Thermal Tolerance of Fruit-Feeding Butterflies (Lepidoptera: Nymphalidae) in Contrasting Mountaintop Environments


Abstract
:1. Introduction
2. Materials and Methods
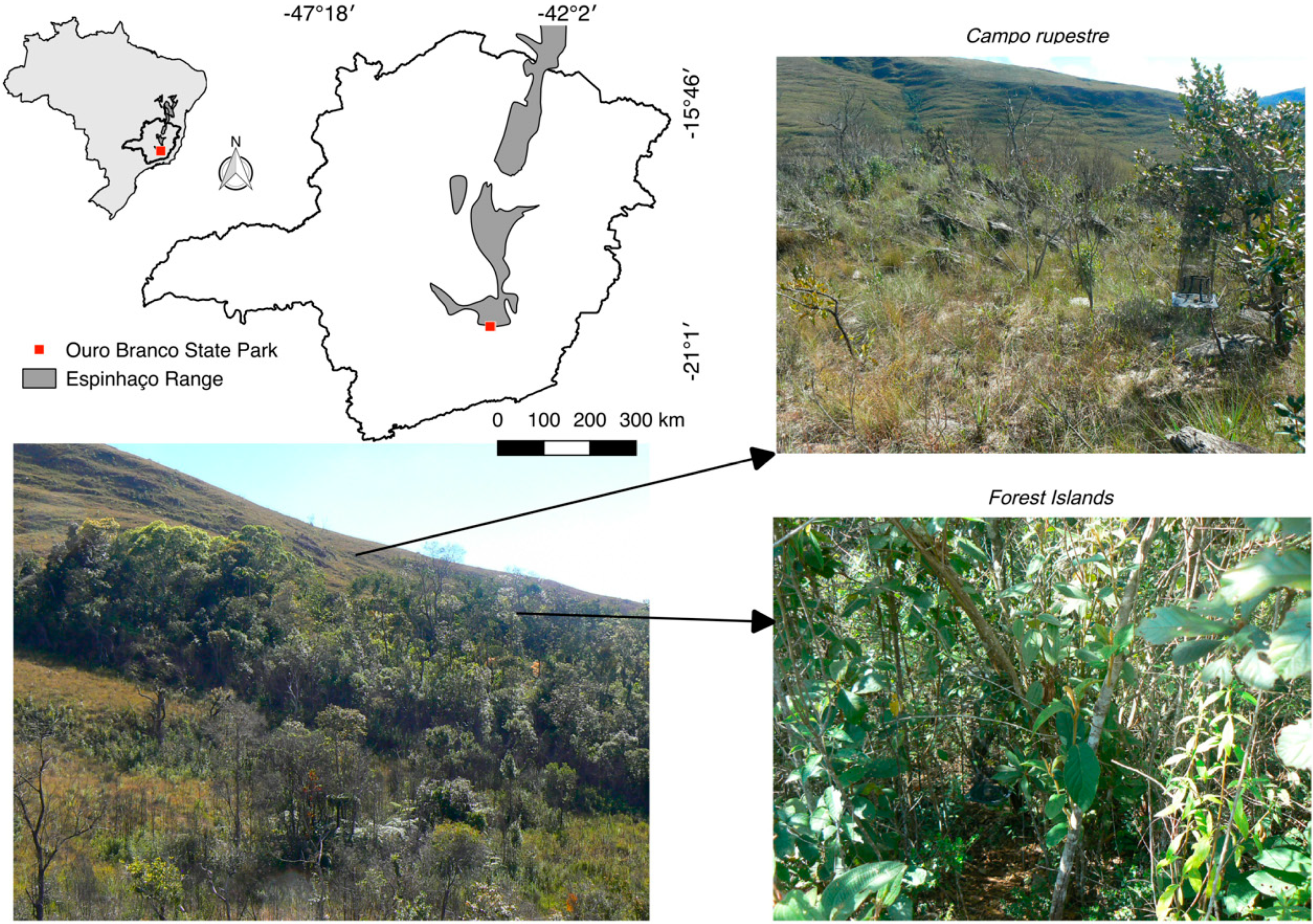
2.1. Study Area
2.2. Sampling Design
2.3. Thermal Tolerance Test
2.4. Statistical Analysis
2.5. Phylogenetic Correlation
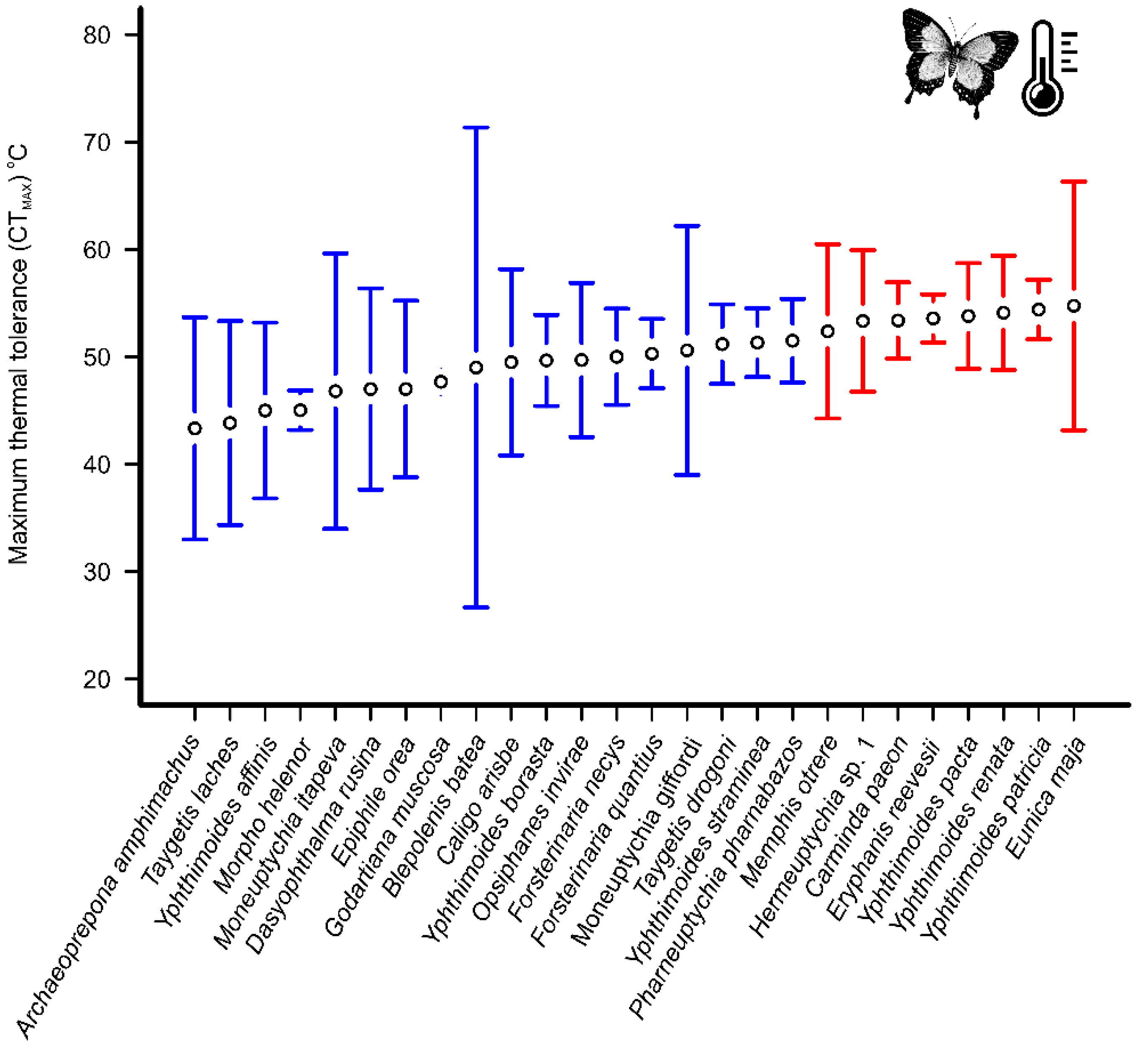
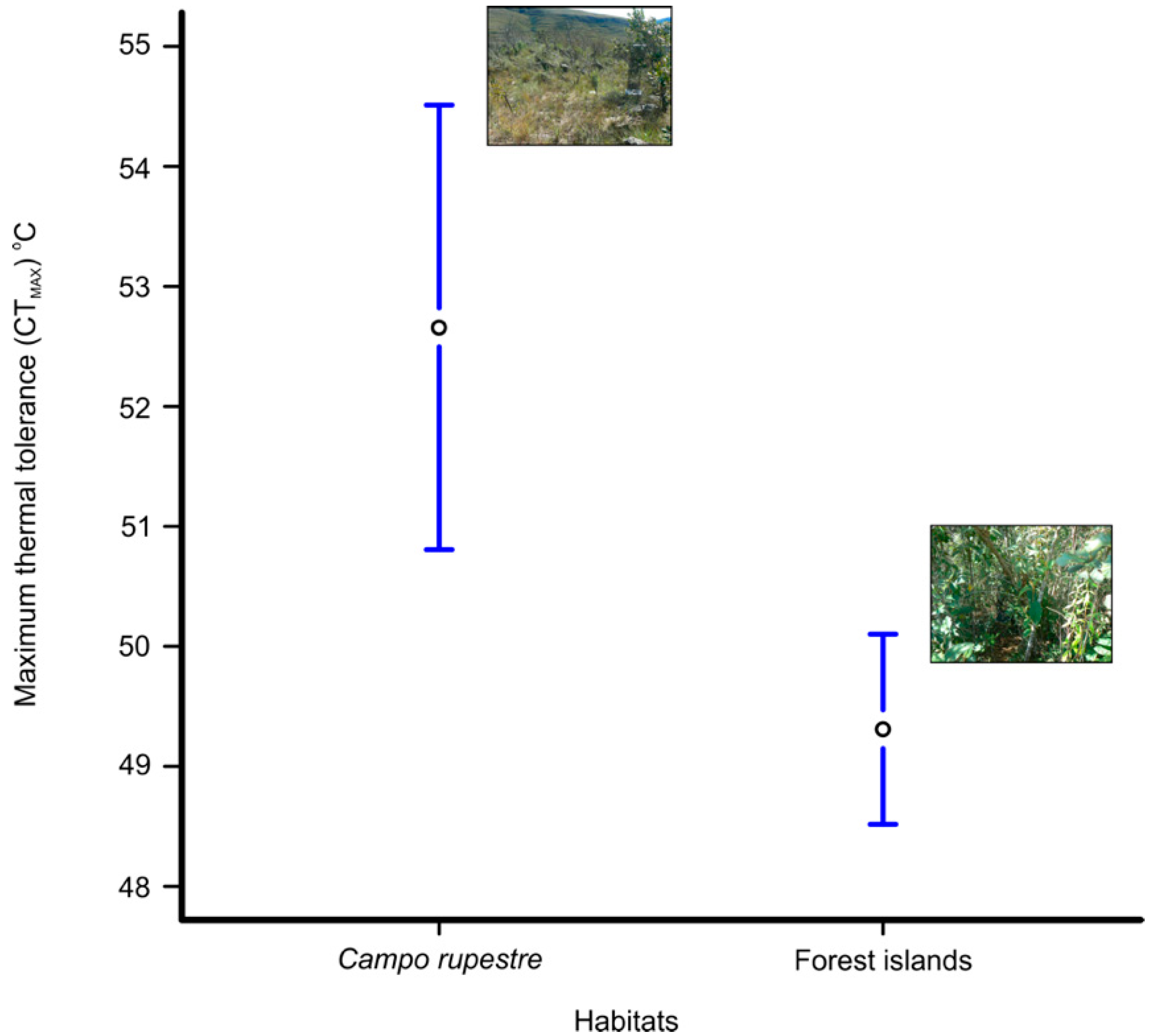
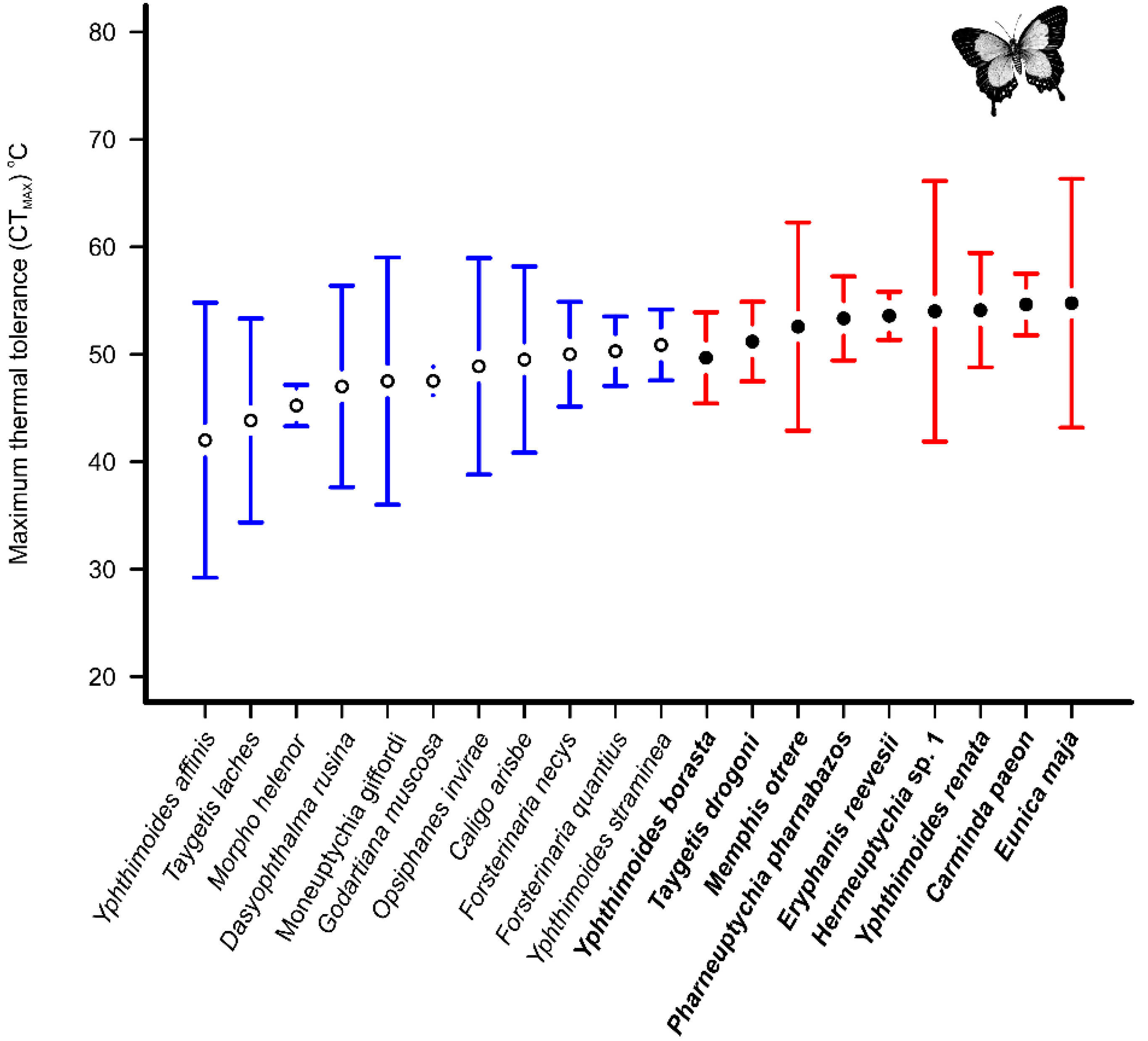
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Oms, C.S.; Cerdá, X.; Boulay, R. Is phenotypic plasticity a key mechanism for responding to thermal stress in ants? Sci. Nat. 2017, 104, 42. [Google Scholar] [CrossRef]
- Damos, P.; Savopoulou-Soultani, M. Temperature-driven models for insect development and vital thermal requirements. Psyche A J. Entomol. 2012, 2012. [Google Scholar] [CrossRef]
- Huey, R.B.; Crill, W.D.; Kingsolver, J.G.; Weber, K.E. A method for rapid measurement of heat or cold resistance of small insects. Funct. Ecol. 1992, 6, 489–494. [Google Scholar] [CrossRef] [Green Version]
- Luoto, M.; Heikkinen, R.K.; Pöyry, J.; Saarinen, K. Determinants of the biogeographical distribution of butterflies in boreal regions. J. Biogeogr. 2006, 33, 1764–1778. [Google Scholar] [CrossRef]
- Araújo, M.B.; Pearson, R.G.; Thuiller, W.; Erhard, M. Validation of species–climate impact models under climate change. Glob. Change Biol. 2005, 11, 1504–1513. [Google Scholar] [CrossRef] [Green Version]
- Baudier, K.M.; Mudd, A.E.; Erickson, S.C.; O’donnell, S. Microhabitat and body size effects on heat tolerance: Implications for responses to climate change (army ants: Formicidae, Ecitoninae). J. Anim. Ecol. 2015, 84, 1322–1330. [Google Scholar] [CrossRef] [Green Version]
- Gaston, K.J.; Chown, S.L.; Calosi, P.; Bernardo, J.; Bilton, D.T.; Clarke, A.; Clusella-Trullas, S.; Ghalambor, C.K.; Konarzewski, M.; Peck, L.S.; et al. Macrophysiology: A conceptual reunification. Am. Nat. 2009, 174, 595–612. [Google Scholar] [CrossRef]
- García-Robledo, C.; Kuprewicz, E.K.; Staines, C.L.; Erwin, T.L.; Kress, W.J. Limited tolerance by insects to high temperatures across tropical elevational gradients and the implications of global warming for extinction. Proc. Natl. Acad. Sci. USA 2016, 113, 680–685. [Google Scholar] [CrossRef] [Green Version]
- Bonebrake, T.C.; Ponisio, L.C.; Boggs, C.L.; Ehrlich, P.R. More than just indicators: A review of tropical butterfly ecology and conservation. Biol. Conserv. 2010, 143, 1831–1841. [Google Scholar] [CrossRef]
- Brown, K.S., Jr. Conservation of neotropical environments: Insects as indicators. In The Conservation of Insects and Their Habitats; Collins, N.M., Thomas, J.A., Eds.; Academic Press: London, UK, 1991. [Google Scholar]
- Ribeiro, D.B.; Freitas, A.V.L. The effect of reduced-impact logging on fruit-feeding butterflies in Central Amazon, Brazil. J. Insect Conserv. 2012, 16, 77–744. [Google Scholar] [CrossRef]
- Fernandes, G.W.; Barbosa, N.P.U.; Negreiros, D.; Paglia, A.P. Challenges for the conservation of vanishing megadiverse rupestrian grasslands. Nat. Conserv. 2014, 12, 162–165. [Google Scholar] [CrossRef] [Green Version]
- Coelho, M.S.; Fernandes, G.W.; Pacheco, P.; Diniz, V.; Meireles, A.; dos Santos, R.M.; Negreiros, D. Archipelago of montane forests surrounded by rupestrian grasslands: New insights and perspectives. In Ecology and Conservation of Mountaintop Grasslands in Brazil; Fernandez, G.W., Ed.; Springer: Cham, Switzerland, 2016; pp. 129–156. [Google Scholar]
- Conceição, A.A.; Pirani, J.R. Delimitação de Habitats em Campos Rupestres na Chapada Diamantina, Bahia: Substratos, Composição Florística e Aspectos Estruturais. Bol. Bot. Univ. São Paulo 2005, 23, 85–111. [Google Scholar] [CrossRef]
- Rapini, A.; Ribeiro, P.L.; Lamberti, S.; Pirani, J.R. A flora dos campos rupestres da Cadeia do Espinhaço. Megadiversidade 2008, 4, 16–24. [Google Scholar]
- Myers, N.; Mittermeier, R.; Mittermeier, C.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Alvares, C.A.; Stape, J.L.; Sentelhas, P.C.; Gonçalves, J.L.M.; Sparovek, G. Köppen’s climate classification map for Brazil. Meteorol. Z. 2013, 22, 711–728. [Google Scholar] [CrossRef]
- IEF—Instituto Estadual de Florestas. Relatório Técnico Para a Criação do Parque Estadual Serra do Ouro Branco, Ouro Branco e Ouro Preto; IEF: Belo Horizonte, Brazil, 2017. [Google Scholar]
- Overgaard, J.; Hoffmann, A.A.; Kristensen, T.N. Assessing population and environmental effects on thermal resistance in Drosophila melanogaster using ecologically relevant assays. J. Therm. Biol. 2012, 36, 409–416. [Google Scholar] [CrossRef]
- Uehara-Prado, M.; Freitas, A.V.L.; Francini, R.B.; Brown, K.S., Jr. Guia das borboletas frugívoras da reserva estadual do Morro Grande e região de Caucaia do Alto, Cotia, SP. Biota Neotropica 2004, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- DeVries, P.J. The Butterflies of Costa Rica and Their Natural History; Volume I: Papilionidae Pieridae Nymphalidae; Princeton University Press: Princeton, NJ, USA, 1987. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: http://www.R-project.org/ (accessed on 25 January 2019).
- Pagel, M. Inferring the historical patterns of biological evolution. Nature 1999, 401, 877–884. [Google Scholar] [CrossRef]
- Blomberg, S.P.; Garland, T., Jr.; Ives, A.R. Testing for phylogenetic signal in comparative data: Behavioral traits are more labile. Evolution 2003, 57, 717–745. [Google Scholar] [CrossRef]
- Abouheif, E. A method for testing the assumption of phylogenetic independence in comparative data. Evol. Ecol. Res. 1999, 1, 895–909. [Google Scholar]
- Gittleman, J.L.; Kot, M. Adaptation: Statistics and a null model for estimating phylogenetic effects. Syst. Zool. 1990, 39, 227–241. [Google Scholar] [CrossRef]
- Münkemüller, T.; Lavergne, S.; Bzeznik, B.; Dray, S.; Jombart, T.; Schiffers, K.; Thuiller, W. How to measure and test phylogenetic signal. Methods Ecol. Evol. 2012, 3, 743–756. [Google Scholar] [CrossRef]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Revell, L.J. Phytools: An R package for phylogenetic comparative biology (and other things). Methods Ecol. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Diamond, S.E.; Chick, L.D. Thermal specialist ant species have restricted, equatorial geographic ranges: Implications for climate change vulnerability and risk of extinction. Ecography 2017, 41, 1507–1509. [Google Scholar] [CrossRef] [Green Version]
- Marini-Filho, O.; Martins, R.P. Nymphalid butterfly dispersal among forest fragments at Serra da Canastra National Park, Brazil. J. Insect Conserv. 2010, 14, 401–411. [Google Scholar] [CrossRef]
- Grime, J.P. Benefits of plant diversity to ecosystems: Immediate, filter and founder effects. J. Ecol. 1998, 86, 902–910. [Google Scholar] [CrossRef]
- Fordyce, J.; Devries, P. A tale of two communities: Neotropical butterfly assemblages show higher beta diversity in the canopy compared to the understory. Oecologia 2016, 181. [Google Scholar] [CrossRef]
- Cros, S.; Retana, J.; Cerdá, X. Spatial and temporal variations in the activity patterns of Mediterranean ant communities. Ecoscience 1997, 4, 269–278. [Google Scholar] [CrossRef] [Green Version]
- Retana, J.; Cerdá, X. Patterns of diversity and composition of Mediterranean ground ant communities tracking spatial and temporal variability in the thermal environment. Oecologia 2000, 123, 436–444. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Maximum Thermal Tolerance (Mean ± SD) in °C | Habitats | Total | |
|---|---|---|---|---|
| Campo Rupestre | Forest Islands | |||
| Archaeoprepona amphimachus (Fabricius, 1775) | 43.3 ± 4.2 | 2 | 1 | 3 |
| Blepolenis batea (Hübner, 1821) | 49.0 ± 9 | 1 | 2 | 3 |
| Caligo arisbe (Hübner, 1822) | 49.5 ± 5.4 | 0 | 4 | 4 |
| Carminda paeon (Godart, 1824) | 53.4 ± 7.8 | 2 | 19 | 21 |
| Colobura dirce (Linnaeus, 1758) | 52 ± 2.8 | 0 | 2 | 2 |
| Dasyophthalma rusina (Godart, 1824) | 47 ± 7.5 | 0 | 5 | 5 |
| Doxocopa laurentia (Godart, 1824) | 59.0 | 1 | 0 | 1 |
| Epiphile orea (Hübner, 1823) | 47 ± 5.2 | 4 | 0 | 4 |
| Eryphanis reevesii (Doubleday, 1849) | 53.6 ± 3.7 | 0 | 13 | 13 |
| Eunica cuvierii (Godart, 1819) | 60.5 ± 2.1 | 2 | 0 | 2 |
| Eunica maja (Fabricius, 1775) | 54.8 ± 7.3 | 0 | 4 | 4 |
| Eunica tatila (Herrich-Schäffer, 1855) | 53 | 1 | 0 | 1 |
| Forsterinaria necys (Godart, 1824) | 50 ± 7.7 | 1 | 13 | 14 |
| Forsterinaria quantius (Godart, 1824) | 50.3 ± 5.6 | 0 | 14 | 14 |
| Godartiana muscosa (Butler, 1870) | 47.7 ± 6.5 | 4 | 94 | 98 |
| Hamadryas epinome (Felder and Felder, 1867) | 53 ± 4.2 | 0 | 2 | 2 |
| Hamdryas februa (Linnaeus, 1758) | 48 | 0 | 1 | 1 |
| Hermeuptychia sp1 (Forster, 1964) | 53.5 ± 6.3 | 2 | 4 | 6 |
| Historis odius (Fabricius, 1775) | 33.0 | 1 | 0 | 1 |
| Junonia evarete (Cramer, 1779) | 69.0 | 1 | 0 | 1 |
| Memphis acidalia (Hübner, 1819) | 47.0 | 1 | 0 | 1 |
| Memphis appias (Hübner, 1825) | 64.0 | 1 | 0 | 1 |
| Memphis moruus (Fabricius, 1775) | 54.5 ± 2.1 | 1 | 1 | 2 |
| Memphis otrere (Hübner, 1825) | 52.4 ± 9.7 | 1 | 7 | 8 |
| Moneuptychia giffordi (Freitas, Emery, and Mielke, 2010) | 50.6 ± 9.3 | 1 | 4 | 5 |
| Moneuptychia itapeva (Freitas, 2007) | 46.8 ± 10.3 | 4 | 1 | 5 |
| Moneuptychia walhbergi (Freitas, Barbosa, Siewert and Mielke, 2015) | 54 ± 1.4 | 0 | 2 | 2 |
| Morpho helenor (Cramer, 1776) | 45 ± 5.6 | 2 | 36 | 38 |
| Opoptera sulcius (Staudinger, 1887) | 39 ± 9.9 | 0 | 2 | 2 |
| Opsiphanes invirae (Hübner, 1808) | 49.7 ± 5.8 | 1 | 4 | 5 |
| Paryphthimoides grimon (Godart, 1824) | 40.0 | 0 | 1 | 1 |
| Paryphthimoides poltys (Prittwitz, 1865) | 47.0 | 0 | 1 | 1 |
| Pharneuptychia pharnabazos (Bryk, 1953) | 51.5 ± 4.7 | 2 | 6 | 8 |
| Taygetis drogoni (Siewert, Zacca, Dias and Freitas 2013) | 51.2 ± 5.5 | 0 | 11 | 11 |
| Taygetis laches (Fabricius, 1793) | 43.4 ± 3.8 | 0 | 3 | 3 |
| Yphthimoides affinis (Butler, 1867) | 45 ± 7.8 | 2 | 4 | 6 |
| Yphthimoides angularis (Butler, 1867) | 58.5 ± 6.4 | 2 | 0 | 2 |
| Yphthimoides borasta (Schaus, 1902) | 49.7 ± 5.5 | 0 | 9 | 9 |
| Yphthimoides pacta (Weymer, 1911) | 53.8 ± 4.0 | 4 | 1 | 5 |
| Yphthimoides patricia (Hayward, 1957) | 54.4 ± 8.1 | 34 | 1 | 35 |
| Yphthimoides renata (Stoll, 1780) | 54.1 ± 7.4 | 0 | 10 | 10 |
| Yphthimoides sp1 | 64 | 1 | 0 | 1 |
| Yphthimoides ps2 | 54 | 1 | 0 | 1 |
| Yphthimoides straminea (Butler, 1867) | 51.3 ± 8.7 | 2 | 29 | 31 |
| Zaretis isidora (Cramer, 1779) | 57.0 | 0 | 1 | 1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, V.D.e.; Beirão, M.V.; Cardoso, D.C. Thermal Tolerance of Fruit-Feeding Butterflies (Lepidoptera: Nymphalidae) in Contrasting Mountaintop Environments. Insects 2020, 11, 278. https://doi.org/10.3390/insects11050278
Silva VDe, Beirão MV, Cardoso DC. Thermal Tolerance of Fruit-Feeding Butterflies (Lepidoptera: Nymphalidae) in Contrasting Mountaintop Environments. Insects. 2020; 11(5):278. https://doi.org/10.3390/insects11050278
Chicago/Turabian StyleSilva, Vanessa Diniz e, Marina Vale Beirão, and Danon Clemes Cardoso. 2020. "Thermal Tolerance of Fruit-Feeding Butterflies (Lepidoptera: Nymphalidae) in Contrasting Mountaintop Environments" Insects 11, no. 5: 278. https://doi.org/10.3390/insects11050278
APA StyleSilva, V. D. e., Beirão, M. V., & Cardoso, D. C. (2020). Thermal Tolerance of Fruit-Feeding Butterflies (Lepidoptera: Nymphalidae) in Contrasting Mountaintop Environments. Insects, 11(5), 278. https://doi.org/10.3390/insects11050278