The Intracellular Symbiont Wolbachia pipientis Enhances Recombination in a Dose-Dependent Manner


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fly Rearing
2.2. Spiroplasma Methods
2.3. DNA Extraction and Polymerase Chain Reaction
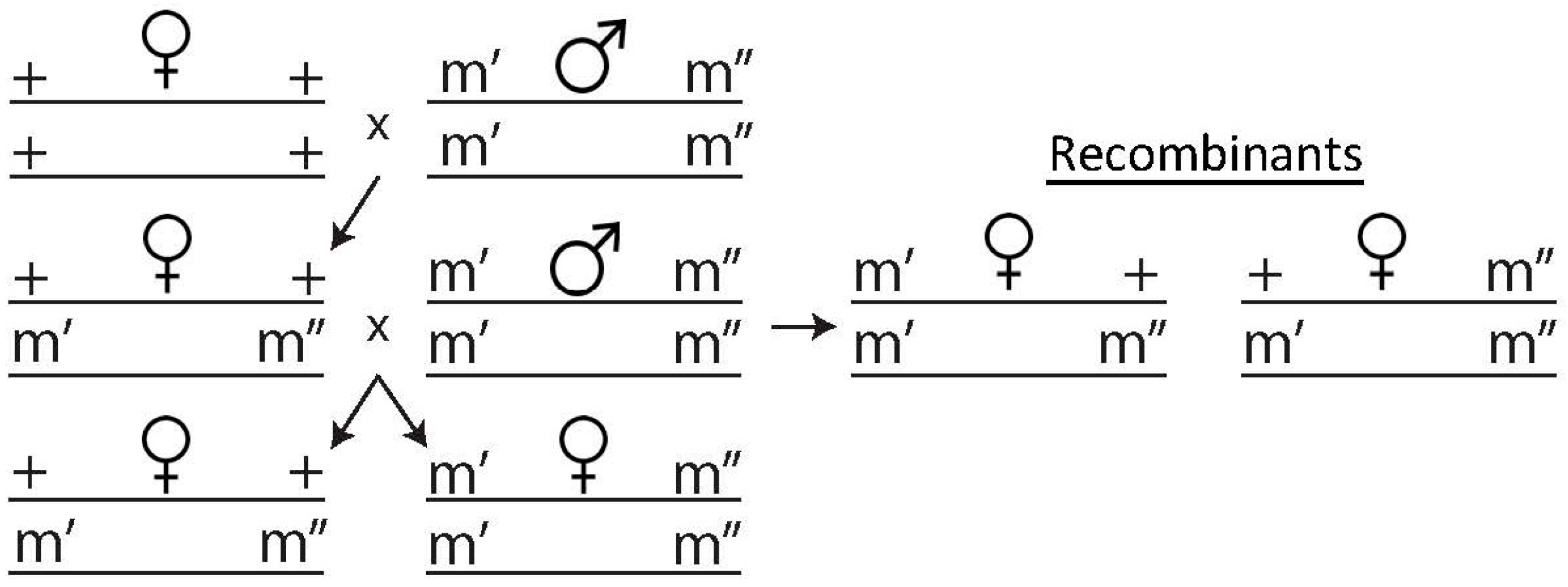
2.4. Recombination Assay
3. Results
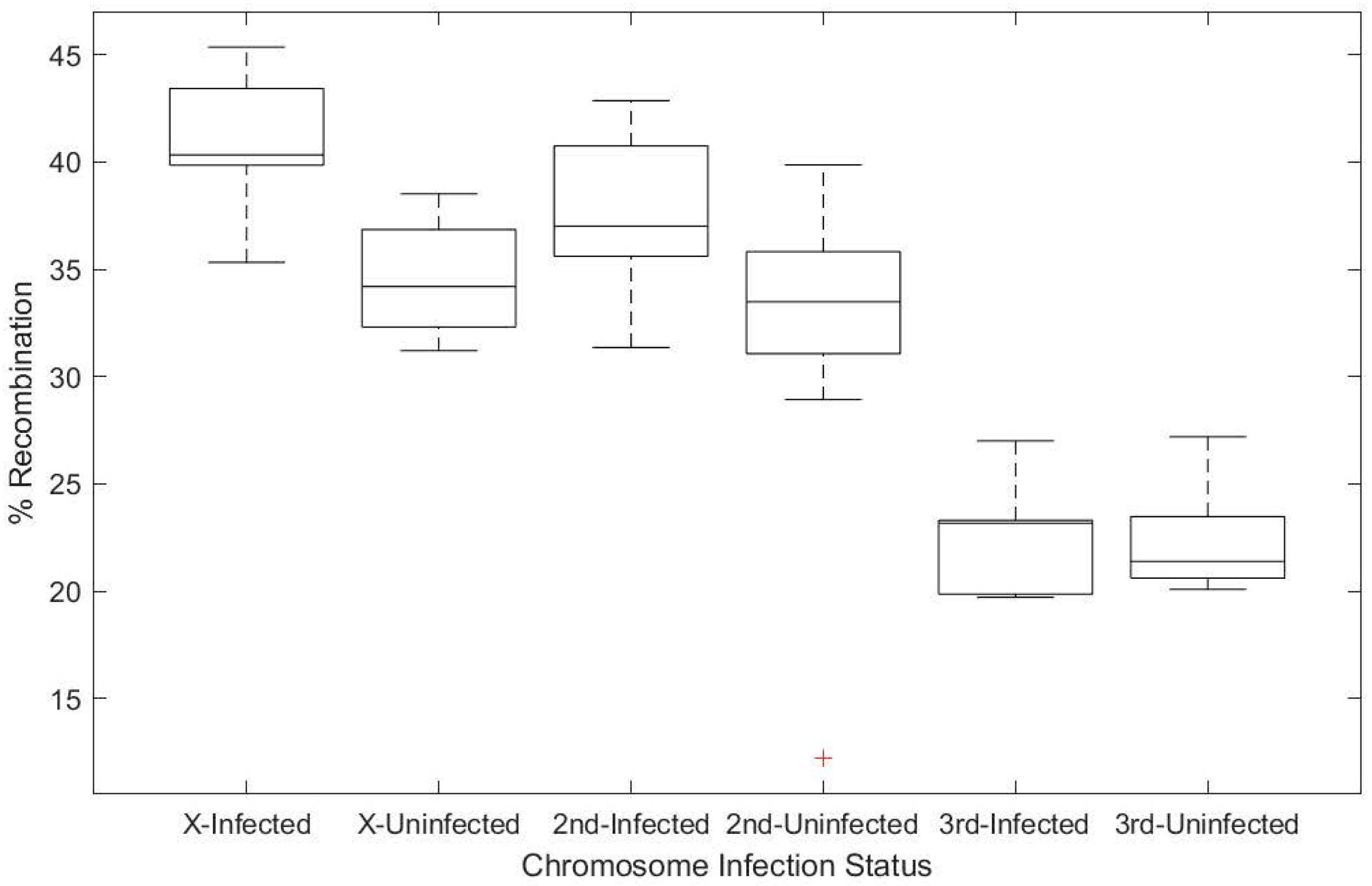
3.1. Wolbachia Infection Increases Host Recombination Rate
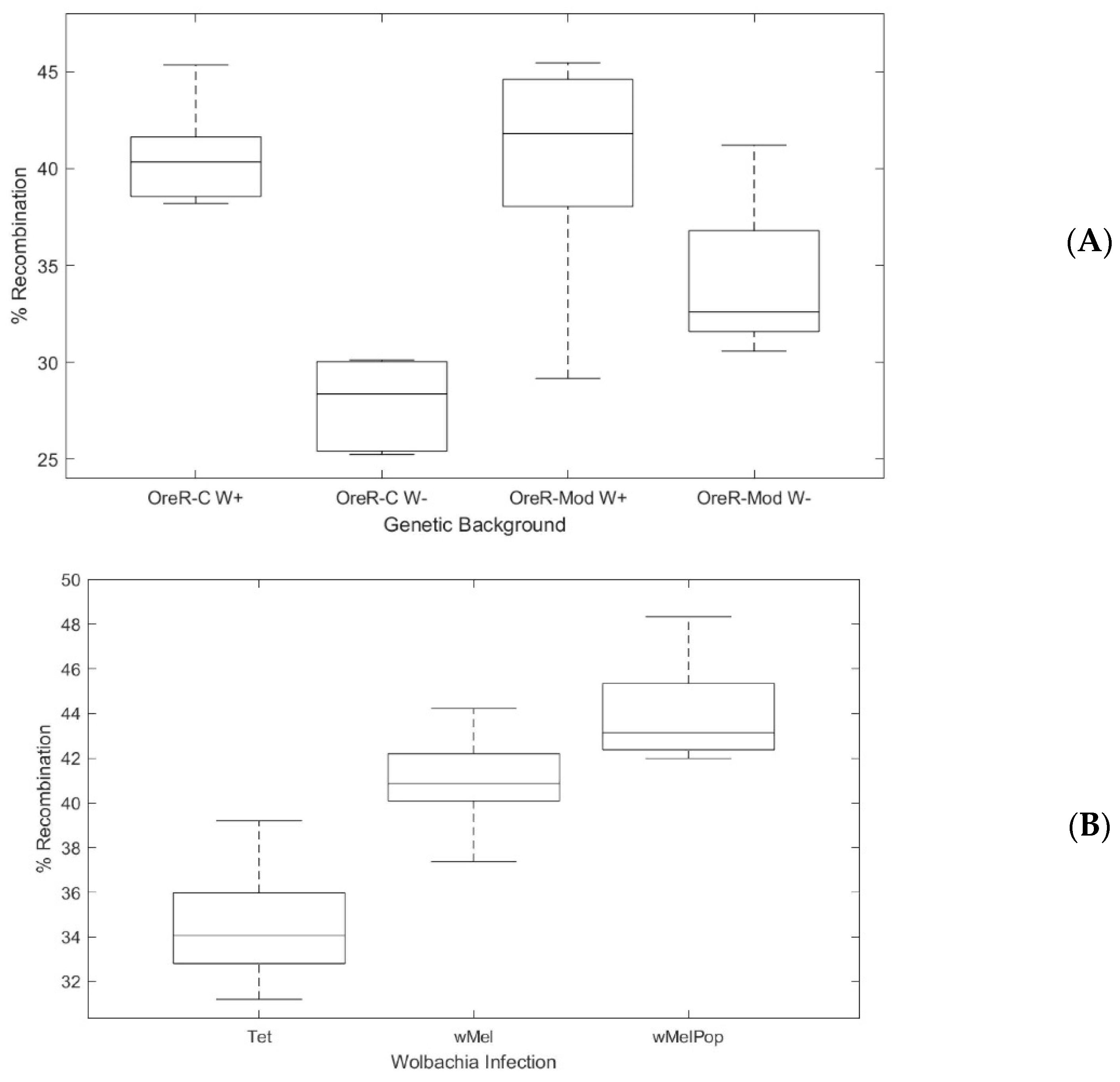
3.2. Dose-Dependent Effect of Wolbachia on Host Recombination Rate
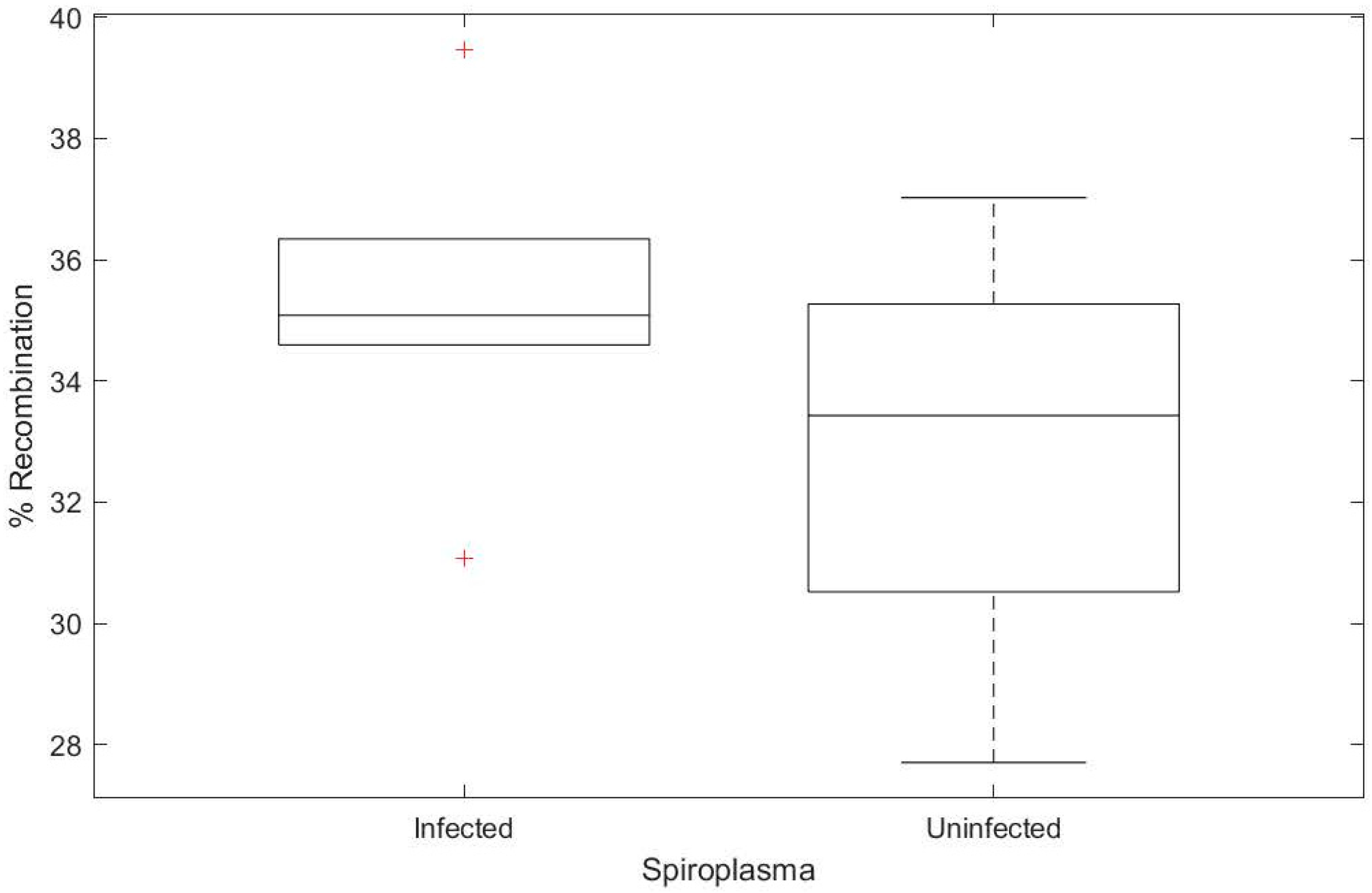
3.3. Spiroplasma Does Not Increase Host Recombination Rate
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Deposition and Reagent Availability
References
- Haddrill, P.R.; Halligan, D.L.; Tomaras, D.; Charlesworth, B. Reduced efficacy of selection in regions of the Drosophila genome that lack crossing over. Genome Biol. 2007, 8, R18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGaugh, S.E.; Heil, C.S.; Manzano-Winkler, B.; Loewe, L.; Goldstein, S.; Himmel, T.L.; Noor, M.A. Recombination modulates how selection affects linked sites in Drosophila. PLoS Biol. 2012, 10, e1001422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Presgraves, D.C. Recombination enhances protein adaptation in Drosophila melanogaster. Curr Biol. 2005, 15, 1651–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, N.D.; Criscoe, D.R.; Skolfield, S.; Kohl, K.P.; Keebaugh, E.S.; Schlenke, T. Fruit flies diversify their offspring in response to parasite infection. Science 2015, 349, 747–750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zug, R.; Hammerstein, P. Still a host of hosts for Wolbachia. analysis of recent data suggests that 40% of terrestrial arthropod species are infected. PLoS ONE 2012, 7, e38544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werren, J.H.; Windsor, D.; Guo, L.R. Distribution of Wolbachia among neotropical arthropods. Proc. R. Soc. Lond. Ser. B 1995, 262, 197–204. [Google Scholar]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef]
- Hamm, C.A.; Begun, D.J.; Vo, A.; Smith, C.C.; Saelao, P.; Shaver, A.O.; Jaenike, J.; Turelli, M. Wolbachia do not live by reproductive manipulation alone: Infection polymorphism in Drosophila suzukii and D. subpulchrella. Mol. Ecol. 2014, 23, 4871–4885. [Google Scholar] [CrossRef] [Green Version]
- Zug, R.; Hammerstein, P. Bad guys turned nice? A critical assessment of Wolbachia mutualisms in arthropod hosts. Biol. Rev. Camb. Philos Soc. 2015, 90, 89–111. [Google Scholar] [CrossRef]
- Hosokawa, T.; Koga, R.; Kikuchi, Y.; Meng, X.Y.; Fukatsu, T. Wolbachia as a bacteriocyte-associated nutritional mutualist. Proc. Natl. Acad. Sci. USA 2010, 107, 769–774. [Google Scholar] [CrossRef] [Green Version]
- Chrostek, E.; Marialva, M.S.; Esteves, S.S.; Weinert, L.A.; Martinez, J.; Jiggins, F.M.; Teixeira, L. Wolbachia variants induce differential protection to viruses in Drosophila melanogaster: A phenotypic and phylogenomic analysis. PLoS Genet. 2013, 9, e1003896. [Google Scholar] [CrossRef] [Green Version]
- Martinez, J.; Longdon, B.; Bauer, S.; Chan, Y.S.; Miller, W.J.; Bourtzis, K.; Teixeira, L.; Jiggins, F.M. Symbionts commonly provide broad spectrum resistance to viruses in insects: A comparative analysis of Wolbachia strains. PLoS Pathog. 2014, 10, e1004369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teixeira, L.; Ferreira, A.; Ashburner, M. The bacterial symbiont Wolbachia induces resistance to RNA viral infections in Drosophila melanogaster. PLoS Biol. 2008, 6, e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Neill, S.L. The Use of Wolbachia by the World Mosquito Program to Interrupt Transmission of Aedes aegypti Transmitted Viruses. Adv. Exp. Med. Biol. 2018, 1062, 355–360. [Google Scholar] [PubMed]
- Fry, A.J.; Rand, D.M. Wolbachia Interactions That Determine Drosophila melanogaster Survival. Evolution 2002, 56, 1976–1981. [Google Scholar] [CrossRef]
- Fry, A.J.; Palmer, M.R.; Rand, D.M. Variable fitness effects of Wolbachia infection in Drosophila melanogaster. Heredity 2004, 93, 379–389. [Google Scholar] [CrossRef] [Green Version]
- Starr, D.J.; Cline, T.W. A host parasite interaction rescues Drosophila oogenesis defects. Nature 2002, 418, 76–79. [Google Scholar] [CrossRef]
- Dedeine, F.; Vavre, F.; Shoemaker, D.D.; Boulétreau, M. Intra-individual coexistence of a wolbachia strain required for host oogenesis with two strains inducing cytoplasmic incompatibility in the wasp asobara tabida. Evolution 2004, 58, 2167–2174. [Google Scholar] [CrossRef]
- Hunter, C.M.; Huang, W.; Mackay, T.F.; Singh, N.D. The Genetic Architecture of Natural Variation in Recombination Rate in Drosophila melanogaster. PLoS Genet. 2016, 12, e1005951. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.D. Wolbachia Infection Associated with Increased Recombination in Drosophila. G3 (Bethesda) 2019, 9, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Herren, J.K.; Paredes, J.C.; Schupfer, F.; Arafah, K.; Bulet, P.; Lemaitre, B. Insect endosymbiont proliferation is limited by lipid availability. Elife 2014, 3, e02964. [Google Scholar] [CrossRef] [PubMed]
- Herren, J.K.; Lemaitre, B. Spiroplasma and host immunity: Activation of humoral immune responses increases endosymbiont load and susceptibility to certain Gram-negative bacterial pathogens in Drosophila melanogaster. Cell Microbiol. 2011, 13, 1385–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anbutsu, H.; Fukatsu, T. Evasion, suppression and tolerance of Drosophila innate immunity by a male-killing Spiroplasma endosymbiont. Insect. Mol. Biol. 2010, 19, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Mackay, T.F.; Richards, S.; Stone, E.A.; Barbadilla, A.; Ayroles, J.F.; Zhu, D.; Casillas, S.; Han, Y.; Magwire, M.M.; Cridland, J.M.; et al. The Drosophila melanogaster Genetic Reference Panel. Nature 2012, 482, 173–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chrostek, E.; Teixeira, L. Mutualism breakdown by amplification of Wolbachia genes. PLoS Biol. 2015, 13, e1002065. [Google Scholar] [CrossRef] [Green Version]
- Hamblin, M.T.; Aquadro, C.F. High nucleotide sequence variation in a region of low recombination in Drosophila simulans is consistent with the background selection model. Mol. Biol. Evol. 1996, 13, 1133–1140. [Google Scholar] [CrossRef] [Green Version]
- Aquadro, C.F.; Begun, D.J.; Kindahl, E.C. Selection, recombination, and DNA polymorphism in Drosophila; Springer: Boston, MA, USA, 1994; pp. 46–56. [Google Scholar]
- Lindsey, A.R.I.; Rice, D.W.; Bordenstein, S.R.; Brooks, A.W.; Bordenstein, S.R.; Newton, I.L.G. Evolutionary Genetics of Cytoplasmic Incompatibility Genes cifA and cifB in Prophage WO of Wolbachia. Genome Biol. Evol. 2018, 10, 434–451. [Google Scholar] [CrossRef]
- Chen, H.; Ronau, J.A.; Beckmann, J.F.; Hochstrasser, M. A Wolbachia nuclease and its binding partner provide a distinct mechanism for cytoplasmic incompatibility. Proc. Natl. Acad. Sci. USA 2019, 116, 22314–22321. [Google Scholar] [CrossRef]
- Rice, D.W.; Sheehan, K.B.; Newton, I.L.G. Large-Scale Identification of Wolbachia pipientis Effectors. Genome Biol. Evol. 2017, 9, 1925–1937. [Google Scholar] [CrossRef] [Green Version]
- Lindsey, A.R.; Werren, J.H.; Richards, S.; Stouthamer, R. Comparative Genomics of a Parthenogenesis-Inducing Wolbachia Symbiont. G3 (Bethesda) 2016, 6, 2113–2123. [Google Scholar] [CrossRef]
- Truitt, A.M.; Kapun, M.; Kaur, R.; Miller, W.J. Wolbachia modifies thermal preference in Drosophila melanogaster. Environ. Microbiol. 2019, 21, 3259–3268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parsons, P.A. Evolutionary Rates-Effects of Stress Upon Recombination. Biol. J. Linn. Soc. 1988, 35, 49–68. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bryant, K.N.; Newton, I.L.G. The Intracellular Symbiont Wolbachia pipientis Enhances Recombination in a Dose-Dependent Manner. Insects 2020, 11, 284. https://doi.org/10.3390/insects11050284
Bryant KN, Newton ILG. The Intracellular Symbiont Wolbachia pipientis Enhances Recombination in a Dose-Dependent Manner. Insects. 2020; 11(5):284. https://doi.org/10.3390/insects11050284
Chicago/Turabian StyleBryant, Kaeli N., and Irene L. G. Newton. 2020. "The Intracellular Symbiont Wolbachia pipientis Enhances Recombination in a Dose-Dependent Manner" Insects 11, no. 5: 284. https://doi.org/10.3390/insects11050284
APA StyleBryant, K. N., & Newton, I. L. G. (2020). The Intracellular Symbiont Wolbachia pipientis Enhances Recombination in a Dose-Dependent Manner. Insects, 11(5), 284. https://doi.org/10.3390/insects11050284