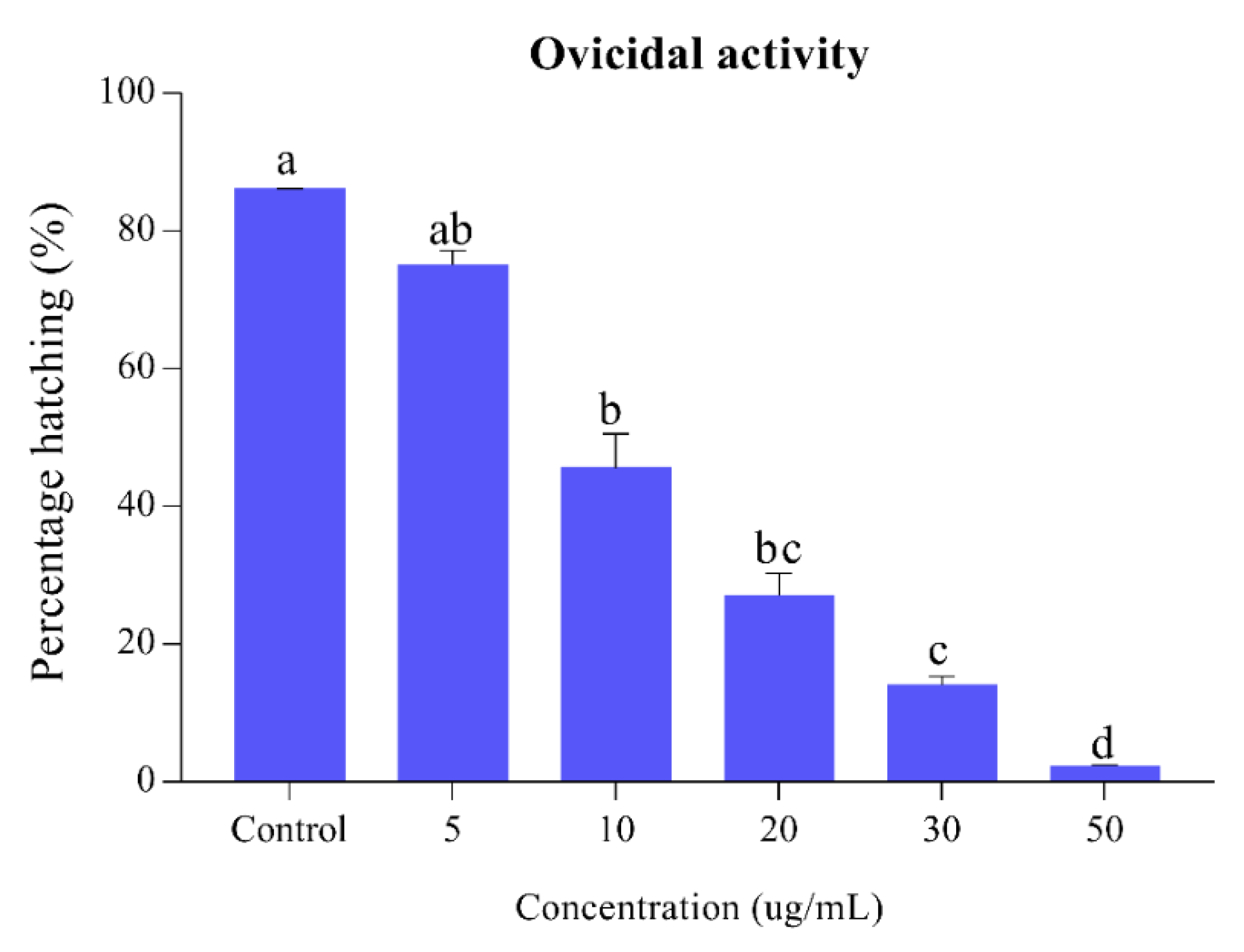
3.1. Larvicidal and Ovicidal Activity of Sophora Alopecuroides Extract and Its Constituents
3.1.1. Larvicidal and Ovicidal Activity of Sophora Alopecuroides Extract
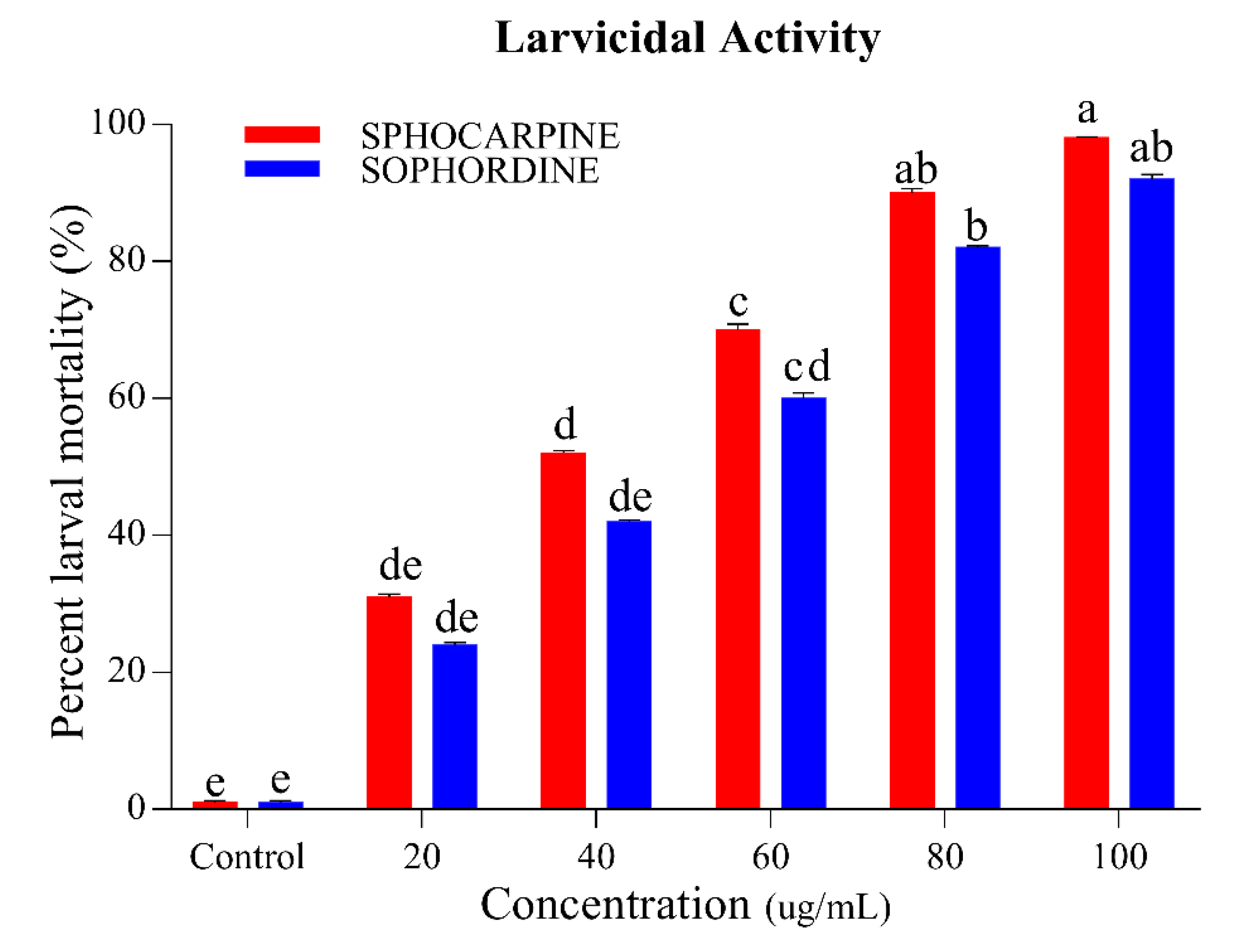
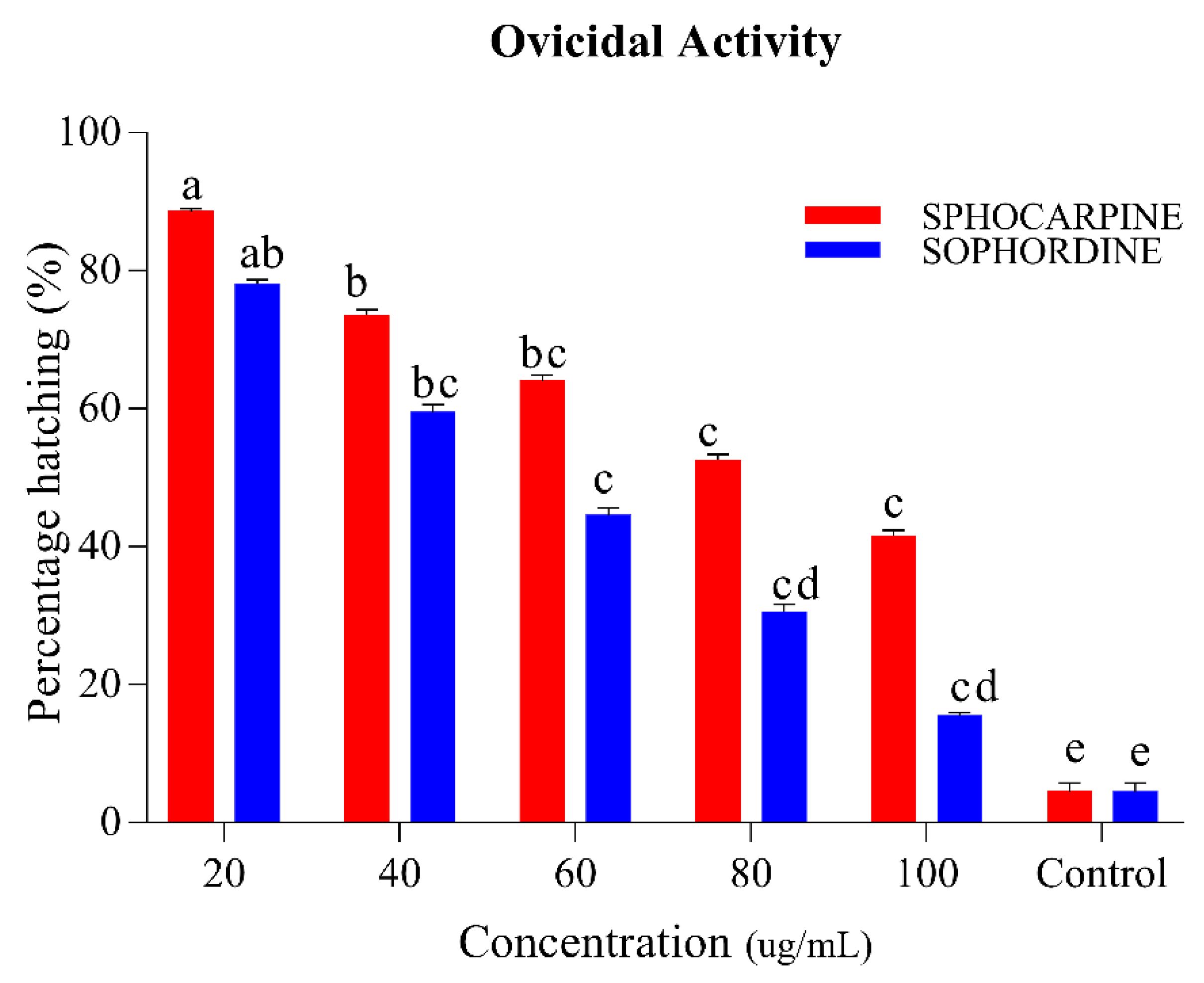
3.1.2. Larvicidal and Ovicidal Activity of Sophocarpin and Sophordine.
3.2. Evaluation of Synergistic/Antagonistic Relation between the Constituents of S. alopecuroides Extract
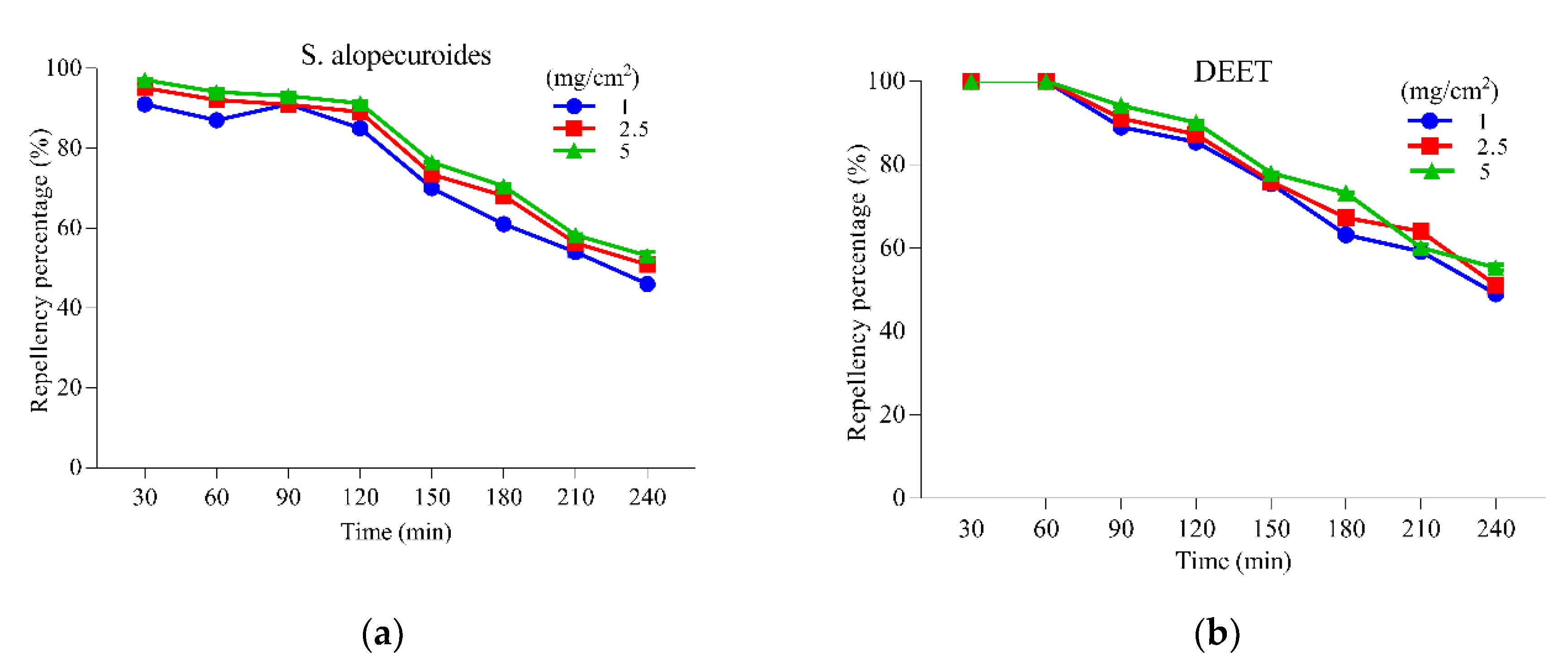
3.3. Adult Repellent Activity of the S. alopecuroides Extract
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gratz, N. Critical review of the vector status of Aedes albopictus. Med. Vet. Entomol. 2004, 18, 215–227. [Google Scholar] [CrossRef] [PubMed]
- Enserink, M. A mosquito goes global. Science 2008, 320, 864–866. [Google Scholar] [CrossRef] [PubMed]
- Chouin-Carneiro, T.; Vega-Rua, A.; Vazeille, M.; Yebakima, A.; Girod, R.; Goindin, D.; Dupont-Rouzeyrol, M.; Lourenço-de-Oliveira, R.; Failloux, A.-B. Differential susceptibilities of Aedes aegypti and Aedes albopictus from the Americas to Zika virus. PLoS Negl. Trop. Dis. 2016, 10, e0004543. [Google Scholar] [CrossRef] [PubMed]
- Rochlin, I.; Ninivaggi, D.V.; Hutchinson, M.L.; Farajollahi, A. Climate change and range expansion of the Asian tiger mosquito (Aedes albopictus) in Northeastern USA: Implications for public health practitioners. PLoS ONE 2013, 8, e60874. [Google Scholar] [CrossRef]
- Vega-Rua, A.; Zouache, K.; Caro, V.; Diancourt, L.; Delaunay, P.; Grandadam, M.; Failloux, A.-B. High efficiency of temperate Aedes albopictus to transmit chikungunya and dengue viruses in the Southeast of France. PLoS ONE 2013, 8, e59716. [Google Scholar] [CrossRef] [Green Version]
- Hanson, S.M.; Craig, G.B., Jr. Cold acclimation, diapause, and geographic origin affect cold hardiness in eggs of Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 1994, 31, 192–201. [Google Scholar] [CrossRef]
- Wu, J.Y.; Lun, Z.R.; James, A.A.; Chen, X.G. Dengue fever in mainland China. Am. J. Trop. Med. Hyg. 2010, 83, 664–671. [Google Scholar] [CrossRef] [Green Version]
- Medlock, J.M.; Hansford, K.M.; Schaffner, F.; Versteirt, V.; Hendrickx, G.; Zeller, H.; Bortel, W.V. A review of the invasive mosquitoes in Europe: Ecology, public health risks, and control options. Vector-Borne Zoonotic Dis. 2012, 12, 435–447. [Google Scholar] [CrossRef] [Green Version]
- Shoukat, R.F.; Hassan, B.; Shakeel, M.; Zafar, J.; Li, S.; Freed, S.; Xu, X.; Jin, F. Pathogenicity and Transgenerational Effects of Metarhizium anisopliae on the Demographic Parameters of Aedes albopictus (Culicidae: Diptera). J. Med. Entomol. 2019. [Google Scholar] [CrossRef]
- Shakeel, M.; Farooq, M.; Nasim, W.; Akram, W.; Khan, F.Z.A.; Jaleel, W.; Zhu, X.; Yin, H.; Li, S.; Fahad, S. Environment polluting conventional chemical control compared to an environmentally friendly IPM approach for control of diamondback moth, Plutella xylostella (L.), in China: A review. Environ. Sci. Pollut. Res. 2017, 24, 14537–14550. [Google Scholar] [CrossRef]
- Ocampo, C.B.; Salazar-Terreros, M.J.; Mina, N.J.; McAllister, J.; Brogdon, W. Insecticide resistance status of Aedes aegypti in 10 localities in Colombia. Acta Trop. 2011, 118, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Miranda, J.E.; Navickiene, H.M.D.; Nogueira-Couto, R.H.; De Bortoli, S.A.; Kato, M.J.; da Silva Bolzani, V.; Furlan, M. Susceptibility of Apis mellifera (Hymenoptera: Apidae) to pellitorine, an amide isolated from Piper tuberculatum (Piperaceae). Apidologie 2003, 34, 409–415. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-Y.; Wu, D.-C.; Yu, J.-Z.; Chen, B.-H.; Wang, C.-L.; Ko, W.-H. Control of silverleaf whitefly, cotton aphid and kanzawa spider mite with oil and extracts from seeds of sugar apple. Neotrop. Entomol. 2009, 38, 531–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoukat, R.F.; Freed, S.; Ahmad, K.W.; Rehman, A.-U. Assessment of Binary Mixtures of Entomopathogenic Fungi and Chemical Insecticides on Biological Parameters of Culex pipiens (Diptera: Culicidae) under Laboratory and Field Conditions. Pak. J. Zool. 2018, 50, 299–309. [Google Scholar] [CrossRef]
- Malik, S.U.; Zia, K.; Ajmal, M.; Shoukat, R.F.; Li, S.; Saeed, M.; Zafar, J.; Shoukat, R.F. Comparative efficacy of different insecticides and estimation of yield losses on BT and non-BT cotton for thrips, red cotton bug, and dusky cotton bug. J. Entomol. Zool. Stud. 2018, 6, 505–512. [Google Scholar]
- Zafar, J.; Freed, S.; Khan, B.A.; Farooq, M. Effectiveness of Beauveria bassiana Against Cotton Whitefly, Bemisia tabaci (Gennadius) (Aleyrodidae: Homoptera) on Different Host Plants. Pak. J. Zool. 2016, 48, 91–99. [Google Scholar]
- Ahmed, K.; Freed, S.; Shoukat, R.F.; Ahmad, K.W. Efficacy of Entomopathogenic Fungi with Insecticides Mixtures against Oxycarenus hyalinipennis (Costa) (Lygaeidae: Hemiptera). Pak. J. Zool 2020, 52, 573–583. [Google Scholar] [CrossRef]
- Ahmed, S.M.; Saeed, M.; Nawaz, A.; Usman, M.; Shoukat, R.F.; Li, S.; Zhang, Y.; Zeng, L.; Zafar, J.; Akash, A. Monitoring of quantitative and qualitative losses by lepidopteran, and homopteran pests in different crop production systems of Brassica oleracea L. J. Entomol. Zool. Stud. 2018, 6, 06–12. [Google Scholar]
- Enayati, A.A.; Hemingway, J. Pyrethroid insecticide resistance and treated bednets efficacy in malaria control. Pest. Biochem. Physiol. 2006, 84, 116–126. [Google Scholar] [CrossRef]
- Shoukat, R.F.; Zafar, J.; Shakeel, M.; Zhang, Y.; Freed, S.; Xu, X.; Jin, F. Assessment of Lethal, Sublethal, and Transgenerational Effects of Beauveria Bassiana on the Demography of Aedes albopictus (Culicidae: Diptera). Insects 2020, 11, 178. [Google Scholar] [CrossRef] [Green Version]
- Mdoe, F.P.; Cheng, S.-S.; Lyaruu, L.; Nkwengulila, G.; Chang, S.-T.; Kweka, E.J. Larvicidal efficacy of Cryptomeria japonica leaf essential oils against Anopheles gambiae. Parasite Vector 2014, 7, 426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shoukat, R.F.; Freed, S.; Ahmad, K.W. Evaluation of binary mixtures of entomogenous fungus and botanicals on biological parameters of Culex pipiens (Diptera: Culicidae) under laboratory and field conditions. Int. J. Mosq. Res. 2016, 3, 17–24. [Google Scholar]
- Park, Y.-L.; Tak, J.-H. Essential oils for arthropod pest management in agricultural production systems. In Essential Oils in Food Preservation. Flavor and Safety; Elsevier: San Diego, CA, USA, 2016; pp. 61–70. [Google Scholar]
- Khan, B.A.; Freed, S.; Zafar, J.; Farooq, M.; Shoukat, R.F.; Ahmad, K.W.; Li, S.; Zhang, Y.; Hua, Y.; Shoukat, R.F. Efficacy of different entomopathogenic fungi on biological parameters of pulse beetle Callosobruchus chinensis L. (Coleoptera: Bruchidae). J. Entomol. Zool. Stud. 2018, 6, 1972–1976. [Google Scholar]
- Bakkali, F.; Averbeck, S.; Averbeck, D.; Idaomar, M. Biological effects of essential oils–A review. Food Chem. Toxicol. 2008, 46, 446–475. [Google Scholar] [CrossRef]
- Rizvi, S.A.H.; Tao, L.; Zeng, X. Chemical composition of essential oil obtained from Artemesia absinthium L. grown under the climatic condition of Skardu Baltistan of Pakistan. Pak. J. Bot. 2018, 50, 599–604. [Google Scholar]
- Rizvi, S.A.H.; Ling, S.; Tian, F.; Liu, J.; Zeng, X. Interference mechanism of Sophora alopecuroides L. alkaloids extract on host finding and selection of the Asian citrus psyllid Diaphorina citri Kuwayama (Hemiptera: Psyllidae). Environ. Sci. Pollut. Res. 2018. [Google Scholar] [CrossRef]
- Rizvi, S.A.H.; Siquan, L.; Fajun, T.; Feng, X.; Xinnian, Z. Toxicity and enzyme inhibition activities of the essential oil and dominant constituents derived from Artemisia absinthium L. against adult Asian citrus psyllid Diaphorina citri Kuwayama (Hemiptera: Psyllidae). Ind. Crop. Prod. 2018, 121, 468–475. [Google Scholar] [CrossRef]
- Joshi, R.K. Volatile composition and antimicrobial activity of the essential oil of Artemisia absinthium growing in Western Ghats region of North West Karnataka, India. Pharm. Biol. 2013, 51, 888–892. [Google Scholar] [CrossRef]
- Sukumar, K.; Perich, M.J.; Boobar, L.R. Botanical derivatives in mosquito control: A review. J. Am. Mosq. Control Assoc. 1991, 7, 210–237. [Google Scholar]
- Saeed, M.; Shoukat, R.F.; Zafar, J. Population dynamics of natural enemies and insect pest in different Brassica oleracea (cabbage) growing seasons with different production systems. J. Entomol. Zool. Stud. 2017, 5, 1669–1674. [Google Scholar]
- Su, T.; Mulla, M.S. Oviposition bioassay responses of Culex tarsalis and Culex quinquefasciatus to neem products containing azadirachtin. Entomol. Exp. Appl. 1999, 91, 337–345. [Google Scholar] [CrossRef]
- Kumar, S.; Mishra, M.; Wahab, N.; Warikoo, R. Larvicidal, repellent, and irritant potential of the seed-derived essential oil of Apium graveolens against dengue vector, Aedes aegypti L. (Diptera: Culicidae). Front. Public Health 2014, 2, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, A.; Mazigo, H.D.; Manjurano, A.; Morona, D.; Kweka, E.J. Evaluation of active ingredients and larvicidal activity of clove and cinnamon essential oils against Anopheles gambiae (sensu lato). Parasite Vector 2017, 10, 411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrade-Ochoa, S.; Sánchez-Aldana, D.; Chacón-Vargas, K.F.; Rivera-Chavira, B.E.; Sánchez-Torres, L.E.; Camacho, A.D.; Nogueda-Torres, B.; Nevárez-Moorillón, G.V. Oviposition deterrent and larvicidal and pupaecidal activity of seven essential oils and their major components against Culex quinquefasciatus Say (Diptera: Culicidae): Synergism–antagonism effects. Insects 2018, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Conti, B.; Leonardi, M.; Pistelli, L.; Profeti, R.; Ouerghemmi, I.; Benelli, G. Larvicidal and repellent activity of essential oils from wild and cultivated Ruta chalepensis L. (Rutaceae) against Aedes albopictus Skuse (Diptera: Culicidae), an arbovirus vector. Parasitol. Res. 2013, 112, 991–999. [Google Scholar] [CrossRef]
- Guidobaldi, F.; May-Concha, I.; Guerenstein, P. Morphology and physiology of the olfactory system of blood-feeding insects. J. Physiol. Paris 2014, 108, 96–111. [Google Scholar] [CrossRef]
- Giatropoulos, A.; Kimbaris, A.; Michaelakis, A.; Papachristos, D.P.; Polissiou, M.G.; Emmanouel, N. Chemical composition and assessment of larvicidal and repellent capacity of 14 Lamiaceae essential oils against Aedes albopictus. Parasitol. Res. 2018, 117, 1953–1964. [Google Scholar] [CrossRef]
- Giatropoulos, A.; Pitarokili, D.; Papaioannou, F.; Papachristos, D.P.; Koliopoulos, G.; Emmanouel, N.; Tzakou, O.; Michaelakis, A. Essential oil composition, adult repellency and larvicidal activity of eight Cupressaceae species from Greece against Aedes albopictus (Diptera: Culicidae). Parasitol. Res. 2013, 112, 1113–1123. [Google Scholar] [CrossRef]
- Barnard, D.R.; Xue, R.-D. Laboratory evaluation of mosquito repellents against Aedes albopictus, Culex nigripalpus, and Ochlerotatus triseriatus (Diptera: Culicidae). J. Med. Entomol. 2004, 41, 726–730. [Google Scholar] [CrossRef] [Green Version]
- Bano, A.; Ahmad, M.; Hadda, T.B.; Saboor, A.; Sultana, S.; Zafar, M.; Khan, M.P.Z.; Arshad, M.; Ashraf, M.A. Quantitative ethnomedicinal study of plants used in the skardu valley at high altitude of Karakoram-Himalayan range, Pakistan. J. Ethnobiol. Ethnomed. 2014, 10, 43. [Google Scholar] [CrossRef] [Green Version]
- Hayat, M.Q.; Ashraf, M.; Khan, M.A.; Yasmin, G.; Shaheen, N.; Jabeen, S. Phylogenetic analysis of Artemisia L. (Asteraceae) based on micromorphological traits of pollen grains. Afr. J. Biotechnol. 2009, 8, 6561–6568. [Google Scholar]
- Wiwattanapatapee, R.; Sae-Yun, A.; Petcharat, J.; Ovatlarnporn, C.; Itharat, A. Development and evaluation of granule and emulsifiable concentrate formulations containing Derris elliptica extract for crop pest control. J. Agric. Food Chem. 2009, 57, 11234–11241. [Google Scholar] [CrossRef] [PubMed]
- WHO. Increasing the Relevance of Education for Health Professionals: Report of a WHO Study Group on Problem-Solving Education for the Health Professions [Meeting Held in Geneva from 20 to 23 October 1992]; World Health Organization: Geneva, Switzerland, 1993. [Google Scholar]
- WHO. Informal Consultation on Malaria Elimination: Setting up the WHO Agenda, Tunis, 25–26 February 2006; World Health Organization: Geneva, Switzerland, 2006. [Google Scholar]
- WHO Study Group. Malaria Vector Control and Personal Protection; WHO: Geneva, Switzerland, 2006; Volume 936. [Google Scholar]
- Ahmad, K.W.; Freed, S.; Shoukat, R.F. Efficacy of entomopathogenic fungi and botanicals on development of Musca domestica. J. Entomol. Zool. Stud. 2017, 5, 593–599. [Google Scholar]
- Trisyono, A.; Whalon, M.E. Toxicity of neem applied alone and in combinations with Bacillus thuringiensis to Colorado potato beetle (Coleoptera: Chrysomelidae). J. Econ. Entomol. 1999, 92, 1281–1288. [Google Scholar] [CrossRef]
- Hummelbrunner, L.A.; Isman, M.B. Acute, sublethal, antifeedant, and synergistic effects of monoterpenoid essential oil compounds on the tobacco cutworm, Spodoptera litura (Lep., Noctuidae). J. Agric. Food Chem. 2001, 49, 715–720. [Google Scholar] [CrossRef]
- WHO. Malaria Vector Control and Personal Protection: Report of a WHO Study Group; World Health Organization: Geneva, Switzerland, 2006. [Google Scholar]
- Keziah, E.A.; Nukenine, E.N.; Danga, S.P.Y.; Younoussa, L.; Esimone, C.O. Creams formulated with Ocimum gratissimum L. and Lantana camara L. crude extracts and fractions as mosquito repellents against Aedes aegypti L.(Diptera: Culicidae). J. Insect Sci. 2015, 15. [Google Scholar] [CrossRef] [Green Version]
- Schreck, C.; McGovern, T. Repellents and other personal protection strategies against Aedes albopictus. J. Am. Mosq. Control Assoc. 1989, 5, 247–250. [Google Scholar]
- Ai, C.; Norton, E.C. Interaction terms in logit and probit models. Econ. Lett. 2003, 80, 123–129. [Google Scholar] [CrossRef]
- Abbott, W.S. Abbott’s formula. J. Econ. Entomol. 1925, 18, 267–268. [Google Scholar]
- Cheng, S.-S.; Huang, C.-G.; Chen, Y.-J.; Yu, J.-J.; Chen, W.-J.; Chang, S.-T. Chemical compositions and larvicidal activities of leaf essential oils from two eucalyptus species. 2009, 100, 452–456. Bioresour Technol. 2009, 100, 452–456. [Google Scholar] [CrossRef]
- Cheng, S.-S.; Liu, J.-Y.; Huang, C.-G.; Hsui, Y.-R.; Chen, W.-J.; Chang, S.-T. Insecticidal activities of leaf essential oils from Cinnamomum osmophloeum against three mosquito species. Bioresour. Technol. 2009, 100, 457–464. [Google Scholar] [CrossRef]
- Alvarez Costa, A.; Naspi, C.V.; Lucia, A.; Masuh, H.M. Repellent and larvicidal activity of the essential oil from Eucalyptus nitens against Aedes aegypti and Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 2017, 54, 670–676. [Google Scholar] [CrossRef] [PubMed]
- Ahbirami, R.; Zuharah, W.F.; Yahaya, Z.S.; Dieng, H.; Thiagaletchumi, M.; Fadzly, N.; Ahmad, A.H.; Bakar, S.A. Oviposition deterring and oviciding potentials of Ipomoea cairica L. leaf extract against dengue vectors. Trop. Biomed. 2014, 31, 456–465. [Google Scholar] [PubMed]
- Ma, T.; Yan, H.; Shi, X.; Liu, B.; Ma, Z.; Zhang, X. Comprehensive evaluation of effective constituents in total alkaloids from Sophora alopecuroides L. and their joint action against aphids by laboratory toxicity and field efficacy. Ind. Crop. Prod. 2018, 111, 149–157. [Google Scholar] [CrossRef]
- Ma, X.M.; Liu, X.X.; Zhang, Q.W.; Zhao, J.Z.; Cai, Q.N.; Ma, Y.A.; Chen, D.M. Assessment of cotton aphids, Aphis gossypii, and their natural enemies on aphid-resistant and aphid-susceptible wheat varieties in a wheat–cotton relay intercropping system. Entomol. Exp. Appl. 2006, 121, 235–241. [Google Scholar] [CrossRef]
- Cai, Q.; Zhang, Q.; Cheo, M. Contribution of indole alkaloids to Sitobion avenae (F.) resistance in wheat. J. Appl. Entomol. 2004, 128, 517–521. [Google Scholar] [CrossRef]
- Cai, Q.-N.; Han, Y.; Cao, Y.-Z.; Hu, Y.; Zhao, X.; Bi, J.-L. Detoxification of gramine by the cereal aphid Sitobion avenae. J. Chem. Ecol. 2009, 35, 320–325. [Google Scholar] [CrossRef] [Green Version]
- Wanchun, L.; Yunshou, L.; Liyi, M.; Shin-Foon, C. Toxicity of cytisine against the mustard aphid Lipaphis erysimi Kaltenbach (Homoptera: Aphididae) and its effect on esterases. Pest. Biochem. Physiol. 1999, 65, 1–5. [Google Scholar] [CrossRef]
- Pavela, R. Acute toxicity and synergistic and antagonistic effects of the aromatic compounds of some essential oils against Culex quinquefasciatus Say larvae. Parasitol. Res. 2015, 114, 3835–3853. [Google Scholar] [CrossRef]
- Pavela, R. History, presence and perspective of using plant extracts as commercial botanical insecticides and farm products for protection against insects–A review. Plant Prot. Sci. 2016, 52, 229–241. [Google Scholar]
- Tak, J.H.; Jovel, E.; Isman, M.B. Comparative and synergistic activity of Rosmarinus officinalis L. essential oil constituents against the larvae and an ovarian cell line of the cabbage looper, Trichoplusia ni (Lepidoptera: Noctuidae). Pest Manag. Sci. 2016, 72, 474–480. [Google Scholar] [CrossRef] [PubMed]
- Paluch, G.; Grodnitzky, J.; Bartholomay, L.; Coats, J. Quantitative structure−activity relationship of botanical sesquiterpenes: Spatial and contact repellency to the yellow fever mosquito, Aedes aegypti. J. Agric. Food Chem. 2009, 57, 7618–7625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Domenech, R.; Garcia-Mujica, P.; Gil, U.; Casanova, C.; Mireilli Beltran, J.; Galvez, J. Search of QSAR models for natural sesquiterpenes repellent activity against the Yellow Fever mosquito, Aedes aegypti. Afinidad 2010, 67, 187–192. [Google Scholar]
- Rizvi, S.A.H.; Xie, F.; Ling, S.; Zeng, X. Development and evaluation of emulsifiable concentrate formulation containing Sophora alopecuroides L. extract for the novel management of Asian citrus psyllid. Environ. Sci. Pollut. Res. 2019, 26, 21871–21881. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | LC50 (ug/mL) | LC30 (ug/mL) | Slope ± SE | χ2 (d.f) | p-Value | |
|---|---|---|---|---|---|---|
| Sophora alopecuroides | Larvicidal activity | 4.81 (3.04–6.33) | 2.94 (2.47–3.90) | 1.99 ± 0.186 | 1.28 (4) | 0.002 |
| Ovicidal activity | 12.08 (10.40–14.28) | 4.22 (3.33–5.39) | 1.81 ± 0.182 | 0.38 (4) | 0.001 | |
| Activities | Chemicals | LC50 ug/mL | LC30 ug/mL | LC20 ug/mL | SLOP ± SE | χ2 (d.f) | p-Value |
|---|---|---|---|---|---|---|---|
| Larvicidal activity | Sophocarpin | 33.44 (29.99 ± 36.69) | 27.63 (24.20 ± 30.74) | 17.73 (14.51 ± 20.65) | 3.05 ± 0.28 | 18.33 (4) | 0.002 |
| Sophordine | 41.09 (37.43 ± 44.69) | 33.52 (29.92 ± 36.85) | 20.89 (17.44 ± 24.03) | 2.86 ± 0.27 | 14.15 (4) | 0.005 | |
| Ovicidal activity | Sophocarpin | 83.82 (76.44 ± 93.68) | 62.47 (57.62 ± 68.11) | 31.57 (27.39 ± 35.28) | 1.98 ± 0.18 | 4.98 (4) | 0.003 |
| Sophordine | 46.69 (43.52 ± 49.91) | 36.64 (33.62 ± 39.50) | 20.87 (18.01 ± 23.51) | 2.40 ± 0.18 | 14 (4) | 0.002 |
| Larvicidal Activity (%) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Pure Compound | Binary Mixture | |||||||
| Compound a | Compound b | Dose (ug/mL) | Observed a | Observed b | Expected | Observed | χ2 | Effect |
| Sophocarpin | Sophordine | LC30 + LC30 | 24.5 | 32.8 | 42.5 | 60.2 | 7.14 | Synergistic |
| Sophocarpin | Sophordine | LC20 + LC20 | 29.7 | 33.6 | 44.1 | 56.3 | 3.26 | Antagonistic |
| Ovicidal Activity(%) | ||||||||
| Sophocarpin | Sophordine | LC30 + LC30 | 36.5 | 33.4 | 44.5 | 43.1 | 0.047 | Antagonistic |
| Sophocarpin | Sophordine | LC20 + LC20 | 14.5 | 18.2 | 27.7 | 27 | 0.022 | Antagonistic |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shoukat, R.F.; Shakeel, M.; Rizvi, S.A.H.; Zafar, J.; Zhang, Y.; Freed, S.; Xu, X.; Jin, F. Larvicidal, Ovicidal, Synergistic, and Repellent Activities of Sophora alopecuroides and Its Dominant Constituents Against Aedes albopictus. Insects 2020, 11, 246. https://doi.org/10.3390/insects11040246
Shoukat RF, Shakeel M, Rizvi SAH, Zafar J, Zhang Y, Freed S, Xu X, Jin F. Larvicidal, Ovicidal, Synergistic, and Repellent Activities of Sophora alopecuroides and Its Dominant Constituents Against Aedes albopictus. Insects. 2020; 11(4):246. https://doi.org/10.3390/insects11040246
Chicago/Turabian StyleShoukat, Rana Fartab, Muhammad Shakeel, Syed Arif Hussain Rizvi, Junaid Zafar, Yuxin Zhang, Shoaib Freed, Xiaoxia Xu, and Fengliang Jin. 2020. "Larvicidal, Ovicidal, Synergistic, and Repellent Activities of Sophora alopecuroides and Its Dominant Constituents Against Aedes albopictus" Insects 11, no. 4: 246. https://doi.org/10.3390/insects11040246
APA StyleShoukat, R. F., Shakeel, M., Rizvi, S. A. H., Zafar, J., Zhang, Y., Freed, S., Xu, X., & Jin, F. (2020). Larvicidal, Ovicidal, Synergistic, and Repellent Activities of Sophora alopecuroides and Its Dominant Constituents Against Aedes albopictus. Insects, 11(4), 246. https://doi.org/10.3390/insects11040246