Are the Yellow and Red Marked Club-Tail Losaria coon the Same Species?



Abstract
:1. Introduction
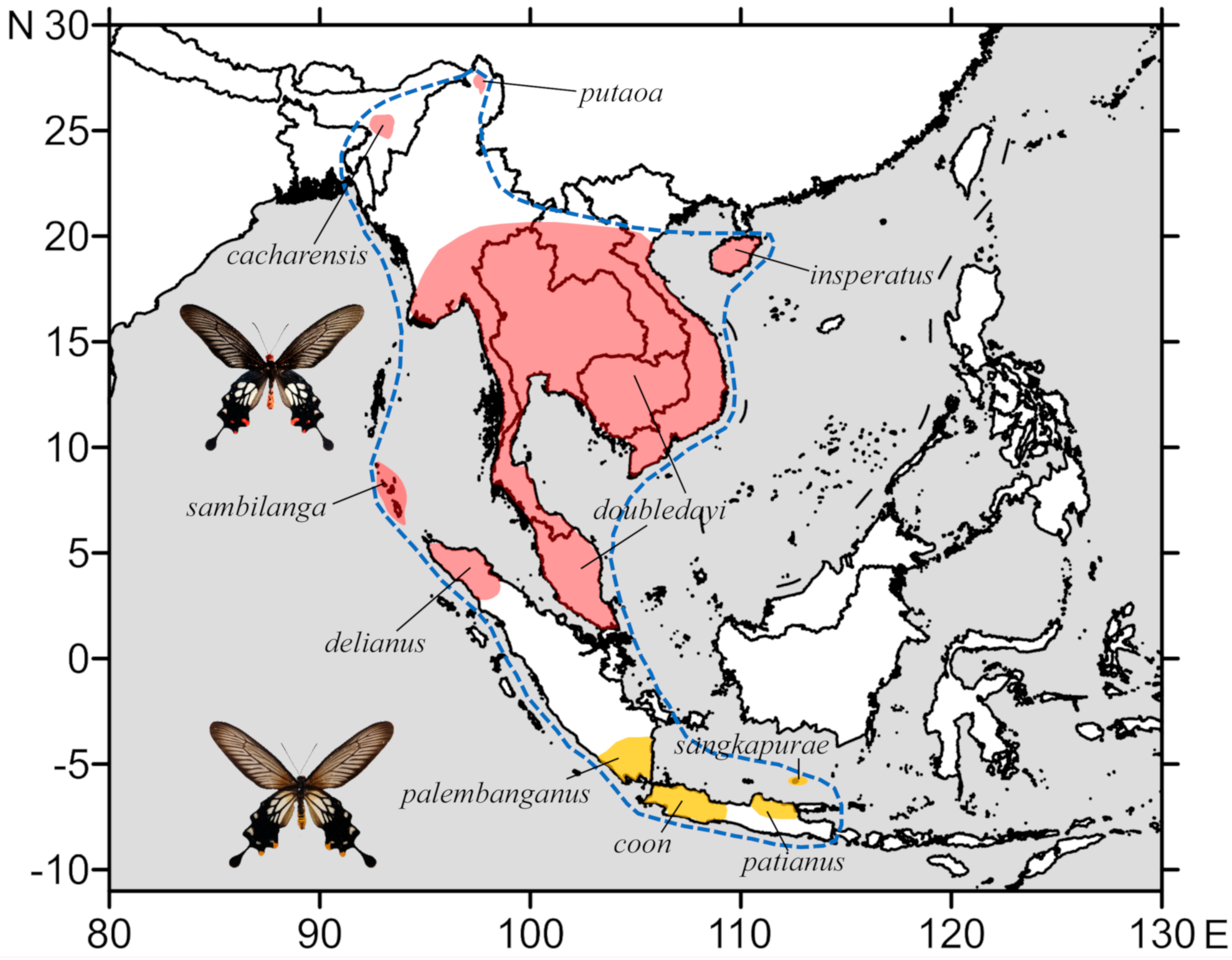
2. Materials and Methods
2.1. Taxon Sampling
2.2. DNA Extraction and Amplification
2.3. Phylogenetic Analyses
2.4. Molecular Species Delimitation
2.5. Morphological Comparison
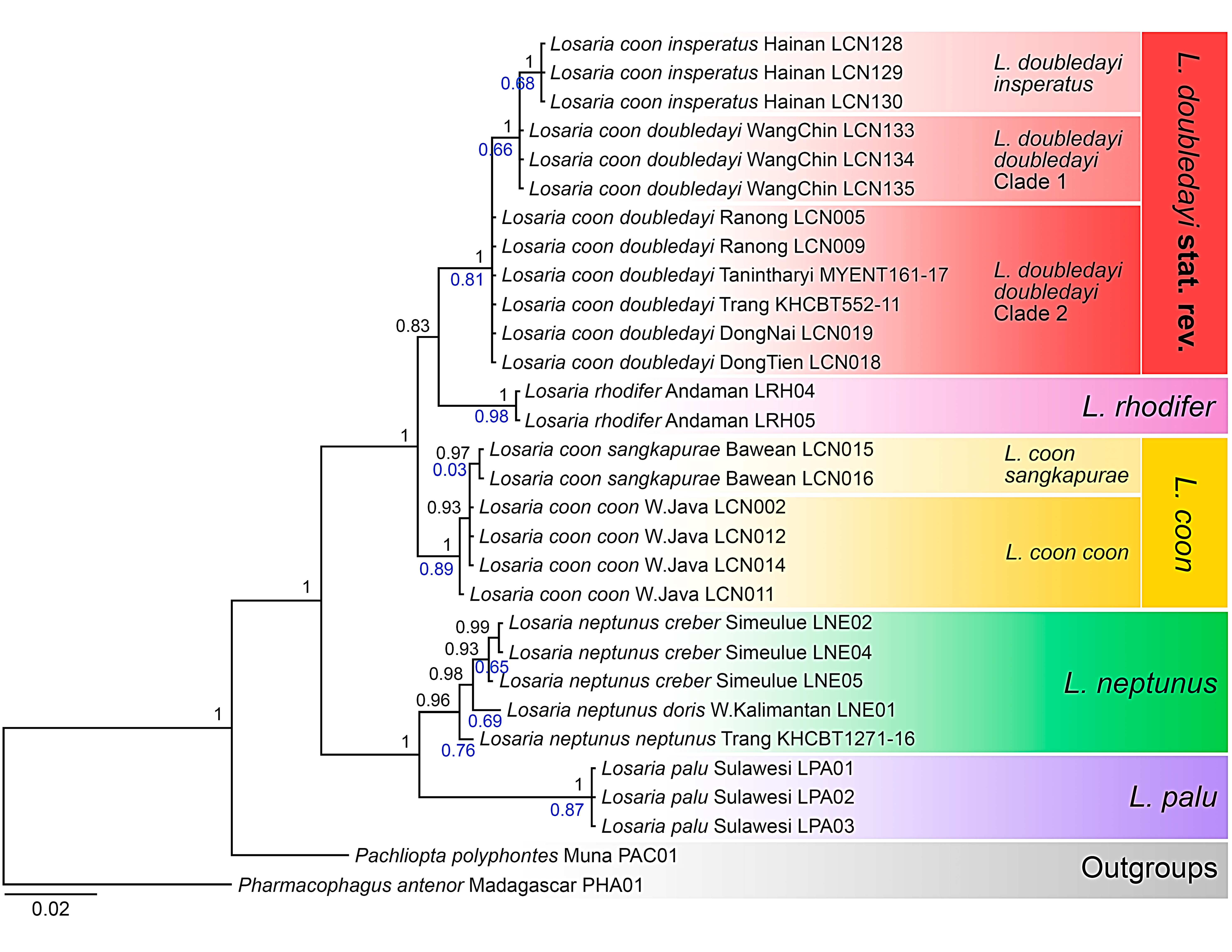
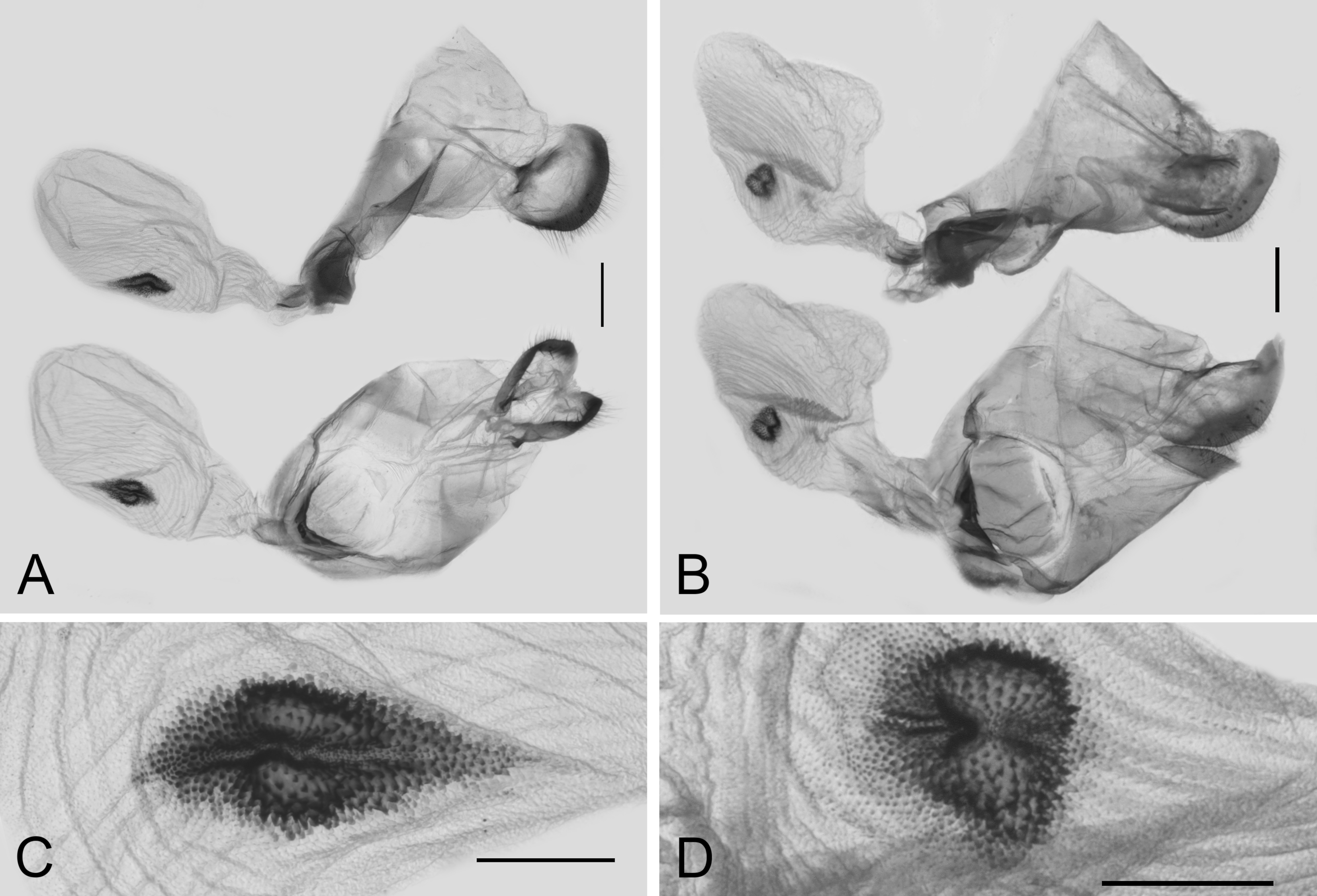
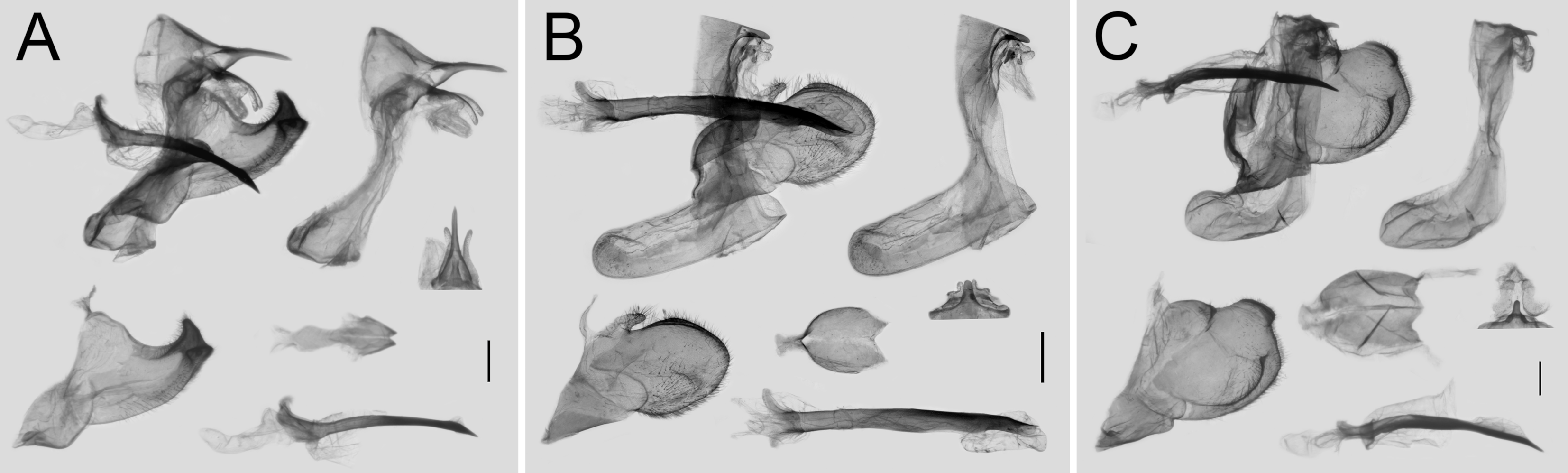
3. Results
3.1. Molecular Phylogenetic Relationships
3.2. Molecular Species Delimitation
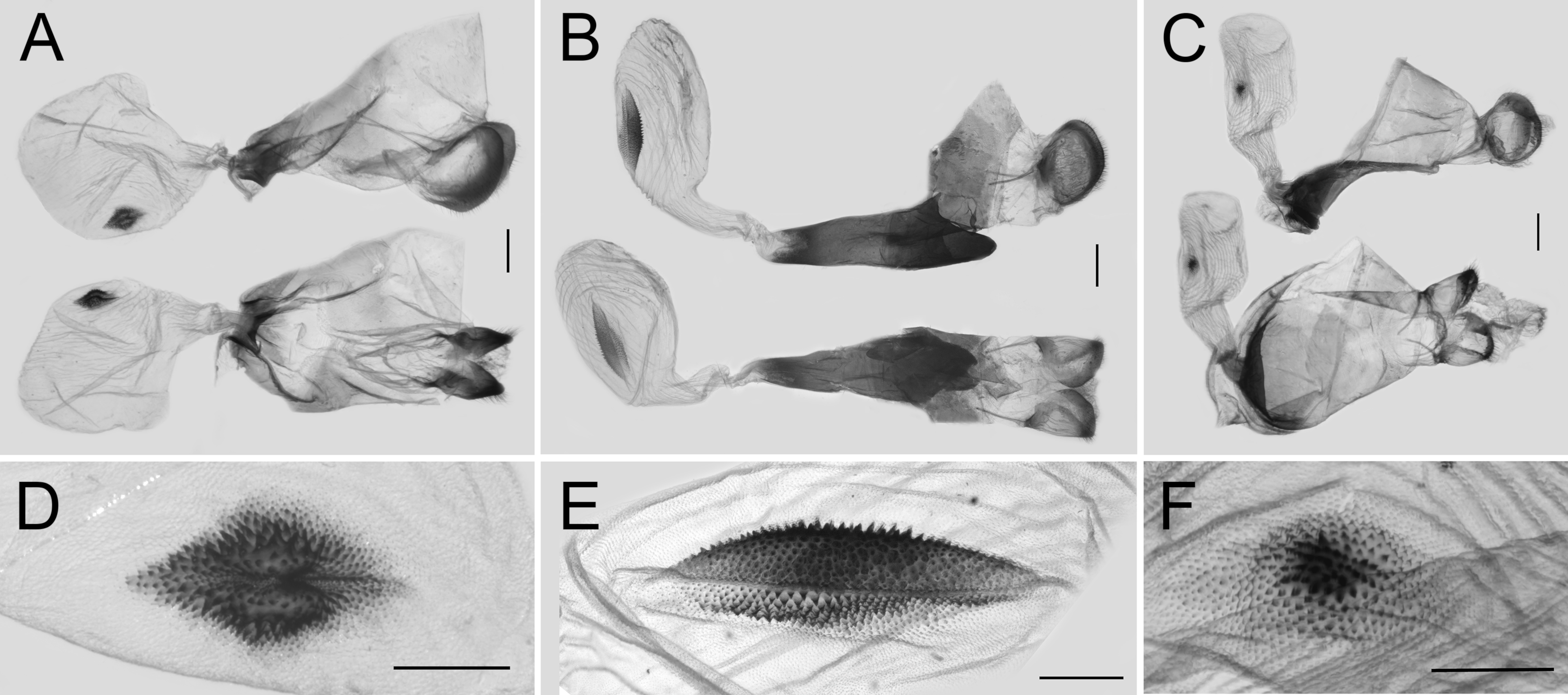
3.3. Morphological Differences
3.4. Updated Subspecies Checklist of Losaria (Moore, 1902)
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. List of Losaria Specimens Examined, Different Labels in Museum Specimens Are Separated with a ‘/’
References
- Hancock, D.L. Phylogeny of the Troidine butterflies (Lepidoptera: Papilionidae) revisited: Are the red-bodied swallowtails monophyletic. Aust. Entomol. 2007, 34, 33–42. [Google Scholar]
- Tsukada, E.; Nishiyama, Y. Butterflies of the South East Asian Islands. Part I. Papilionidae; Plapac Co., Ltd.: Tokyo, Japan, 1980. [Google Scholar]
- Wallace, A.R. On the phenomena of variation and geographical distribution as illustrated by the Papilionidae of the Malayan Region. Trans. Linn. Soc. Lond. 1865, 25, 1–71, pl. 71–78. [Google Scholar] [CrossRef]
- Fabricius, J.C. Entomologia Systematica: Emendata et Aucta. Tom. III. Pars I; Impensis C. G. Proft, Fil. et Soc.: Hafniae, Denmark, 1793. [Google Scholar]
- Rothschild, W. A Revision of the Papilios of the Eastern Hemisphere, Exclusive of Africa. Novit. Zool. 1895, 2, 167–463. [Google Scholar]
- Moore, F. Lepidoptera Indica Vol. V. Rhopalocera. Family Nymphalidæ. Family Riodinidæ. Family Papilionidæ; Lovell Reeve: London, UK, 1902. [Google Scholar]
- Jordan, H.E.K. Papilio cont’d, Eurycus, Leptocircus, Teinopalpus & Armandia. In Seitz, Macrolepidoptera of the World 9, Fauna Indoaustralica; Fritz Lehmann Verlag: Stuttgart, Germany, 1909. [Google Scholar]
- Evans, W.H. The Identification of Indian Butterflies, 2nd ed. Revised; Bombay Natural History Society: Madras, India, 1932. [Google Scholar]
- Talbot, G. The Fauna of British India, Including Ceylon and Burma. Butterflies. Vol. I. (Papilionidae & Pieridae); Taylor & Francis: London, UK, 1939. [Google Scholar]
- Ford, E.B. Studies on the chemistry of pigments in the Lepidoptera, with reference to their bearing on systematics. 4. The classification of of the Papilionidae. Trans. R. Entomol. Soc. 1944, 94, 201–223. [Google Scholar] [CrossRef]
- Condamine, F.L.; Sperling, F.A.H.; Wahlberg, N.; Rasplus, J.-Y.; Kergoat, G.J. What causes latitudinal gradients in species diversity? Evolutionary processes and ecological constraints on swallowtail biodiversity. Ecol. Lett. 2012, 15, 267–277. [Google Scholar] [CrossRef]
- Fernando, E.; Jangid, A.K.; Chowdhury, S.; Kehimar, I.; Lo, P.; Moonen, J. Common Clubtail. IUCN Red List Threat. Species 2019. [Google Scholar] [CrossRef]
- Bollino, M.; Sala, G. A new species of Atrophaneura coon from Bawean Island (Indonesia) (Lepidoptera: Papilionidae). Trop. Lepid. 1992, 3, 119–122. [Google Scholar]
- Tytler, H.C. Notes on some new and interesting butterflies chiefly from Burma. Part I. J. Bombay Nat. Hist. Soc. 1939, 41, 235–252. [Google Scholar]
- Hu, S.J.; Zhang, X.; Cotton, A.M.; Ye, H. Discovery of a third species of Lamproptera Gray, 1832 (Lepidoptera: Papilionidae). Zootaxa 2014, 3786, 469–482. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.J.; Cotton, A.M.; Condamine, F.L.; Duan, K.; Wang, R.J.; Hsu, Y.F.; Zhang, X.; Cao, J. Revision of Pazala Moore, 1888: The Graphium (Pazala) mandarinus (Oberthür, 1879) group, with treatments of known taxa and descriptions of new species and new subspecies (Lepidoptera: Papilionidae). Zootaxa 2018, 4441, 401–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, S.J.; Condamine, F.L.; Monastyrskii, A.L.; Cotton, A.M. A new species of the Graphium (Pazala) mandarinus group from Central Vietnam (Lepidoptera: Papilionidae). Zootaxa 2019, 4554, 286–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.H.; Cotton, A.M.; Condamine, F.L.; Wang, R.J.; Hsu, Y.F.; Duan, K.; Zhang, X.; Hu, S.J. Revision of Pazala Moore, 1888: The Graphium (Pazala) alebion and G. (P.) tamerlanus groups, with notes on taxonomic and distribution confusions (Lepidoptera: Papilionidae). Zootaxa 2020, 4759, 77–97. [Google Scholar] [CrossRef]
- Ylla, J.; Peigler, R.S.; Kawahara, A.Y. Cladistic analysis of moon moths using morphology, molecules, and behaviour: Actias Leach, 1815; Argema Wallengren, 1858; Graellsia Grote, 1896 (Lepidoptera: Saturniidae). Shilap Rev. Lepidopterol. 2005, 33, 299–317. [Google Scholar]
- Ivshin, N.; Krutov, V.; Romanov, D. Three new taxa of the genus Cechetra Zolotuhin & Ryabov, 2012 (Lepidoptera, Sphingidae) from South-East Asia with notes on other species of the genus. Zootaxa 2018, 4450, 1–25. [Google Scholar] [PubMed]
- Zolotuhin, V.V. Laothoe Fabricius, 1807 (Lepidoptera: Sphingidae): How many species? Eversmannia 2018, 54, 3–12. [Google Scholar]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; de Waard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.J.; Ning, T.; Fu, D.Y.; Haack, R.A.; Zhang, Z.; Chen, D.D.; Ma, X.Y.; Ye, H. Dispersal of the Japanese pine sawyer, Monochamus alternatus (Coleoptera: Cerambycidae), in mainland China as inferred from molecular data and associations to indices of human activity. PLoS ONE 2013, 8, e57568. [Google Scholar] [CrossRef] [Green Version]
- Folmer, O.; Black, M.B.; Hoch, W.; Lutz, R.A.; Vrijehock, R.C. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucl. Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, H.; Buhay, J.E.; Whiting, M.F.; Crandall, K.A. Many species in one: DNA barcoding overestimates the number of species when nuclear mitochondrial pseudogenes are coamplified. Proc. Natl. Acad. Sci. USA 2008, 105, 13486–13491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertheau, C.; Schuler, H.; Krumböck, S.; Arthofer, W.; Stauffer, C. Hit or miss in phylogenetic analyses: The case of the cryptic NUMTs. Mol. Ecol. Resour. 2011, 11, 1056–1059. [Google Scholar] [CrossRef]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2017, 34, 772–773. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, J.P.; Larget, B.; Alfaro, M.E. Bayesian phylogenetic model selection using reversible jump Markov Chain Monte Carlo. Mol. Biol. Evol. 2004, 21, 1123–1133. [Google Scholar] [CrossRef]
- Xie, W.G.; Lewis, P.O.; Fan, Y.; Kuo, L.; Chen, M.H. Improving marginal likelihood estimation for Bayesian phylogenetic model selection. Syst. Biol. 2011, 60, 150–160. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [Green Version]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef]
- Mutanen, M.; Kivelä, S.M.; Vos, R.A.; Doorenweerd, C.; Ratnasingham, S.; Hausmann, A.; Huemer, P.; Dincă, V.; van Nieukerken, E.J.; Lopez-Vaamonde, C.; et al. Species-level para- and polyphyly in DNA barcode gene trees: Strong operational bias in European Lepidoptera. Syst. Biol. 2016, 65, 1024–1040. [Google Scholar] [CrossRef]
- Meyer, C.P.; Paulay, G. DNA barcoding: Error rates based on comprehensive sampling. PLoS Biol. 2005, 3, e422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moonen, J.J.M. Notes on some Papilionidae (Lepidoptera) from Indonesia. Trans. Lepidopterol. Soc. Jpn. 1998, 49, 219–228. [Google Scholar]
- Condamine, F.L.; Toussaint, E.F.A.; Cotton, A.M.; Genson, G.S.; Sperling, F.A.H.; Kergoat, G.J. Fine-scale biogeographical and temporal diversification processes of peacock swallowtails (Papilio subgenus Achillides) in the Indo-Australian Archipelago. Cladistics 2013, 29, 88–111. [Google Scholar] [CrossRef]
- Condamine, F.L.; Toussaint, E.F.A.; Clamens, A.-L.; Genson, G.S.; Sperling, F.A.H.; Kergoat, G.J. Deciphering the evolution of birdwing butterflies 150 years after Alfred Russel Wallace. Sci. Rep. 2015, 5, 11860. [Google Scholar] [CrossRef] [Green Version]
- Beck, J.; Kitching, I.J.; Linsenmair, K.E. Wallace’s line revisited: Has vicariance or dispersal shaped the distribution of Malesian hawkmoths (Lepidoptera: Sphingidae)? Biol. J. Linn. Soc. 2006, 89, 455–468. [Google Scholar] [CrossRef] [Green Version]
- Voris, H.K. Maps of Pleistocene sea levels in Southeast Asia: Shorelines, river systems and time durations. J. Biogeogr. 2000, 27, 1153–1167. [Google Scholar] [CrossRef] [Green Version]
- Miyazaki, S.; Saito, T.; Saito, K. Notes on the butterflies of the southern part of Vietnam (1). Yadoriga 2006, 208, 2–19. [Google Scholar]
- Kunte, K.; Sondhi, S.; Roy, P. Butterflies of India, v. 2.53. Available online: http://www.ifoundbutterflies.org/ (accessed on 1 May 2020).
- Igarashi, S.; Fukuda, K. The Life Histories of Asian Butterflies; Tokai University Press: Tokyo, Japan, 1997; Volume 1. [Google Scholar]
- Yao, T.L. Nine new species of Thottea (Aristolochiaceae) in Peninsular Malaysia and Singapore, with two taxa in Peninsular Malaysia redefined and a taxon lectotypified. Blumea 2013, 58, 245–262. [Google Scholar] [CrossRef] [Green Version]
- Robi, A.J.; Jose, P.A.; Udayan, P.S. Thottea sasidharaniana sp. nov (Aristolochiaceae) from Peninsular India. Nord. J. Bot. 2014, 32, 11–14. [Google Scholar] [CrossRef]
- Pao, N.T.; Upadhaya, K. Effect of fragmentation and anthropogenic disturbances on floristic composition and structure of subtropical broad leaved humid forest in Meghalaya, Northeast India. Appl. Ecol. Environ. Res. 2017, 15, 385–407. [Google Scholar] [CrossRef]
- Ansari, F.; Jeong, Y.; Putri, I.A.; Kim, S. Sociopsychological aspects of butterfly souvenir purchasing behavior at Bantimurung Bulusaraung National Park in Indonesia. Sustainability 2019, 11, 1789. [Google Scholar] [CrossRef] [Green Version]
- Hansen, M.C.; Wang, L.; Song, X.P.; Tyukavina, A.; Turubanova, S.; Potapov, P.V.; Stehman, S.V. The fate of tropical forest fragments. Sci. Adv. 2020, 6, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMorrow, J.; Talip, M.A. Decline of forest area in Sabah, Malaysia: Relationship to state policies, land code and land capability. Glob. Environ. Chang.-Hum. Policy Dimens. 2001, 11, 217–230. [Google Scholar] [CrossRef]
- Latinne, A.; Saputro, S.; Kalengkongan, J.; Kowel, C.L.; Gaghiwu, L.; Ransaleleh, T.A.; Nangoy, M.J.; Wahyuni, I.; Kusumaningrum, T.; Safari, D.; et al. Characterizing and quantifying the wildlife trade network in Sulawesi, Indonesia. Glob. Ecol. Conserv. 2020, 21, e00887. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon (Sample Code) | Locality | Collecting Date | Accession No. |
|---|---|---|---|
| Losaria coon coon (LCN002) | West Java, Indonesia | 2018-I | MT417883 |
| Losaria coon coon (LCN011) | West Java, Indonesia | 2018-I | MT417884 |
| Losaria coon coon (LCN012) | West Java, Indonesia | 2018-I | MT417883 |
| Losaria coon coon (LCN014) | West Java, Indonesia | 2018-I | MT417883 |
| Losaria coon sangkapurae (LCN015) | Bawean Is., Indonesia | 2018-II | MT417885 |
| Losaria coon sangkapurae (LCN016) | Bawean Is., Indonesia | 2018-II | MT417885 |
| Losaria coon doubledayi (MYENT161-17) | Lenya, Tanintharyi, Myanmar | 2015-V-29 | MYENT161-17 |
| Losaria coon doubledayi (KHCBT552-11) | Khao Chong, Trang, Thailand | 2010-XII-26 | KHCBT552-11 |
| Losaria coon doubledayi (LCN005) | Ranong, Thailand | 2018-II | MT417886 |
| Losaria coon doubledayi (LCN009) | Ranong, Thailand | 2018-II | MT417886 |
| Losaria coon doubledayi (LCN018) | Dong Tien, Binh Thuan, Vietnam | 2018-VII | MT417886 |
| Losaria coon doubledayi (LCN019) | Thac Mai, Dong Nai, Vietnam | 2018-VIII | MT417886 |
| Losaria coon doubledayi (LCN133) | Wang Chin, Phrae, Thailand | 2017-XI-11 | MT417888 |
| Losaria coon doubledayi (LCN134) | Wang Chin, Phrae, Thailand | 2017-XI-11 | MT417888 |
| Losaria coon doubledayi (LCN135) | Wang Chin, Phrae, Thailand | 2017-XI-11 | MT417888 |
| Losaria coon insperatus (LCN128) | Sanya, Hainan Is., China | 2018-V | MT417887 |
| Losaria coon insperatus (LCN129) | Sanya, Hainan Is., China | 2018-V | MT417887 |
| Losaria coon insperatus (LCN130) | Sanya, Hainan Is., China | 2018-V | MT417887 |
| Losaria rhodifer (LRH04) | Andaman Is., India | 2014-X | MT417889 |
| Losaria rhodifer (LRH05) | Andaman Is., India | 1962-III | MT417890 |
| Losaria neptunus neptunus (KHCBT1271-16) | Khao Chong, Trang, Thailand | 2014-V-2 | KHCBT1271-16 |
| Losaria neptunus doris (LNE01) | West Kalimantan, Indonesia | 2018-I | MT417891 |
| Losaria neptunus creber (LNE02) | Simeulue Is., Indonesia | 2010-VIII | MT417892 |
| Losaria neptunus creber (LNE04) | Simeulue Is., Indonesia | 2010-VIII | MT417893 |
| Losaria neptunus creber (LNE05) | Simeulue Is., Indonesia | 2010-VIII | MT417894 |
| Losaria palu (LPA01) | Donggala, Sulawesi, Indonesia | 2017-X | MT417895 |
| Losaria palu (LPA02) | Donggala, Sulawesi, Indonesia | 2018-II | MT417895 |
| Losaria palu (LPA03) | Donggala, Sulawesi, Indonesia | 2018-II | MT417895 |
| Pachliopta polyphontes (PAC01) | Muna Is., Indonesia | 2017-XII | MT417896 |
| Pharmacophagus antenor (PHA01) | Madagascar | 2016-VI | MT417897 |
| Taxon | Assessment | Tanglees |
|---|---|---|
| 1. L. coon | monophyletic | — |
| 1a. L. coon coon | paraphyletic | L. coon sangkapurae |
| 1b. L. coon sangkapurae | monophyletic | — |
| 2. L. doubledayi | monophyletic | — |
| 2a. L. doubledayi doubledayi 1 | paraphyletic | L. doubledayi doubledayi 2;L. doubledayi insperatus |
| 2b. L. doubledayi doubledayi 2 | paraphyletic | L. doubledayi insperatus |
| 2c. L. doubledayi insperatus | monophyletic | — |
| 3. L. rhodifer | monophyletic | — |
| 4. L. neptunus | monophyletic | — |
| 5. L. palu | monophyletic |
| 1a | 1b | 2a | 2b | 2c | 3 | 4 | 5 | |
|---|---|---|---|---|---|---|---|---|
| 1a. L. coon coon | ||||||||
| 1b. L. coon sangkapurae | 0.19 | |||||||
| 2a. L. doubledayi doubledayi 1 | 3.08 | 3.28 | ||||||
| 2b. L. doubledayi doubledayi 2 | 2.44 | 2.64 | 0.61 | |||||
| 2c. L. doubledayi insperatus | 3.57 | 3.77 | 0.46 | 1.08 | ||||
| 3. L. rhodifer | 3.17 | 3.35 | 3.35 | 3.00 | 3.35 | |||
| 4. L. neptunus | 6.11 | 6.32 | 6.76 | 6.32 | 7.03 | 6.47 | ||
| 5. L. palu | 7.83 | 8.04 | 8.92 | 8.40 | 9.28 | 9.61 | 5.19 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Z.-B.; Wang, Y.-Y.; Condamine, F.L.; Cotton, A.M.; Hu, S.-J. Are the Yellow and Red Marked Club-Tail Losaria coon the Same Species? Insects 2020, 11, 392. https://doi.org/10.3390/insects11060392
Xu Z-B, Wang Y-Y, Condamine FL, Cotton AM, Hu S-J. Are the Yellow and Red Marked Club-Tail Losaria coon the Same Species? Insects. 2020; 11(6):392. https://doi.org/10.3390/insects11060392
Chicago/Turabian StyleXu, Zhen-Bang, Yun-Yu Wang, Fabien L. Condamine, Adam M. Cotton, and Shao-Ji Hu. 2020. "Are the Yellow and Red Marked Club-Tail Losaria coon the Same Species?" Insects 11, no. 6: 392. https://doi.org/10.3390/insects11060392
APA StyleXu, Z. -B., Wang, Y. -Y., Condamine, F. L., Cotton, A. M., & Hu, S. -J. (2020). Are the Yellow and Red Marked Club-Tail Losaria coon the Same Species? Insects, 11(6), 392. https://doi.org/10.3390/insects11060392