Her Majesty’s Desert Throne: The Ecology of Queen Butterfly Oviposition on Mojave Milkweed Host Plants


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Davidson, D.W.; Samson, D.A.; Inouye, R.S. Granivory on the Chihuahuan Desert: Interactions within and between trophic levels. Ecology 1985, 66, 486–502. [Google Scholar] [CrossRef]
- Polis, G.A. Complex trophic interactions in deserts: An empirical critique of food-web theory. Am. Nat. 1991, 138, 123–155. [Google Scholar] [CrossRef]
- Davidson, D.W.; Inouye, R.S.; Brown, J.H. Granivory in a desert ecosystem: Experimental evidence for indirect facilitation of ants by rodents. Ecology 1984, 65, 1780–1786. [Google Scholar] [CrossRef] [Green Version]
- Nevo, D. The desert locust, Schistocerca gregaria, and its control in the land of Israel and the near east in antiquity, with some reflections on its appearance in Israel in modern times. Phytoparasitica 1996, 24, 7–32. [Google Scholar] [CrossRef]
- Cane, J.H.; Minckley, R.L.; Kervin, L.J.; Roulston, T.H.; Williams, N.M. Complex responses within a desert bee guild (Hymenoptera: Apiformes) to urban habitat fragmentation. Ecol. Appl. 2006, 16, 632–644. [Google Scholar] [CrossRef] [Green Version]
- Aranda-Rickert, A.; Diez, P.; Marazzi, B. Extrafloral nectar fuels ant life in deserts. AoB Plants 2014, 6, plu068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emmel, T.C.; Emmel, J.F. Selection and host overlap in two desert Papilio butterflies. Ecology 1968, 50, 158–159. [Google Scholar] [CrossRef]
- Marques, E.S.D.A.; Price, P.W.; Cobb, N.S. Resource abundance and insect herbivore diversity on woody Fabaceous desert plants. Environ. Entomol. 2000, 29, 696–703. [Google Scholar] [CrossRef] [Green Version]
- Pfeiler, E.; Markow, T.A. Phylogeography of the cactophilic Drosophila and other arthropods associated with cactus necroses in the Sonoran Desert. Insects 2011, 2, 218–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gustafsson, K.M.; Agrawal, A.A.; Lewenstein, B.V.; Wolf, S.A. The monarch butterfly through space and time: The social construction of an icon. BioScience 2015, 65, 612–622. [Google Scholar] [CrossRef] [Green Version]
- Dilts, T.; Steele, M.; Engler, J.D.; Pelton, E.M.; Jepsen, S.J.; McKnight, S.J.; Taylor, A.R.; Fallon, C.E.; Black, S.H.; Cruz, E.E.; et al. Host plants and climate structure habitat associations of the western monarch butterfly. Front. Ecol. Evol. 2019, 7, 188. [Google Scholar] [CrossRef] [Green Version]
- Schultz, C.B.; Brown, L.M.; Pelyon, E.; Crone, E.E. Citizen science monitoring demonstrates dramatic declines of monarch butterflies in western North America. Biol. Conserv. 2017, 214, 343–346. [Google Scholar] [CrossRef]
- Emmel, T.C.; Emmel, J.F. Butterflies of Southern California; Natural History Museum of Los Angeles: Los Angeles, CA, USA, 1973; pp. 1–148. [Google Scholar]
- Agrawal, A.A.; Petschenka, G.; Bingham, R.A.; Weber, M.G.; Rasmann, S. Toxic cardenolides: Chemical ecology and coevolution of specialized plant–herbivore interactions. New Phytol. 2012, 194, 28–45. [Google Scholar] [CrossRef] [PubMed]
- Hughes, P.R.; Radke, C.D.; Renwick, J.A.A. A simple, low-input method for continuous laboratory rearing of the monarch butterfly (Lepidoptera: Danaidae) for research. Am. Entomol. 1993, 29, 109–112. [Google Scholar] [CrossRef]
- Robertson, G.F.; Zalucki, M.P.; Paine, T.D. Larval host choice of the monarch butterfly (Danaus plexippus L.) on four native California desert milkweed species. J. Insect Behav. 2015, 28, 582–592. [Google Scholar] [CrossRef]
- Grodsky, S.M.; Hernandez, R.R. Digital (natural) histories of species existing in the Internet ecosystem. BioScience. in press.
- Grodsky, S.M.; Saul-Gerschenz, L.S.; Moore-O’Leary, K.A.; Whitney, J.P.; Hernandez, R.R. Hare don’t care! Consumption of a rare, desert milkweed containing phytochemicals by the black-tailed jackrabbit. J. Arid. Environ. 2020, 174, 103991. [Google Scholar] [CrossRef]
- Moore, K.A.; André, J.M. Rare plant diversity in the California deserts: Priorities for research and conservation. Fremontia 2004, 42, 9–14. [Google Scholar]
- Agrawal, A.A.; Fishbein, M.; Jetter, R.; Salminen, J.; Goldstein, J.B.; Freitag, A.E.; Sparks, J.P. Phylogenetic ecology of leaf surface traits in the milkweeds (Asclepias spp.): Chemistry, ecophysiology, and insect behavior. New Phytol. 2009, 183, 848–867. [Google Scholar] [CrossRef]
- Panter, K.E.; Gardner, D.R.; Lee, S.T.; Pfister, J.A.; Ralphs, M.H.; Stegelmeier, B.L.; James, L.F. Important Poisonous Plants of the United States in Veterinary Toxicology: Basic and Clinical Principles; Gupta, R.C., Ed.; Academic Press: Waltham, MA, USA, 2007; pp. 825–872. [Google Scholar]
- Van Dyck, H.; Wiklund, C. Seasonal butterfly design: Morphological plasticity among three developmental pathways relative to sex, flight and thermoregulation. J. Evol. Biol. 2002, 15, 216–225. [Google Scholar] [CrossRef]
- Pitman, G.M.; Flockhart, D.T.T.; Norris, D.R. Patterns and causes of oviposition in monarch butterflies: Implications for milkweed restoration. Biol. Conserv. 2018, 217, 54–65. [Google Scholar] [CrossRef] [Green Version]
- Mayhew, P.J. Adaptive patterns of host-plant selection by phytophagous insects. Oikos 1997, 79, 417–428. [Google Scholar] [CrossRef]
- Zalucki, M.P.; Clarke, A.R.; Malcolm, S.B. Ecology and behavior of first instar larval Lepidoptera. Annu. Rev. Entomol. 2002, 47, 361–393. [Google Scholar] [CrossRef] [PubMed]
- Feeny, P. Plant Apparency and Chemical Defense in Biochemical Interaction between Plants and Insects; Wallace, J.W., Mansell, R.L., Eds.; Plenum Press: New York, NY, USA, 1976; pp. 1–40. [Google Scholar]
- House, P.K.; Buck, B.J.; Ramelli, A.R. Geological Assessment of Piedmont and Playa Flood Hazards in the Ivanpah Valley Area, Clark County, Nevada. 2010. Available online: https://pdfs.semanticscholar.org/8a9c/70c99093a3d043bacc9993c2447337398370.pdf (accessed on 15 February 2020).
- Coolidge, K.R. The life history of Danaus Berenice strigosa Bates (Lepidoptera: Danaidae). Trans. Am. Entomol. Soc. 1926, 51, 27–33. [Google Scholar]
- Hothorn, T.; Bretz, F.; Westfall, P.; Heiberger, R.M.; Schützenmeister, A. Package ‘Multcomp’. 2013. Available online: https://cran.rproject.org/web/packages/multcomp/multcomp.pdf (accessed on 15 February 2020).
- Hughes, J. Package ‘Reghelper’. 2020. Available online: https://cran.r-project.org/web/packages/reghelper/reghelper.pdf (accessed on 3 April 2020).
- Gigot, C. Package ‘Epiphy’. 2018. Available online: https://cran.r-project.org/web/packages/epiphy/epiphy.pdf (accessed on 15 February 2020).
- Sakai, W.H. Notes on Danaus gilippus strigosus (Nymphalidae: Danainae) in southern California. J. Lepid. Soc. 1993, 47, 160–161. [Google Scholar]
- Austin, G.T.; Leary, P.J. Larval hostplants of butterflies in Nevada. Holarct. Lepid. 2008, 12, 1–2. [Google Scholar]
- Saul-Gershenz, L.S.; Grodsky, S.M.; Hernandez, R.R. Ecology of the western queen butterfly Danaus gilippus thersippus (Lepidoptera:Nymphalidae) in the Mojave and Sonoran Deserts. Insects. under review.
- California Native Plant Society, Rare Plant Program. 2020. Inventory of Rare and Endangered Plants of California (Online Edition, v8-03 0.39). Available online: http://www.rareplants.cnps.org (accessed on 2 March 2020).
- Dingle, H.; Zalucki, M.P.; Rochester, W.A.; Armijo-Prewitt, T. Distribution of the monarch butterfly, Danaus plexippus (L.) (Lepidoptera: Nymphalidae), in western North America. Biol. J. Linn. Soc. 2005, 4, 491–500. [Google Scholar] [CrossRef] [Green Version]
- Brower, L.P. Experimental analyses of egg cannibalism in the monarch and queen butterflies, Danaus plexippus and D. gilippus berenice. Physiol. Biochem. Zool. 1961, 34, 287–296. [Google Scholar] [CrossRef]
- Brower, L.P. Evidence for interspecific competition in natural populations of the monarch and queen butterflies, Danaus plexippus and D. gilippus berenice in south central Florida. Ecology 1962, 43, 549–552. [Google Scholar] [CrossRef]
- Jones, P.L.; Agrawal, A.A. Beyond preference and performance: Host plant selection by monarch butterflies, Danaus plexippus. Oikos 2019, 128, 1092–1102. [Google Scholar] [CrossRef]
- Levins, R.; MacArthur, R. An hypothesis to explain the incidence of monophagy. Ecology 1969, 50, 910–911. [Google Scholar] [CrossRef]
- Cohen, J.A.; Brower, L.P. Oviposition and larval success of wild monarch butterflies (Lepidoptera: Danaidae) in relation to host plant size and cardenolide concentration. J. Kansas Entomol. Soc. 1982, 55, 343–348. [Google Scholar]
- Zalucki, M.P.; Kitching, R.L. Dynamics of oviposition in Danaus plexippus (Insecta: Lepidoptera) on milkweed, Asclepias spp. J. Zool. 1982, 198, 103–116. [Google Scholar] [CrossRef]
- Van Hook, T.; Zalucki, M.P. Oviposition by Danaus plexippus (Nymphalidae: Danainae) on Ascelpias viridis in northern Florida. J. Lepid. Soc. 1991, 45, 215–221. [Google Scholar]
- Oyeyele, S.O.; Zalucki, M.P. Cardiac glycosides and oviposition by Danaus plexippus on Asclepias fruticose in south-east Queensland (Australia), with notes of the effect of plant nitrogen content. Ecol. Entomol. 1990, 15, 177–185. [Google Scholar] [CrossRef]
- Moranz, R.; Brower, L.P. Geographic and temporal variation of cardenolide-based chemical defenses of queen butterfly (Danaus gilippus) in northern Florida. J. Chem. Ecol. 1998, 24, 905–932. [Google Scholar] [CrossRef]
- Bazzaz, F.A.; Chiariello, N.R.; Coley, P.D.; Pitelka, L.F. Allocating resources to reproduction and defense. BioScience 1987, 37, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Flockhart, D.T.T. Experimental examination of intraspecific density-dependent competition during the breeding period in monarch butterflies (Danaus plexippus). PLoS ONE 2012, 7, e45080. [Google Scholar] [CrossRef] [Green Version]
- Nail, K.R.; Stenoien, C.; Oberhauser, K.S. Immature monarch survival: Effects of site characteristics, density and time. Ann. Entomol. Soc. Am. 2015, 108, 680–690. [Google Scholar] [CrossRef]
- Dussourd, D.E.; Harvis, C.A.; Meinwald, J.; Eisner, T. Paternal allocation of sequestered plant pyrrolizidine alkaloid to eggs in the danaine butterfly, Danaus gilippus. Experientia 1989, 45, 896–898. [Google Scholar] [CrossRef]
- Grodsky, S.M.; Moore-O’Leary, K.A.; Hernandez, R.R. From butterflies to bighorns: Multi-dimensional species-species and species-process interactions may inform sustainable solar energy development in desert ecosystems. In Proceedings of the 31st Annual Desert Symposium, Zzyzx, CA, USA, 15–17 April 2017; Reynolds, R.L., Ed.; California State University Desert Studies Center: Zzyzx, CA, USA, 2017; pp. 322–327. [Google Scholar]
- Kremen, C.; Colwell, R.K.; Erwin, T.L.; Murphy, D.D.; Noss, E.F.; Sanijayan, M.A. Terrestrial arthropod assemblages: Their use in conservation planning. Conserv. Biol. 1993, 7, 79–808. [Google Scholar] [CrossRef] [Green Version]
- Grodsky, S.M.; Iglay, R.B.; Sorenson, C.E.; Moorman, C.E. Should invertebrates receive greater inclusion in wildlife research journals? J. Wildl. Manag. 2015, 79, 529–536. [Google Scholar] [CrossRef]
- Kuletz, V.L. The Tainted Desert: Environmental and Social Ruin in the American West; Routledge: New York, NY, USA, 1998; pp. 3–15. [Google Scholar]
- Hernandez, R.R.; Hoffacker, M.K.; Murphy-Mariscal, M.L.; Wu, G.C.; Allen, M.F. Solar energy development impacts on land cover change and protected areas. Proc. Natl. Acad. Sci. USA 2015, 112, 13579–13584. [Google Scholar] [CrossRef] [Green Version]
- Moore-O’Leary, K.A.; Hernandez, R.R.; Johnson, D.S.; Abella, S.R.; Tanner, K.E.; Swanson, A.C.; Kreitler, J.; Lovich, J.E. Sustainability of utility-scale solar energy—Critical ecological concepts. Front. Ecol. Environ. 2017, 15, 385–394. [Google Scholar] [CrossRef]
- Henry, E.; Brammer-Robbins, E.; Aschehoug, E.; Haddad, N. Do substitute species help or hinder enagered species management? Biol. Conserv. 2019, 232, 127–130. [Google Scholar] [CrossRef]







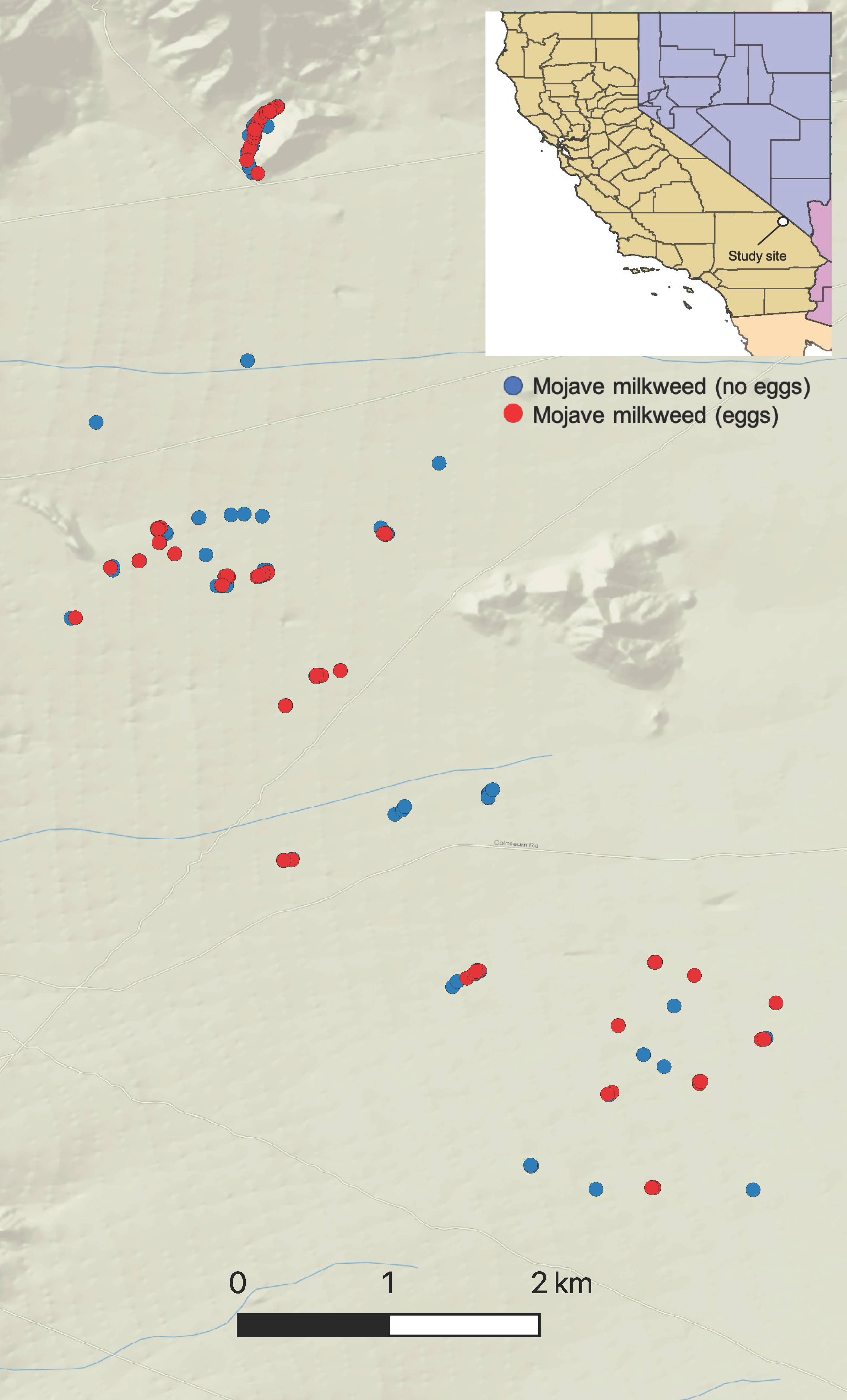{kind=link}
{kind=link}
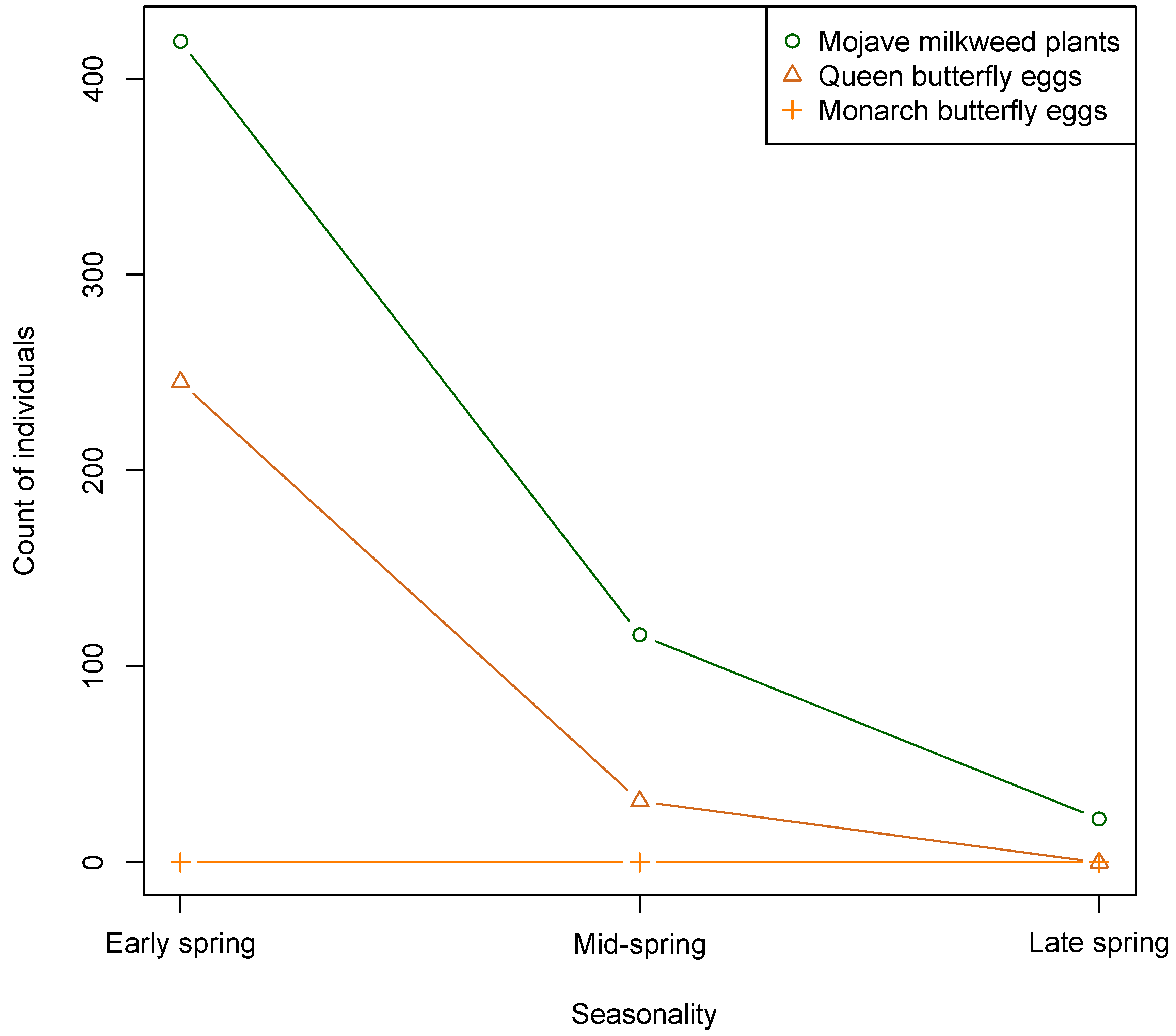{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Covariate | Standard Error | z Value | p Value | |
|---|---|---|---|---|
| Seasonality1 | ||||
| Early spring | 0.51 | 0.10 | 4.42 | <0.001 |
| Mid-spring | −0.48 | 0.11 | −4.46 | <0.001 |
| Life stage | ||||
| Seedling | −1.00 | 0.18 | −6.11 | <0.001 |
| Juvenile | −0.54 | 0.09 | −5.83 | <0.001 |
| Adult | 0.31 | 0.05 | 5.94 | <0.001 |
| Plant Traits | ||||
| Canopy cover | 0.37 | 0.09 | 4.03 | <0.001 |
| Height | −0.18 | 0.15 | −1.19 | 0.23 |
| Volume | 0.11 | 0.07 | 1.63 | 0.10 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grodsky, S.M.; Saul-Gershenz, L.S.; Moore-O’Leary, K.A.; Hernandez, R.R. Her Majesty’s Desert Throne: The Ecology of Queen Butterfly Oviposition on Mojave Milkweed Host Plants. Insects 2020, 11, 257. https://doi.org/10.3390/insects11040257
Grodsky SM, Saul-Gershenz LS, Moore-O’Leary KA, Hernandez RR. Her Majesty’s Desert Throne: The Ecology of Queen Butterfly Oviposition on Mojave Milkweed Host Plants. Insects. 2020; 11(4):257. https://doi.org/10.3390/insects11040257
Chicago/Turabian StyleGrodsky, Steven M., Leslie S. Saul-Gershenz, Kara A. Moore-O’Leary, and Rebecca R. Hernandez. 2020. "Her Majesty’s Desert Throne: The Ecology of Queen Butterfly Oviposition on Mojave Milkweed Host Plants" Insects 11, no. 4: 257. https://doi.org/10.3390/insects11040257
APA StyleGrodsky, S. M., Saul-Gershenz, L. S., Moore-O’Leary, K. A., & Hernandez, R. R. (2020). Her Majesty’s Desert Throne: The Ecology of Queen Butterfly Oviposition on Mojave Milkweed Host Plants. Insects, 11(4), 257. https://doi.org/10.3390/insects11040257