Detection of Wolbachia Infections in Natural and Laboratory Populations of the Moroccan Hessian Fly, Mayetiola destructor (Say)

 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and DNA Isolation
2.2. Screening of Reproductive Symbionts and Sanger Sequencing
2.3. PCR Amplification of V3-V4 Region, PCR Indexing and Illumina Sequencing
2.4. Bioinformatic Analysis of Amplicon Sequencing Data
2.5. Phylogenetic Analysis
3. Results
3.1. Reproductive Infection Status Assessed by PCR Screening of Natural and Laboratory Hessian Fly Populations
3.1.1. Reproduction Infection Prevalence in Natural and Laboratory Hessian Fly Populations
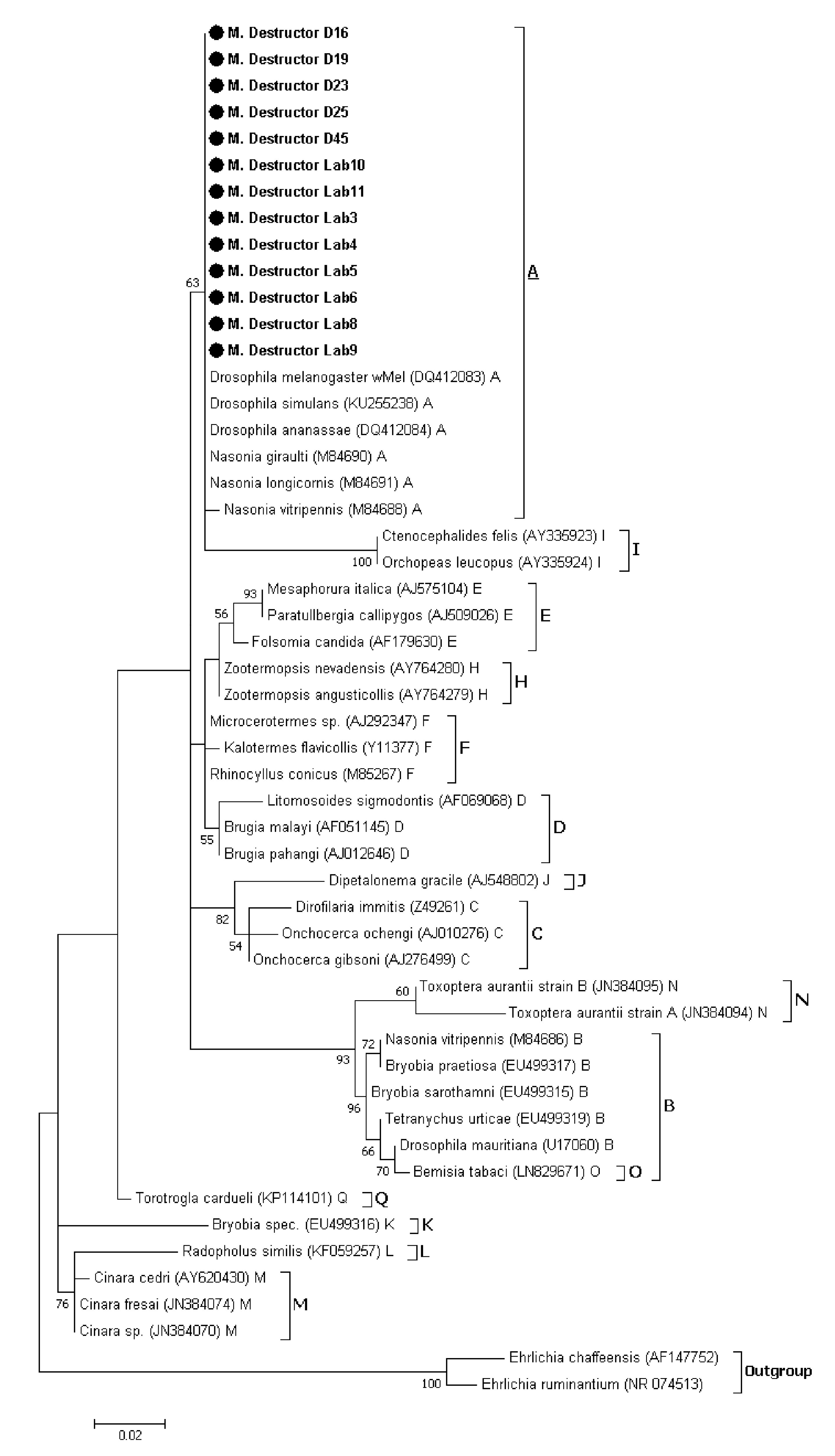
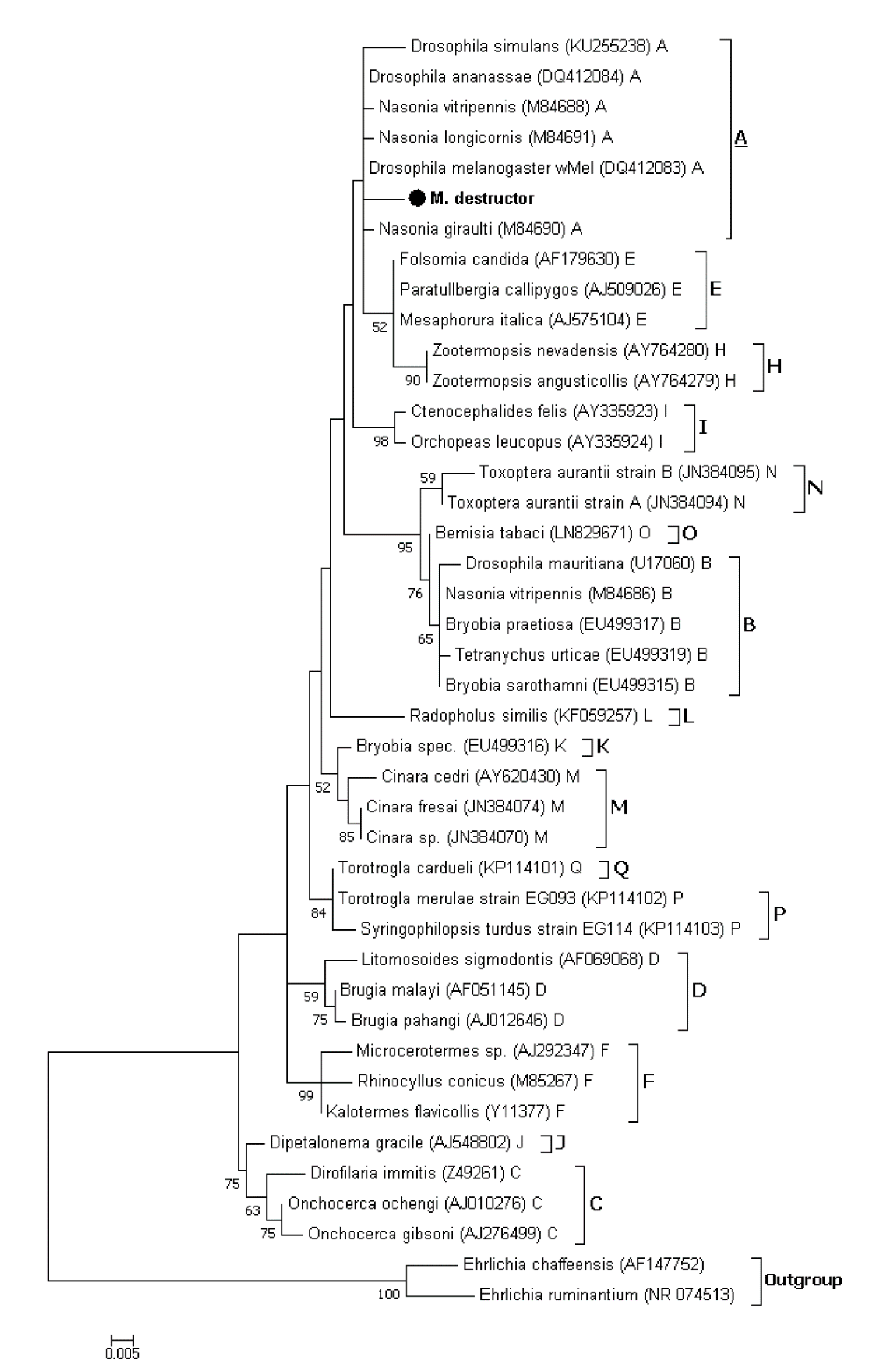
3.1.2. Phylogenetic Analysis of Wolbachia Sequences Obtained by Sanger Sequencing
3.2. 16S rRNA Gene Amplicon Sequencing Reveals the Presence of Wolbachia in Natural Populations of Hessian Fly and Dynamics Indicate an Origin Effect
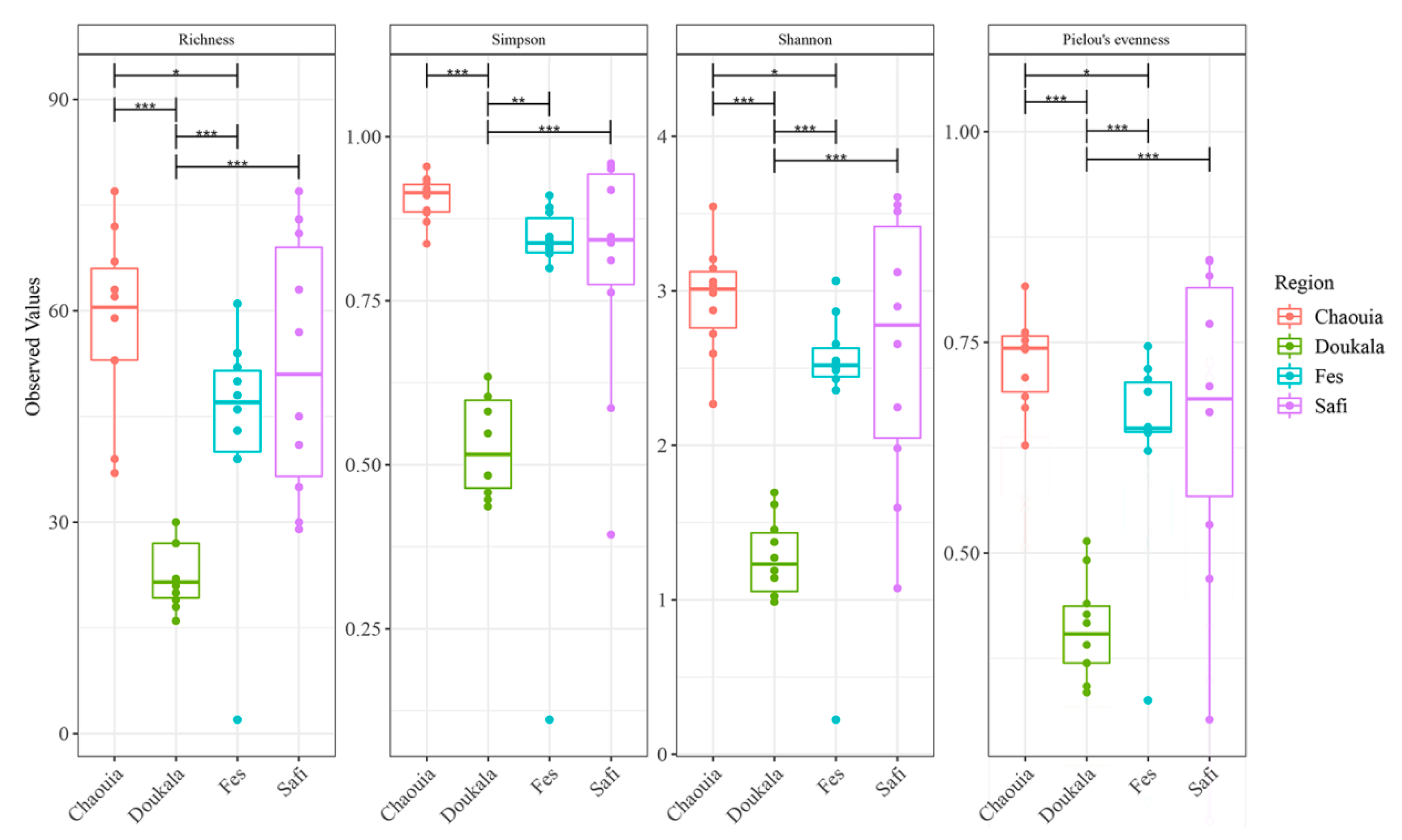
3.2.1. Bacterial Diversity within Hessian Flies’ Natural Populations
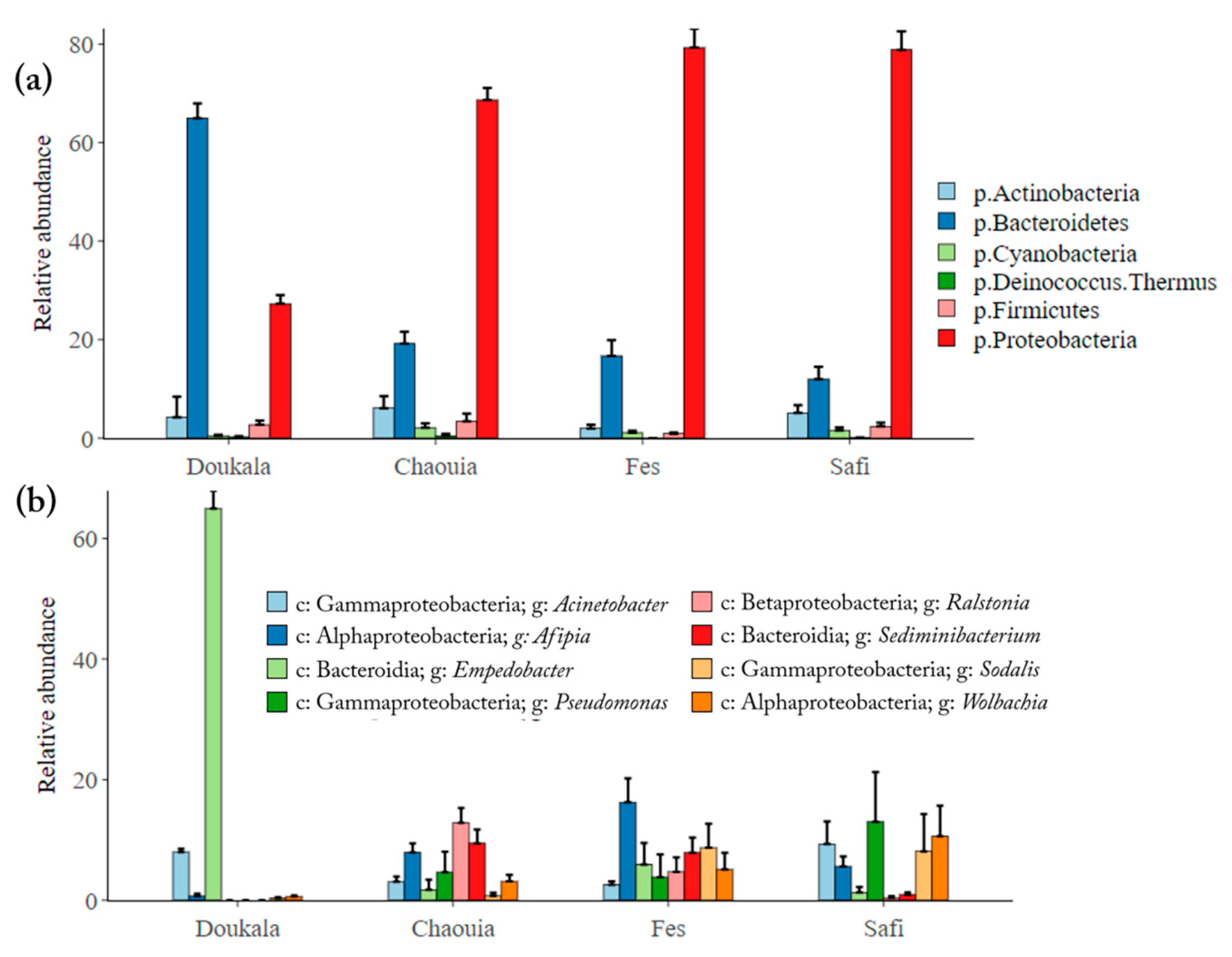
3.2.2. Bacterial Composition of Hessian Flies’ Natural Populations
3.2.3. Phylogenetic Analysis of Wolbachia Related Sequence Obtained by HTS
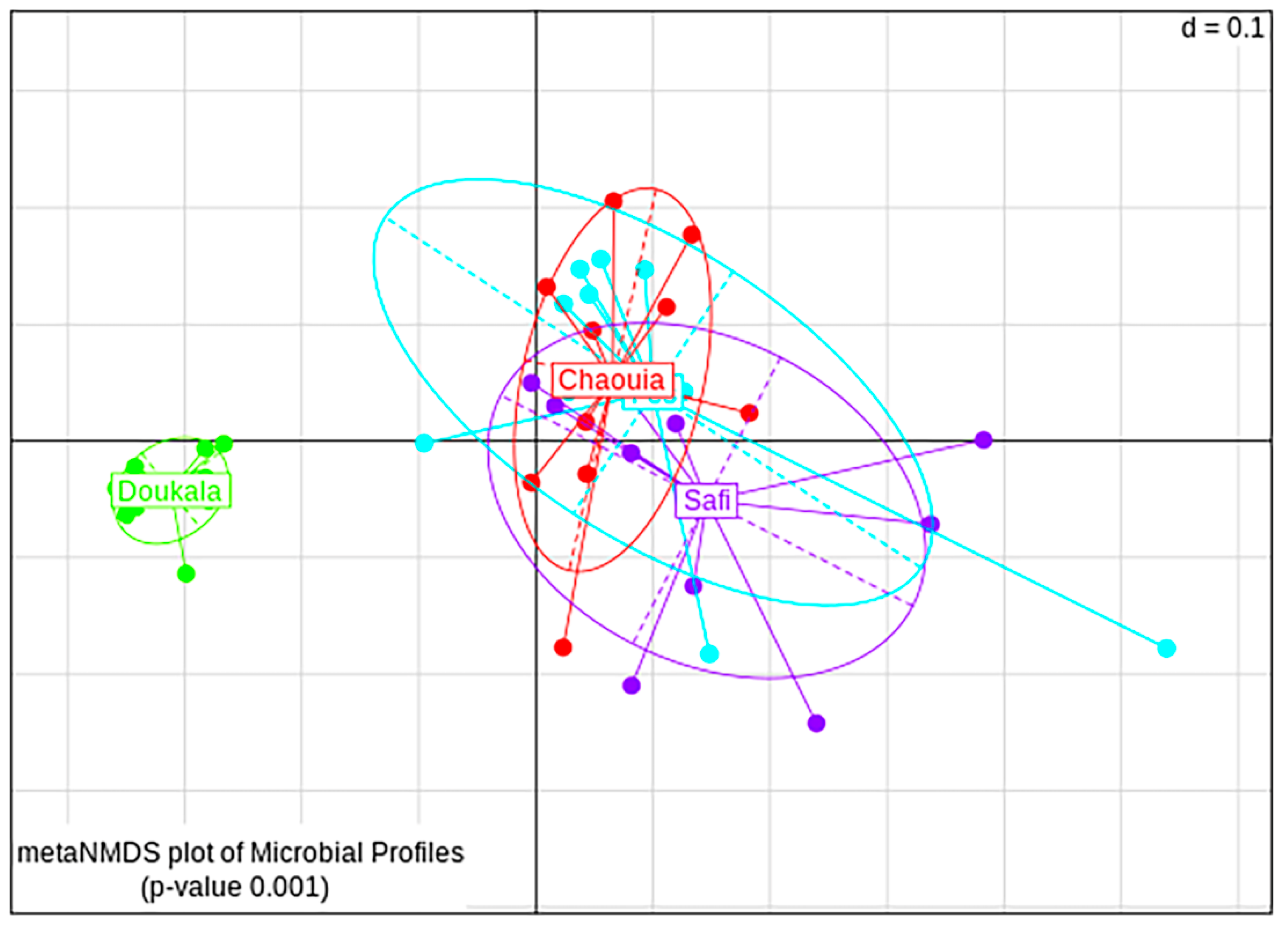
3.2.4. Bacterial Diversity between Hessian Flies’ Natural Populations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- El Bouhssini, M.; Ogbonnaya, F.C.; Chen, M.; Lhaloui, S.; Rihawi, F.; Dabbous, A. Sources of resistance in primary synthetic hexaploid wheat (Triticum aestivum L.) to insect pests: Hessian fly, Russian wheat aphid and Sunn pest in the fertile crescent. Genet. Resour. Crop Evol. 2013, 60, 621–627. [Google Scholar] [CrossRef]
- Lhaloui, S.; Buschman, L.; El Bouhssini, M.; Starks, K.; Keith, D.L.; El Houssaini, K. Control of Mayetiola species (Diptera: Cecidomyiidae) with carbofuran in bread wheat, durum wheat and barley with yield loss assessment and its economic analysis. Al Awamia 1992, 77, 55–73. [Google Scholar]
- Lhaloui, S. Biology, Host Preference, Host Suitability, and Plant. Resistance Studies of the Barley Stem Gall. Midge and Hessian Fly (Diptera: Cecidomyiidae) in Morocco; Kansas State University: Manhattan, KS, USA, 1995. [Google Scholar]
- Harris, M.O.; Rose, S. Factors influencing the onset of egglaying in a cecidomyiid fly. Physiol. Entomol. 1991, 16, 183–190. [Google Scholar] [CrossRef]
- Bergh, J.C.; Harris, M.O.; Rose, S. Temporal Patterns of Emergence and Reproductive Behavior of the Hessian Fly (Diptera: Cecidomyiidae). Ann. Entomol. Soc. Am. 1990, 83, 998–1004. [Google Scholar] [CrossRef]
- Stuart, J.J.; Chen, M.-S.; Shukle, R.; Harris, M.O. Gall Midges (Hessian Flies) as Plant Pathogens. Annu. Rev. Phytopathol. 2012, 50, 339–357. [Google Scholar] [CrossRef]
- Shukle, R.H. Hessian Fly, Mayetiola destructor (Say) (Diptera: Cecidomyiidae). In Encyclopedia of Entomology; Capinera, J.L., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 1794–1797. ISBN 978-1-4020-6359-6. [Google Scholar]
- Gagné, R.J.; Hatchett, J.H. Instars of the Hessian Fly (Diptera: Cecidomyiidae). Ann. Entomol. Soc. Am. 1989, 82, 73–79. [Google Scholar] [CrossRef]
- Schmid, R.B.; Knutson, A.; Giles, K.L.; McCornack, B.P. Hessian Fly (Diptera: Cecidomyiidae) Biology and Management in Wheat. J. Integr. Pest. Manag. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Naber, N.; El Bouhssini, M.; Lhaloui, S. Biotypes of Hessian fly (Dipt., Cecidomyiidae) in Morocco. J. Appl. Entomol. 2003, 127, 174–176. [Google Scholar] [CrossRef]
- El Bouhssini, M.; Lhaloui, S.; Amri, A.; Jlibene, M.; Hatchett, J.H.; Nssarellah, N.; Nachitt, M. Wheat genetic control of Hessian fly (Diptera: Cecidomyiidae) in Morocco. Field Crops Res. 1996, 45, 111–114. [Google Scholar] [CrossRef]
- Ami, E.B.; Yuval, B.; Jurkevitch, E. Manipulation of the microbiota of mass-reared Mediterranean fruit flies Ceratitis capitata (Diptera: Tephritidae) improves sterile male sexual performance. ISME J. 2010, 4, 28–37. [Google Scholar] [CrossRef]
- Augustinos, A.A.; Kyritsis, G.A.; Papadopoulos, N.T.; Abd-Alla, A.M.M.; Cáceres, C.; Bourtzis, K. Exploitation of the Medfly Gut Microbiota for the Enhancement of Sterile Insect Technique: Use of Enterobacter sp. in Larval Diet-Based Probiotic Applications. PLoS ONE 2015, 10, e0136459. [Google Scholar] [CrossRef] [Green Version]
- Ben-Yosef, M.; Pasternak, Z.; Jurkevitch, E.; Yuval, B. Symbiotic bacteria enable olive flies (Bactrocera oleae) to exploit intractable sources of nitrogen. J. Evol. Biol. 2014, 27, 2695–2705. [Google Scholar] [CrossRef] [PubMed]
- Gavriel, S.; Jurkevitch, E.; Gazit, Y.; Yuval, B. Bacterially enriched diet improves sexual performance of sterile male Mediterranean fruit flies. J. Appl. Entomol. 2011, 135, 564–573. [Google Scholar] [CrossRef]
- Gavriel, S.; Gazit, Y.; Yuval, B. Effect of diet on survival, in the laboratory and the field, of sterile male Mediterranean fruit flies. Entomol. Exp. Appl. 2010, 135, 96–104. [Google Scholar] [CrossRef]
- Kyritsis, G.A.; Augustinos, A.A.; Cáceres, C.; Bourtzis, K. Medfly Gut Microbiota and Enhancement of the Sterile Insect Technique: Similarities and Differences of Klebsiella oxytoca and Enterobacter sp. AA26 Probiotics during the Larval and Adult Stages of the VIENNA 8D53+ Genetic Sexing Strain. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Contreras, M.; Vlisidou, I. The Diversity of Insect-bacteria Interactions and its Applications for Disease Control. Biotechnol. Genet. Eng. Rev. 2008, 25, 203–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourtzis, K. Wolbachia-Based Technologies for Insect Pest Population Control. In Transgenesis and the Management of Vector-Borne Disease; Aksoy, S., Ed.; Springer: New York, NY, USA, 2008; Volume 627, pp. 104–113. ISBN 978-0-387-78224-9. [Google Scholar]
- Jeyaprakash, A.; Hoy, M.A. Long PCR improves Wolbachia DNA amplification: Wsp sequences found in 76% of sixty-three arthropod species. Insect Mol. Biol. 2000, 9, 393–405. [Google Scholar] [CrossRef] [PubMed]
- Saridaki, A.; Bourtzis, K. Wolbachia: More than just a bug in insects genitals. Curr. Opin. Microbiol. 2010, 13, 67–72. [Google Scholar] [CrossRef]
- Bourtzis, K. Wolbachia-Induced Cytoplasmic Incompatibility to Control Insect Pests? In Area-Wide Control of Insect Pests; Vreysen, M.J.B., Robinson, A.S., Hendrichs, J., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 125–135. ISBN 978-1-4020-6058-8. [Google Scholar]
- Bourtzis, K.; Braig, H.R.; Karr, T.L. Cytoplasmic Incompatibility. In Insect Symbiosis; Bourtzis, K., Miller, T.A., Eds.; CRC Press: Boca Raton, FL, USA, 2003; Volume 1, pp. 217–246. ISBN 0-8493-1286-8. [Google Scholar]
- Correa, C.C.; Ballard, J.W.O. Wolbachia Associations with Insects: Winning or Losing Against a Master Manipulator. Front. Ecol. Evol. 2016, 3. [Google Scholar] [CrossRef] [Green Version]
- Braquart-Varnier, C.; Altinli, M.; Pigeault, R.; Chevalier, F.D.; Grève, P.; Bouchon, D.; Sicard, M. The Mutualistic Side of Wolbachia-Isopod Interactions: Wolbachia Mediated Protection against Pathogenic Intracellular Bacteria. Front. Microbiol. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Nikoh, N.; Hosokawa, T.; Moriyama, M.; Oshima, K.; Hattori, M.; Fukatsu, T. Evolutionary origin of insect–Wolbachia nutritional mutualism. Proc. Natl. Acad. Sci. USA 2014, 111, 10257–10262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, G.; Xu, Y.; Lu, P.; Xie, Y.; Xi, Z. The Endosymbiotic Bacterium Wolbachia Induces Resistance to Dengue Virus in Aedes aegypti. PLoS Pathog. 2010, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glaser, R.L.; Meola, M.A. The Native Wolbachia Endosymbionts of Drosophila melanogaster and Culex quinquefasciatus Increase Host Resistance to West Nile Virus Infection. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osborne, S.E.; Leong, Y.S.; O’Neill, S.L.; Johnson, K.N. Variation in Antiviral Protection Mediated by Different Wolbachia Strains in Drosophila simulans. PLoS Pathog. 2009, 5. [Google Scholar] [CrossRef] [Green Version]
- Hedges, L.M.; Brownlie, J.C.; O’Neill, S.L.; Johnson, K.N. Wolbachia and Virus Protection in Insects. Science 2008, 322, 702. [Google Scholar] [CrossRef]
- Dobson, S.L.; Rattanadechakul, W.; Marsland, E.J. Fitness advantage and cytoplasmic incompatibility in Wolbachia single- and superinfected Aedes albopictus. Heredity 2004, 93, 135–142. [Google Scholar] [CrossRef]
- Brownlie, J.C.; Cass, B.N.; Riegler, M.; Witsenburg, J.J.; Iturbe-Ormaetxe, I.; McGraw, E.A.; O’Neill, S.L. Evidence for Metabolic Provisioning by a Common Invertebrate Endosymbiont, Wolbachia pipientis, during Periods of Nutritional Stress. PLoS Pathog. 2009, 5. [Google Scholar] [CrossRef] [Green Version]
- Jiggins, F.M.; Hurst, G.D.; Jiggins, C.D.; vd Schulenburg, J.H.; Majerus, M.E. The butterfly Danaus chrysippus is infected by a male-killing Spiroplasma bacterium. Parasitology 2000, 120, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Xie, J.; Butler, S.; Sanchez, G.; Mateos, M. Male killing Spiroplasma protects Drosophila melanogaster against two parasitoid wasps. Heredity 2014, 112, 399. [Google Scholar] [CrossRef]
- Łukasik, P.; Guo, H.; van Asch, M.; Ferrari, J.; Godfray, H.C.J. Protection against a fungal pathogen conferred by the aphid facultative endosymbionts Rickettsia and Spiroplasma is expressed in multiple host genotypes and species and is not influenced by co-infection with another symbiont. J. Evol. Biol. 2013, 26, 2654–2661. [Google Scholar] [CrossRef]
- Xie, J.; Vilchez, I.; Mateos, M. Spiroplasma Bacteria Enhance Survival of Drosophila hydei Attacked by the Parasitic Wasp Leptopilina heterotoma. PLoS ONE 2010, 5, e12149. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-K.; Chen, Y.-T.; Yang, K.; Hong, X.-Y. A review of prevalence and phylogeny of the bacterial symbiont Cardinium in mites (subclass: Acari). Syst. Appl. Acarol. 2016, 21, 978–990. [Google Scholar] [CrossRef] [Green Version]
- Kageyama, D.; Narita, S.; Watanabe, M. Insect Sex Determination Manipulated by Their Endosymbionts: Incidences, Mechanisms and Implications. Insects 2012, 3, 161–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giorgini, M.; Monti, M.M.; Caprio, E.; Stouthamer, R.; Hunter, M.S. Feminization and the collapse of haplodiploidy in an asexual parasitoid wasp harboring the bacterial symbiont Cardinium. Heredity 2009, 102, 365–371. [Google Scholar] [CrossRef] [Green Version]
- Provencher, L.M.; Morse, G.E.; Weeks, A.R.; Normark, B.B. Parthenogenesis in the Aspidiotus nerii Complex (Hemiptera: Diaspididae): A Single Origin of a Worldwide, Polyphagous Lineage Associated with Cardinium Bacteria. Ann. Entomol. Soc. Am. 2005, 98, 629–635. [Google Scholar] [CrossRef] [Green Version]
- Zchori-Fein, E.; Perlman, S.J.; Kelly, S.E.; Katzir, N.; Hunter, M.S. Characterization of a ‘Bacteroidetes’ symbiont in Encarsia wasps (Hymenoptera: Aphelinidae): Proposal of ‘Candidatus Cardinium hertigii’. Int. J. Syst. Evol. Microbiol. 2004, 54, 961–968. [Google Scholar] [CrossRef]
- Duron, O.; Bouchon, D.; Boutin, S.; Bellamy, L.; Zhou, L.; Engelstädter, J.; Hurst, G.D. The diversity of reproductive parasites among arthropods: Wolbachiado not walk alone. BMC Biol. 2008, 6, 27. [Google Scholar] [CrossRef] [Green Version]
- Ferree, P.M.; Avery, A.; Azpurua, J.; Wilkes, T.; Werren, J.H. A Bacterium Targets Maternally Inherited Centrosomes to Kill Males in Nasonia. Curr. Biol. 2008, 18, 1409–1414. [Google Scholar] [CrossRef] [Green Version]
- Bansal, R.; Hulbert, S.; Schemerhorn, B.; Reese, J.C.; Whitworth, R.J.; Stuart, J.J.; Chen, M.-S. Hessian Fly-Associated Bacteria: Transmission, Essentiality, and Composition. PLoS ONE 2011, 6, e23170. [Google Scholar] [CrossRef] [Green Version]
- Johnson, A.J.; Schemerhorn, B.J.; Shukle, R.H. A First Assessment of Mitochondrial DNA Variation and Geographic Distribution of Haplotypes in Hessian fly (Diptera: Cecidomyiidae). Ann. Entomol. Soc. Am. 2004, 97, 940–948. [Google Scholar] [CrossRef]
- Augustinos, A.A.; Santos-Garcia, D.; Dionyssopoulou, E.; Moreira, M.; Papapanagiotou, A.; Scarvelakis, M.; Doudoumis, V.; Ramos, S.; Aguiar, A.F.; Borges, P.A.V.; et al. Detection and Characterization of Wolbachia Infections in Natural Populations of Aphids: Is the Hidden Diversity Fully Unraveled? PLoS ONE 2011, 6, e28695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartley, L.J.; Bowen, H. PEG precipitation for selective removal of small DNA fragments. Focus 1996, 18, 27. [Google Scholar]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, Interactive, Scalable, and Extensible Microbiome Data Science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Edgar, R.C. UNCROSS2: Identification of cross-talk in 16S rRNA OTU tables. BioRxiv 2018, 400762. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Chen, J.; Bittinger, K.; Charlson, E.S.; Hoffmann, C.; Lewis, J.; Wu, G.D.; Collman, R.G.; Bushman, F.D.; Li, H. Associating microbiome composition with environmental covariates using generalized UniFrac distances. Bioinformatics 2012, 28, 2106–2113. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefort, V.; Longueville, J.-E.; Gascuel, O. SMS: Smart Model Selection in PhyML. Mol. Biol. Evol. 2017, 34, 2422–2424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akaike, H. Autoregressive model fitting for control. Ann. Inst. Stat. Math. 1971, 23, 163–180. [Google Scholar] [CrossRef]
- Comar, M.; D’Accolti, M.; Cason, C.; Soffritti, I.; Campisciano, G.; Lanzoni, L.; Bisi, M.; Volta, A.; Mazzacane, S.; Caselli, E. Introduction of NGS in Environmental Surveillance for Healthcare-Associated Infection Control. Microorganisms 2019, 7, 708. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.; Fanning, S.; Proos, S.; Jordan, K.; Srikumar, S. A Review on the Applications of Next Generation Sequencing Technologies as Applied to Food-Related Microbiome Studies. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef]
- Johnson, A.J.; Morton, P.K.; Schemerhorn, B.J.; Shukle, R.H. Use of a Nuclear Marker to Assess Population Structure in Hessian Fly (Diptera: Cecidomyiidae). Ann. Entomol. Soc. Am. 2011, 104, 666–674. [Google Scholar] [CrossRef]
- Behura, S.K.; Sahu, S.C.; Mohan, M.; Nair, S. Wolbachia in the Asian rice gall midge, Orseolia oryzae (Wood-Mason): Correlation between host mitotypes and infection status. Insect Mol. Biol. 2001, 10, 163–171. [Google Scholar] [CrossRef]
- KimMi, G.; HyunWoo, O.; HeeMoon, P.; HoYong, P. Molecular identification of Wolbachia naturally infected in Thecodiplosis japonensis (Diptera: Cecidominideii). Korean J. Entomol. 2000, 30, 139–146. [Google Scholar]
- Zytynska, S.E.; Weisser, W.W. The natural occurrence of secondary bacterial symbionts in aphids. Ecol. Entomol. 2016, 41, 13–26. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Shen, Z.-R.; Song, Y.; Liu, H.-Y.; Li, Z.-X. Distribution and diversity of Wolbachia in different populations of the wheat aphid Sitobion miscanthi (Hemiptera: Aphididae) in China. EJE 2009, 106, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Asimakis, E.D.; Doudoumis, V.; Hadapad, A.B.; Hire, R.S.; Batargias, C.; Niu, C.; Khan, M.; Bourtzis, K.; Tsiamis, G. Detection and characterization of bacterial endosymbionts in Southeast Asian tephritid fruit fly populations. BMC Microbiol. 2019, 19, 290. [Google Scholar] [CrossRef] [PubMed]
- Mateos, M.; Martinez, H.; Lanzavecchia, S.B.; Conte, C.; Guillén, K.; Morán-Aceves, B.M.; Toledo, J.; Liedo, P.; Asimakis, E.D.; Doudoumis, V.; et al. Wolbachia pipientis associated to tephritid fruit fly pests: From basic research to applications. BioRxiv 2018, 358333. [Google Scholar] [CrossRef] [Green Version]
- Yong, H.-S.; Song, S.-L.; Chua, K.-O.; Lim, P.-E. Predominance of Wolbachia endosymbiont in the microbiota across life stages of Bactrocera latifrons (Insecta: Tephritidae). Meta Gene 2017, 14, 6–11. [Google Scholar] [CrossRef]
- Schuler, H.; Arthofer, W.; Riegler, M.; Bertheau, C.; Krumböck, S.; Köppler, K.; Vogt, H.; Teixeira, L.A.; Stauffer, C. Multiple Wolbachia infections in Rhagoletis pomonella. Entomol. Exp. Appl. 2011, 139, 138–144. [Google Scholar] [CrossRef]
- Sarakatsanou, A.; Diamantidis, A.D.; Papanastasiou, S.A.; Bourtzis, K.; Papadopoulos, N.T. Effects of Wolbachia on fitness of the Mediterranean fruit fly (Diptera: Tephritidae). J. Appl. Entomol. 2011, 135, 554–563. [Google Scholar] [CrossRef]
- Sun, X.; Cui, L.; Li, Z. Diversity and phylogeny of Wolbachia infecting Bactrocera dorsalis (Diptera: Tephritidae) populations from China. Environ. Entomol. 2007, 36, 1283–1289. [Google Scholar] [CrossRef]
- Richardson, K.M.; Schiffer, M.; Griffin, P.C.; Lee, S.F.; Hoffmann, A.A. Tropical Drosophila pandora carry Wolbachia infections causing cytoplasmic incompatibility or male killing. Evolution 2016, 70, 1791–1802. [Google Scholar] [CrossRef]
- Hamm, C.A.; Begun, D.J.; Vo, A.; Smith, C.C.R.; Saelao, P.; Shaver, A.O.; Jaenike, J.; Turelli, M. Wolbachia do not live by reproductive manipulation alone: Infection polymorphism in Drosophila suzukii and D. subpulchrella. Mol. Ecol. 2014, 23, 4871–4885. [Google Scholar] [CrossRef] [Green Version]
- Mateos, M.; Castrezana, S.J.; Nankivell, B.J.; Estes, A.M.; Markow, T.A.; Moran, N.A. Heritable Endosymbionts of Drosophila. Genetics 2006, 174, 363–376. [Google Scholar] [CrossRef] [Green Version]
- Bourtzis, K.; Nirgianaki, A.; Markakis, G.; Savakis, C. Wolbachia Infection and Cytoplasmic Incompatibility in Drosophila Species. Genetics 1996, 144, 1063–1073. [Google Scholar] [PubMed]
- Moreira, M.; Aguiar, A.M.F.; Bourtzis, K.; Latorre, A.; Khadem, M. Wolbachia (Alphaproteobacteria: Rickettsiales) Infections in Isolated Aphid Populations from Oceanic Islands of the Azores Archipelago: Revisiting the Supergroups M and N. Environ. Entomol. 2019, 48, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Gerth, M. Classification of Wolbachia (Alphaproteobacteria, Rickettsiales): No evidence for a distinct supergroup in cave spiders. Infect. Genet. Evol. 2016, 43, 378–380. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-H.; Jia, L.-Y.; Xiao, J.-H.; Huang, D.-W. Discovery of a new Wolbachia supergroup in cave spider species and the lateral transfer of phage WO among distant hosts. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2016, 41, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Glowska, E.; Dragun-Damian, A.; Dabert, M.; Gerth, M. New Wolbachia supergroups detected in quill mites (Acari: Syringophilidae). Infect. Genet. Evol. 2015, 30, 140–146. [Google Scholar] [CrossRef]
- Ros, V.I.D.; Fleming, V.M.; Feil, E.J.; Breeuwer, J.A.J. How Diverse Is the Genus Wolbachia? Multiple-Gene Sequencing Reveals a Putatively New Wolbachia Supergroup Recovered from Spider Mites (Acari: Tetranychidae). Appl. Environ. Microbiol. 2009, 75, 1036–1043. [Google Scholar] [CrossRef] [Green Version]
- Baldo, L.; Hotopp, J.C.D.; Jolley, K.A.; Bordenstein, S.R.; Biber, S.A.; Choudhury, R.R.; Hayashi, C.; Maiden, M.C.J.; Tettelin, H.; Werren, J.H. Multilocus Sequence Typing System for the Endosymbiont Wolbachia pipientis. Appl. Environ. Microbiol. 2006, 72, 7098–7110. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Rousset, F.; O’Neil, S. Phylogeny and PCR-based classification of Wolbachia strains using wsp gene sequences. Proc. R. Soc. B Biol. Sci. 1998, 265, 509–515. [Google Scholar] [CrossRef]
- De Miguel, T.; Zhu, O.; Villa, T.G. Horizontal Gene Transfer Between Wolbachia and Animals. In Horizontal Gene Transfer: Breaking Borders Between Living Kingdoms; Villa, T.G., Viñas, M., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 227–234. ISBN 978-3-030-21862-1. [Google Scholar]
- Husnik, F.; McCutcheon, J.P. Functional horizontal gene transfer from bacteria to eukaryotes. Nat. Rev. Microbiol. 2018, 16, 67. [Google Scholar] [CrossRef]
- Doudoumis, V.; Tsiamis, G.; Wamwiri, F.; Brelsfoard, C.; Alam, U.; Aksoy, E.; Dalaperas, S.; Abd-Alla, A.; Ouma, J.; Takac, P.; et al. Detection and characterization of Wolbachia infections in laboratory and natural populations of different species of tsetse flies (genus Glossina). BMC Microbiol. 2012, 12, S3. [Google Scholar] [CrossRef] [Green Version]
- Hotopp, J.C.D. Horizontal gene transfer between bacteria and animals. Trends Genet. 2011, 27, 157–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, N.; Nikoh, N.; Ijichi, N.; Shimada, M.; Fukatsu, T. Genome fragment of Wolbachia endosymbiont transferred to X chromosome of host insect. Proc. Natl. Acad. Sci. USA 2002, 99, 14280–14285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordenstein, S.R.; Bordenstein, S.R. Temperature Affects the Tripartite Interactions between Bacteriophage WO, Wolbachia, and Cytoplasmic Incompatibility. PLoS ONE 2011, 6, e29106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmed, M.Z.; Barro, P.J.D.; Ren, S.-X.; Greeff, J.M.; Qiu, B.-L. Evidence for Horizontal Transmission of Secondary Endosymbionts in the Bemisia tabaci Cryptic Species Complex. PLoS ONE 2013, 8, e53084. [Google Scholar] [CrossRef] [PubMed]
- Huigens, M.E.; de Almeida, R.P.; Boons, P.A.H.; Luck, R.F.; Stouthamer, R. Natural interspecific and intraspecific horizontal transfer of parthenogenesis-inducing Wolbachia in Trichogramma wasps. Proc. Biol. Sci. 2004, 271, 509–515. [Google Scholar] [CrossRef] [Green Version]
- Li, S.-J.; Ahmed, M.Z.; Lv, N.; Shi, P.-Q.; Wang, X.-M.; Huang, J.-L.; Qiu, B.-L. Plantmediated horizontal transmission of Wolbachia between whiteflies. ISME J. 2017, 11, 1019–1028. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, M.Z.; Li, S.-J.; Xue, X.; Yin, X.-J.; Ren, S.-X.; Jiggins, F.M.; Greeff, J.M.; Qiu, B.-L. The Intracellular Bacterium Wolbachia Uses Parasitoid Wasps as Phoretic Vectors for Efficient Horizontal Transmission. PLoS Pathog. 2015, 11, e1004672. [Google Scholar] [CrossRef] [Green Version]
- Kubota, M.; Morii, T.; Miura, K. In vitro cultivation of parthenogenesis-inducing Wolbachia in an Aedes albopictus cell line. Entomol. Exp. Appl. 2005, 117, 83–87. [Google Scholar] [CrossRef]
- Calvitti, M.; Moretti, R.; Lampazzi, E.; Bellini, R.; Dobson, S.L. Characterization of a New Aedes albopictus (Diptera: Culicidae)—Wolbachia pipientis (Rickettsiales: Rickettsiaceae) Symbiotic Association Generated by Artificial Transfer of the w Pip Strain From Culex pipiens (Diptera: Culicidae). J. Med. Entomol. 2010, 47, 179–187. [Google Scholar] [CrossRef]
- Bian, G.; Joshi, D.; Dong, Y.; Lu, P.; Zhou, G.; Pan, X.; Xu, Y.; Dimopoulos, G.; Xi, Z. Wolbachia Invades Anopheles stephensi Populations and Induces Refractoriness to Plasmodium Infection. Science 2013, 340, 748–751. [Google Scholar] [CrossRef]
- Andrews, E.S.; Crain, P.R.; Fu, Y.; Howe, D.K.; Dobson, S.L. Reactive Oxygen Species Production and Brugia pahangi Survivorship in Aedes polynesiensis with Artificial Wolbachia Infection Types. PLoS Pathog. 2012, 8, e1003075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMeniman, C.J.; Lane, R.V.; Cass, B.N.; Fong, A.W.C.; Sidhu, M.; Wang, Y.-F.; O’Neill, S.L. Stable Introduction of a Life-Shortening Wolbachia Infection into the Mosquito Aedes aegypti. Science 2009, 323, 141–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, A.A.; Montgomery, B.L.; Popovici, J.; Iturbe-Ormaetxe, I.; Johnson, P.H.; Muzzi, F.; Greenfield, M.; Durkan, M.; Leong, Y.S.; Dong, Y.; et al. Successful establishment of Wolbachia in Aedes populations to suppress dengue transmission. Nature 2011, 476, 454–457. [Google Scholar] [CrossRef] [PubMed]
- Zabalou, S.; Riegler, M.; Theodorakopoulou, M.; Stauffer, C.; Savakis, C.; Bourtzis, K. Wolbachia-induced cytoplasmic incompatibility as a means for insect pest population control. Proc. Natl. Acad. Sci. USA 2004, 101, 15042–15045. [Google Scholar] [CrossRef] [Green Version]
- Apostolaki, A.; Livadaras, I.; Saridaki, A.; Chrysargyris, A.; Savakis, C.; Bourtzis, K. Transinfection of the olive fruit fly Bactrocera oleae with Wolbachia: Towards a symbiont-based population control strategy. J. Appl. Entomol. 2011, 135, 546–553. [Google Scholar] [CrossRef]
- Douglas, A.E. Multiorganismal insects: Diversity and function of resident microorganisms. Ann. Rev. Entomol. 2015, 60, 17–34. [Google Scholar] [CrossRef] [Green Version]
- Bansal, R.; Hulbert, S.H.; Reese, J.C.; Whitworth, R.J.; Stuart, J.J.; Chen, M.-S. Pyrosequencing Reveals the Predominance of Pseudomonadaceae in Gut Microbiome of a Gall Midge. Pathogens 2014, 3, 459–472. [Google Scholar] [CrossRef] [Green Version]
- Jones, R.T.; Sanchez, L.G.; Fierer, N. A Cross-Taxon Analysis of Insect-Associated Bacterial Diversity. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Kikuchi, Y. Endosymbiotic Bacteria in Insects: Their Diversity and Culturability. Microbes Environ. 2009, 24, 195–204. [Google Scholar] [CrossRef] [Green Version]
- Toju, H.; Fukatsu, T. Diversity and infection prevalence of endosymbionts in natural populations of the chestnut weevil: Relevance of local climate and host plants. Mol. Ecol. 2011, 20, 853–868. [Google Scholar] [CrossRef]
- Heinen, R.; Biere, A.; Harvey, J.A.; Bezemer, T.M. Effects of Soil Organisms on Aboveground Plant-Insect Interactions in the Field: Patterns, Mechanisms and the Role of Methodology. Front. Ecol. Evol. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Pineda, A.; Zheng, S.-J.; Van Loon, J.J.; Pieterse, C.M.; Dicke, M. Helping plants to deal with insects: The role of beneficial soil-borne microbes. Trends Plant Sci. 2010, 15, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Dittmer, J.; Bouchon, D. Feminizing Wolbachia influence microbiota composition in the terrestrial isopod Armadillidium vulgare. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Simhadri, R.K.; Fast, E.M.; Guo, R.; Schultz, M.J.; Vaisman, N.; Ortiz, L.; Bybee, J.; Slatko, B.E.; Frydman, H.M. The Gut Commensal Microbiome of Drosophila melanogaster Is Modified by the Endosymbiont Wolbachia. mSphere 2017, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Region | Location | Coordinates | Number of Insects | |||
|---|---|---|---|---|---|---|
| Altitude | Latitude | Longitude | Female | Male | ||
| Safi | Jamaat Shaim | 173 | 32.24076 | −8.46976 | 32 | 27 |
| Fes | Fes | 584 | 34.01436 | −5.34543 | 23 | 30 |
| Doukkala | Khemis Zemamra | 162 | 32.614952 | −8.664802 | 19 | 8 |
| Chaouia | Sidi El Aidi | 247 | 33.07341 | −7.37935 | 16 | 9 |
| - | Laboratory colony | - | - | - | 40 | 40 |
| Population | Gender | Sample Size | Wolbachia | Spiroplasma | Cardinium | Arsenophonus |
|---|---|---|---|---|---|---|
| Doukkala | Female | 19 | + (3) | − | − | − |
| Male | 8 | + (2) | − | − | − | |
| Safi | Female | 32 | − | − | − | − |
| Male | 27 | − | − | − | − | |
| Fes | Female | 23 | − | − | − | − |
| Male | 30 | − | − | − | − | |
| Chaouia | Female | 16 | − | − | − | − |
| Male | 9 | − | − | − | − | |
| Laboratory | Female | 40 | − | − | − | − |
| Male | 40 | + (8) | ||||
| Total | 244 | + (13) | − | − | − |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bel Mokhtar, N.; Maurady, A.; Britel, M.R.; El Bouhssini, M.; Batargias, C.; Stathopoulou, P.; Asimakis, E.; Tsiamis, G. Detection of Wolbachia Infections in Natural and Laboratory Populations of the Moroccan Hessian Fly, Mayetiola destructor (Say). Insects 2020, 11, 340. https://doi.org/10.3390/insects11060340
Bel Mokhtar N, Maurady A, Britel MR, El Bouhssini M, Batargias C, Stathopoulou P, Asimakis E, Tsiamis G. Detection of Wolbachia Infections in Natural and Laboratory Populations of the Moroccan Hessian Fly, Mayetiola destructor (Say). Insects. 2020; 11(6):340. https://doi.org/10.3390/insects11060340
Chicago/Turabian StyleBel Mokhtar, Naima, Amal Maurady, Mohammed Reda Britel, Mustapha El Bouhssini, Costas Batargias, Panagiota Stathopoulou, Elias Asimakis, and George Tsiamis. 2020. "Detection of Wolbachia Infections in Natural and Laboratory Populations of the Moroccan Hessian Fly, Mayetiola destructor (Say)" Insects 11, no. 6: 340. https://doi.org/10.3390/insects11060340
APA StyleBel Mokhtar, N., Maurady, A., Britel, M. R., El Bouhssini, M., Batargias, C., Stathopoulou, P., Asimakis, E., & Tsiamis, G. (2020). Detection of Wolbachia Infections in Natural and Laboratory Populations of the Moroccan Hessian Fly, Mayetiola destructor (Say). Insects, 11(6), 340. https://doi.org/10.3390/insects11060340