Pan Traps for Tracking Honey Bee Activity-Density: A Case Study in Soybeans


Abstract
:1. Introduction
2. Methods
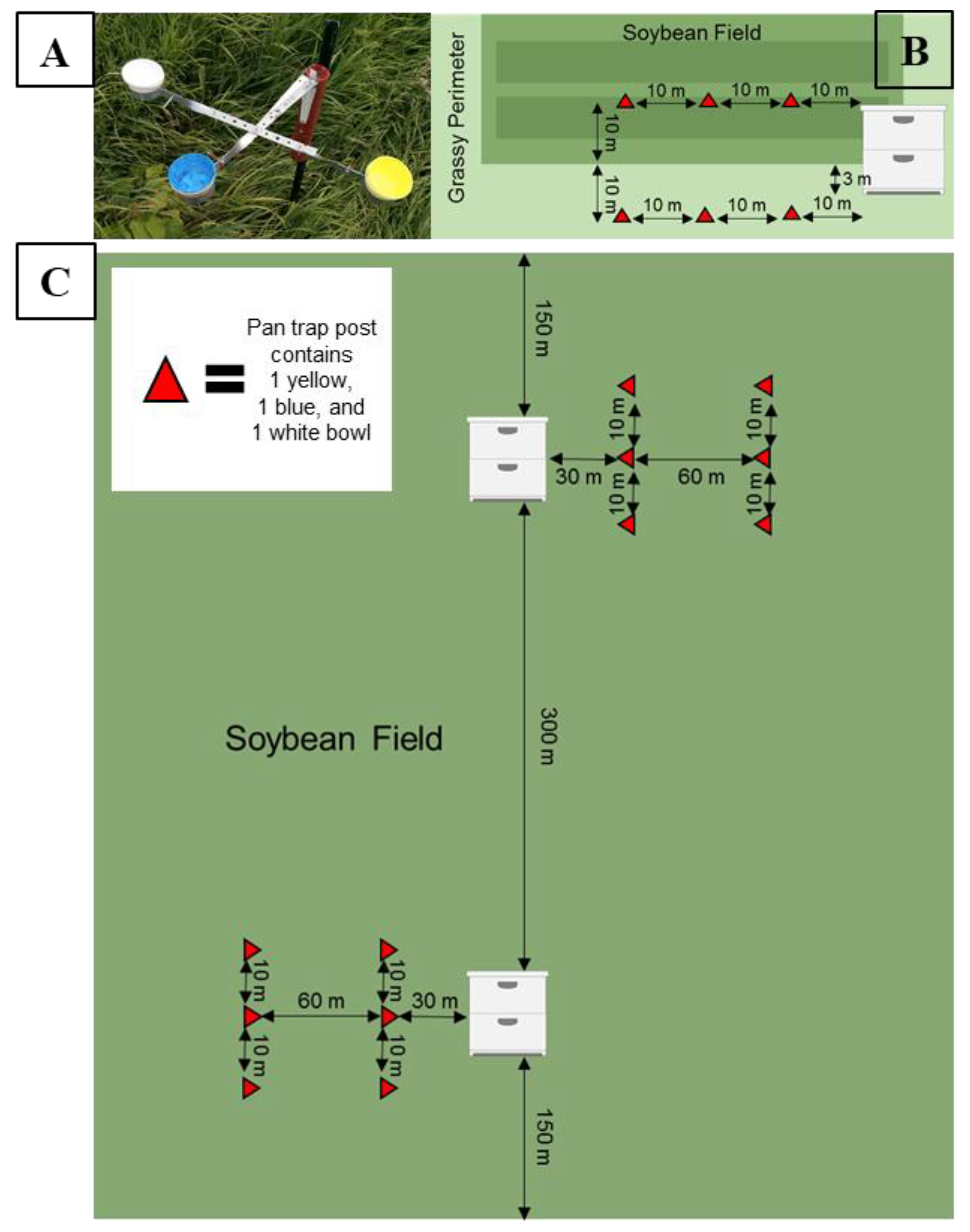
2.1. Experiment One: Does Honey Bee Activity-Density Vary with Trap Color, the Presence of an Apiary, Bee Population within an Apiary, Surrounding Landscape Composition, Trap Placement, and Soybean Phenology?
2.2. Site Selection
2.3. Honey Bee Apiary Placement
2.4. Estimating Honey Bee Activity-Density
2.5. Statistical Analysis
2.6. Experiment Two: Does Honey Bee Activity-Density Vary with Distance from Colonies?
2.7. Site Selection
2.8. Honey Bee Apiary Placement
2.9. Estimating Honey Bee Activity-Density
2.10. Assessing Honey Bee Wing Wear
2.11. Statistical Analyses
2.12. Combined Analysis: Does Honey Bee Activity-Density in Soybeans Vary with the Number of Colonies Present?
3. Results
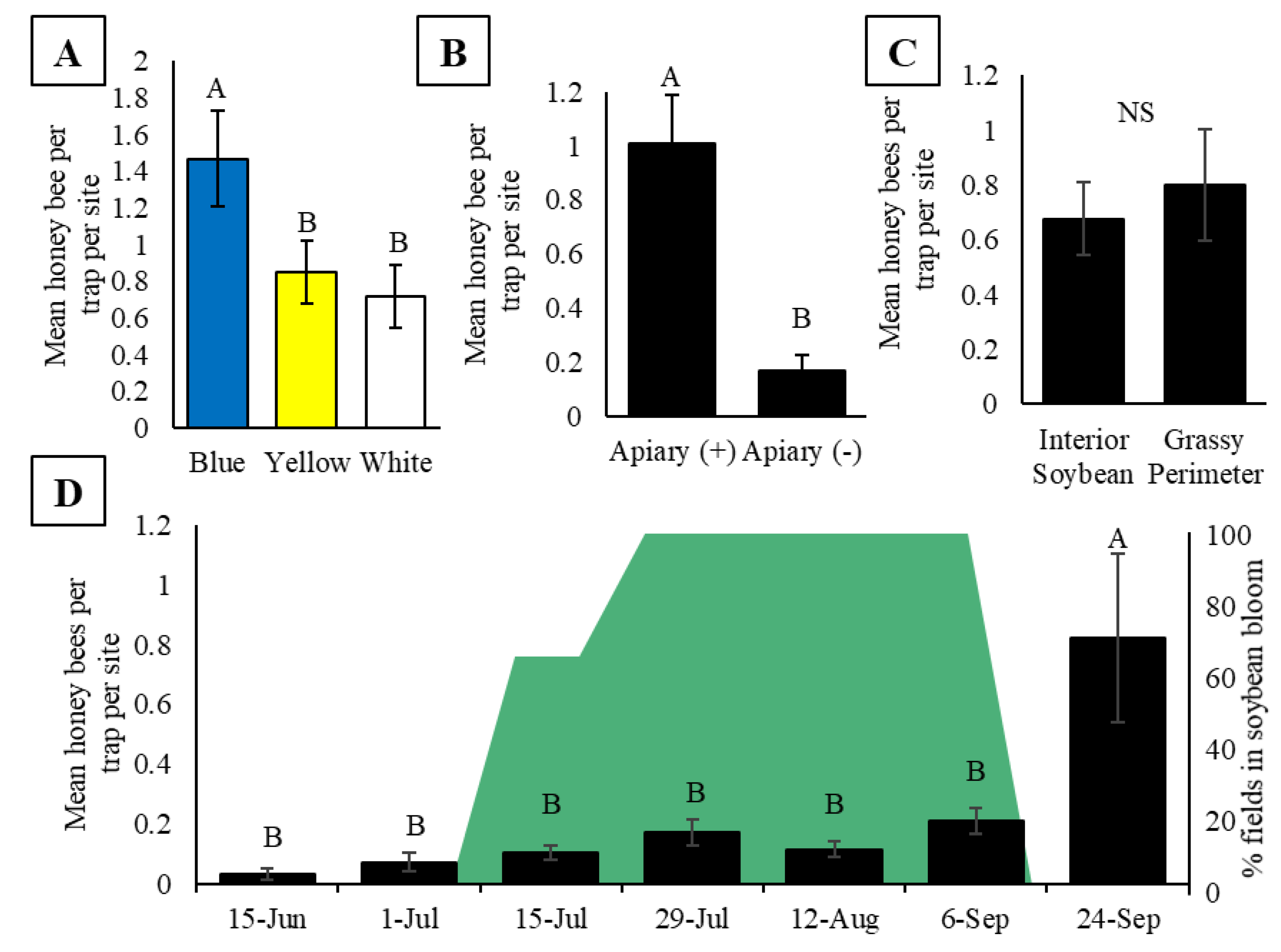
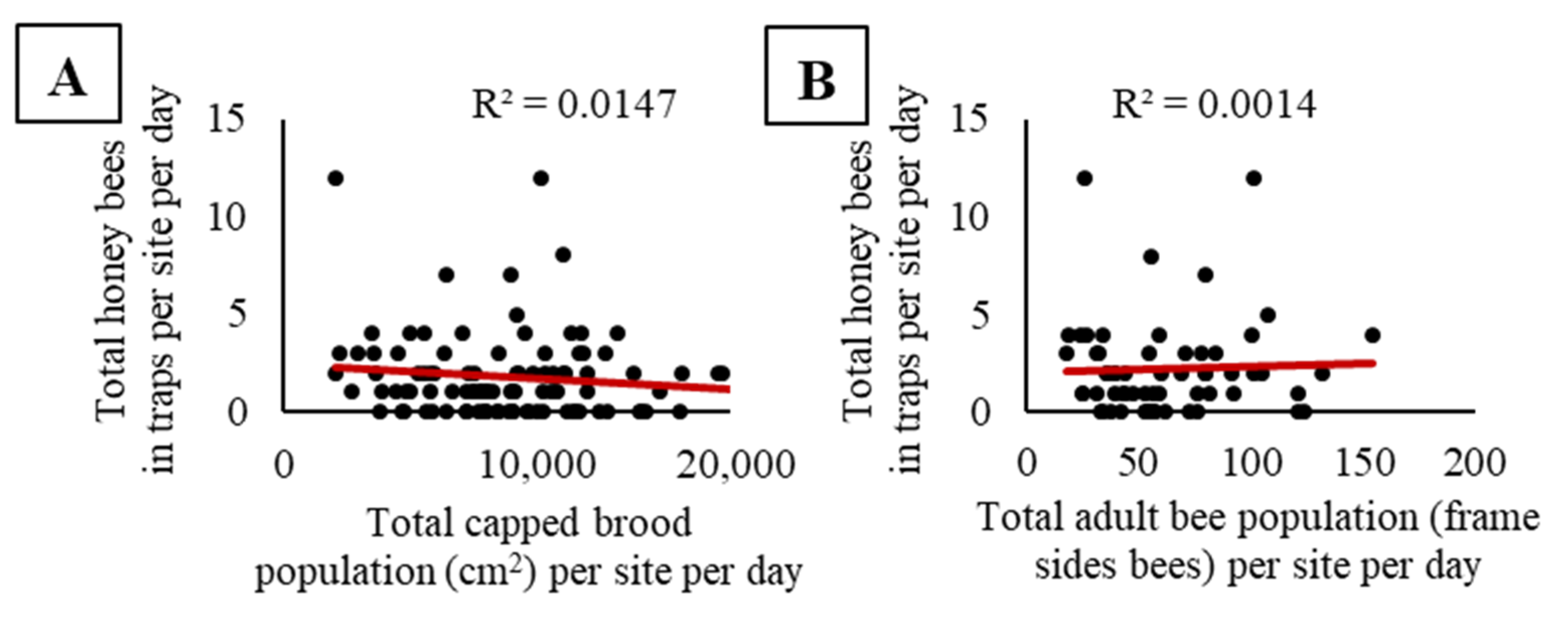
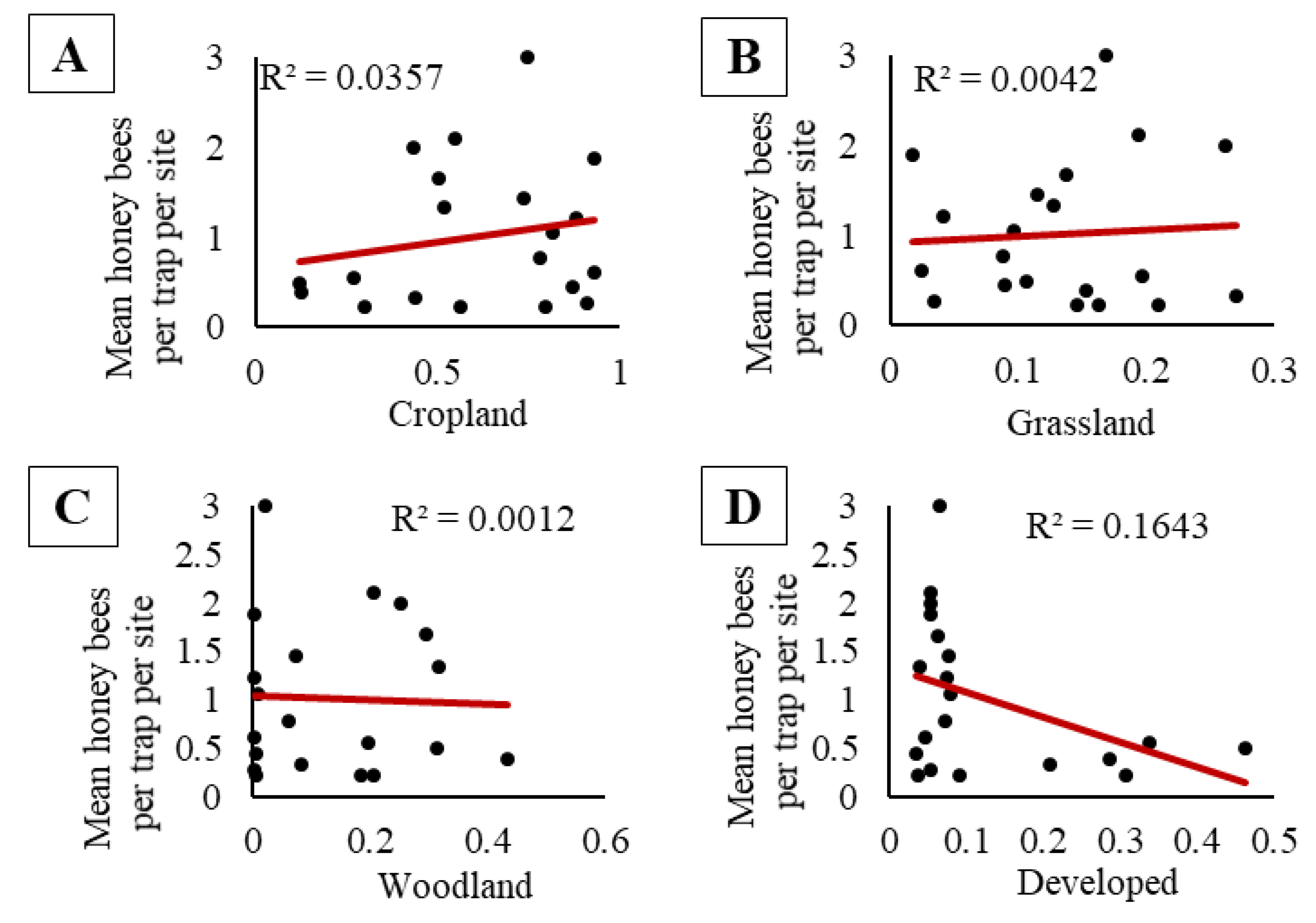
3.1. Experiment One: Does Honey Bee Activity-Density Vary with Trap Color, the Presence of an Apiary, Bee Population within an Apiary, Surrounding Landscape Composition, Trap Placement, and Soybean Phenology?
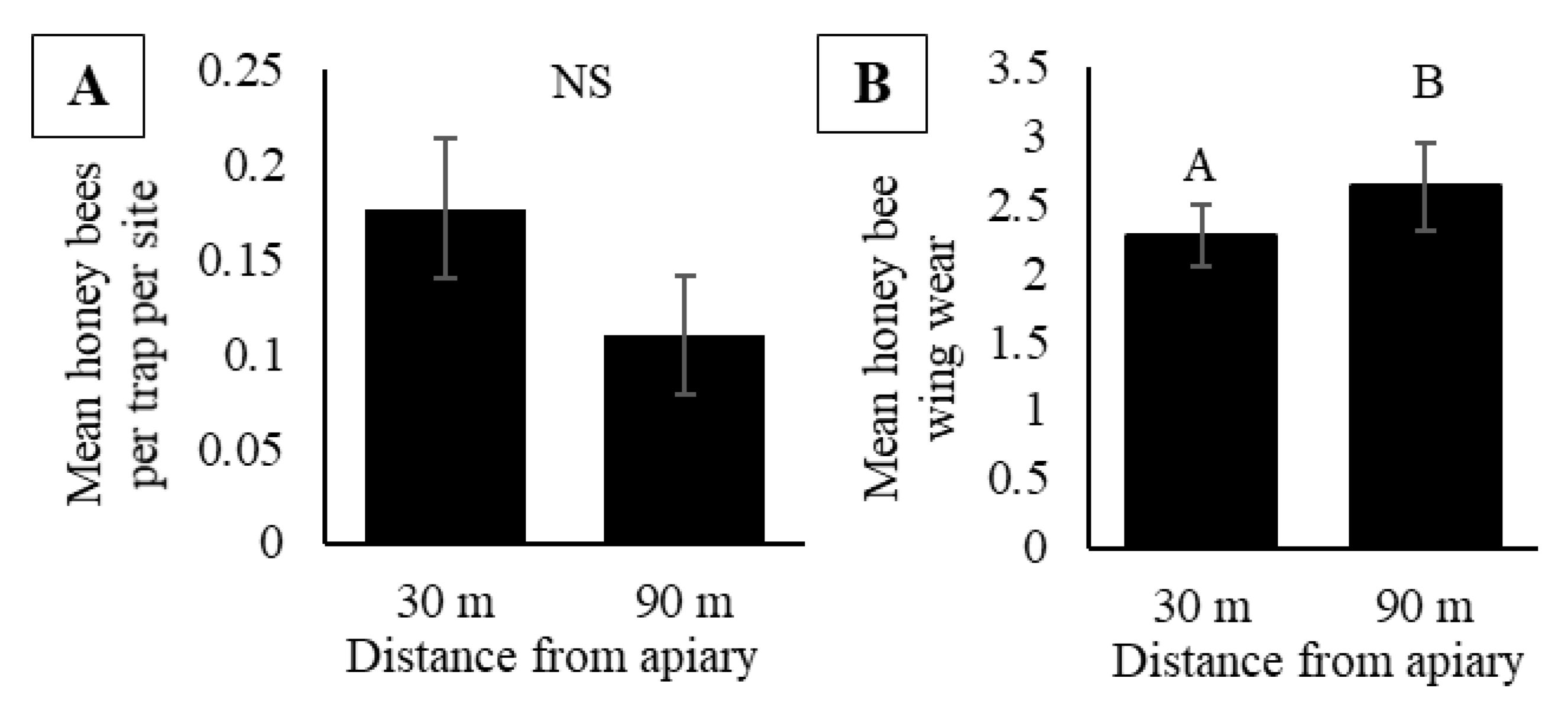
3.2. Experiment Two: Does Honey Bee Activity-Density Vary with Distance from Colonies and by Site Type?
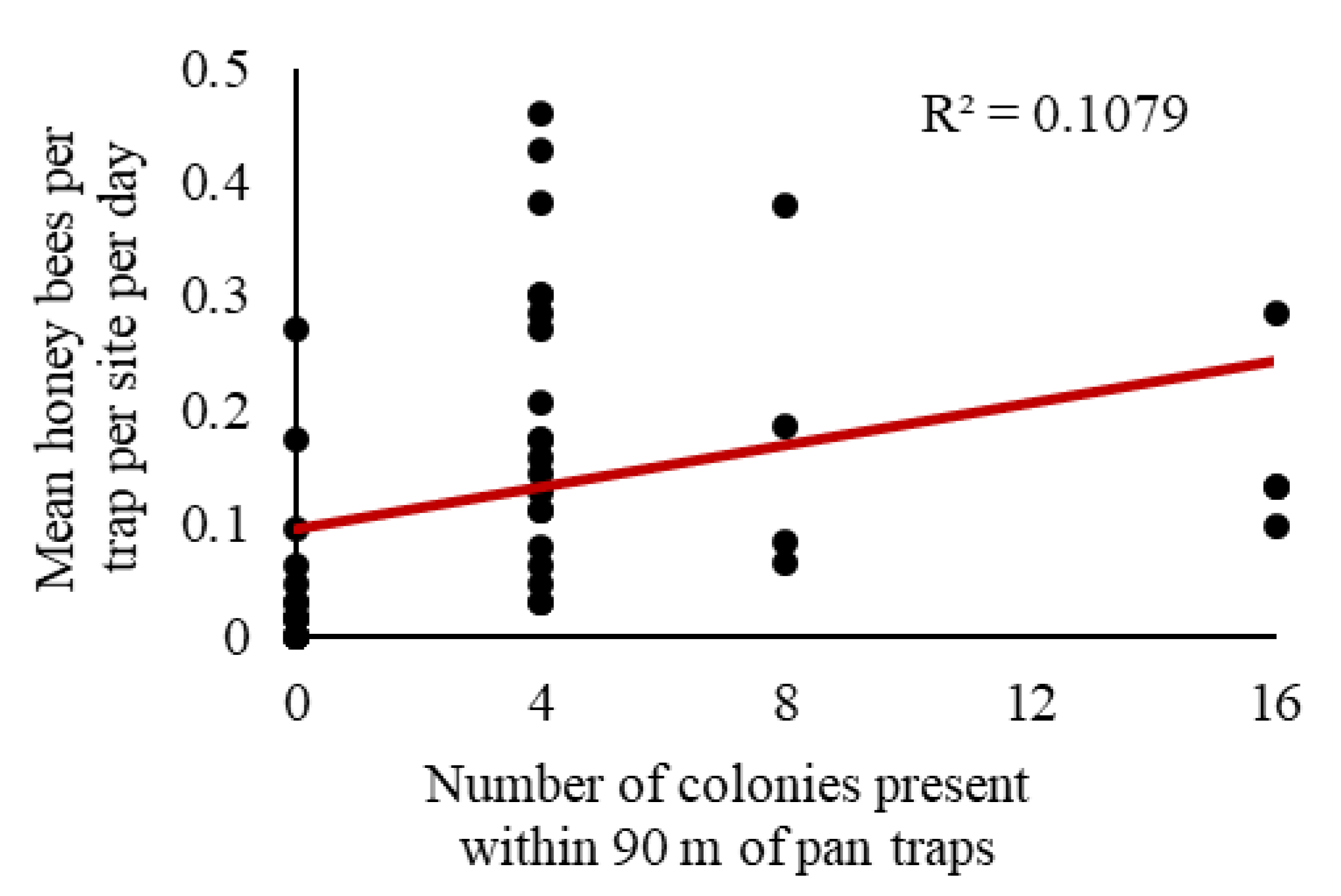
3.3. Combined Analysis: Does Honey Bee Activity-Density in Soybeans Vary with the Number of Colonies Present?
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability
References
- Bąkowski, M.; Piekarska-Boniecka, H.; Dolańska-Niedbała, E. Monitoring of the red-belted clearwing moth, Synanthedon myopaeformis, and its parasitoid Liotryphon crassiseta in apple orchards in yellow Moericke traps. J. Insect Sci. 2013, 13, 4. [Google Scholar] [CrossRef] [Green Version]
- Mitra, A.; Majid, F.; Toshko, L. A contribution to the fauna of Crabronidae (Hymenoptera, Apoidea) in South-Western Iran. J. Insect Biodivers. 2015, 3, 1–30. [Google Scholar] [CrossRef] [Green Version]
- Skvarla, M.J.; Dowling, A.P.G. A comparison of trapping techniques (Coleoptera: Carabidae, Buprestidae, Cerambycidae, and Curculionoidea excluding Scolytinae). J. Insect Sci. 2017, 17, 1–28. [Google Scholar] [CrossRef]
- Droege, S. Impact of Color and Size of Bowl Trap on Numbers of Bees Captured. Available online: http://online.sfsu.edu/~beeplot/pdfs/color%20and%20size.pdf (accessed on 12 December 2019).
- Droege, S.; Tepedino, V.; Lebuhn, G.; Link, W.; Minckley, R.; Chen, Q.; Conrad, C. Spatial patterns of bee captures in North American bowl trapping surveys. Insect Conserv. Divers. 2010, 3, 15–23. [Google Scholar] [CrossRef]
- Mccravy, K.W.; Ruholl, J.D. Bee (Hymenoptera: Apoidea) Diversity and Sampling Methodology in a Midwestern USA Deciduous Forest. Insects 2017, 8, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prado, S.; Ngo, H.; Florez, J.; Collazo, J. Sampling bees in tropical forests and agroecosystems: A review. J. Insect Conserv. 2017, 21, 753–770. [Google Scholar] [CrossRef]
- Cane, J.H.; Minckley, R.L.; Kervin, L.J. Sampling Bees (Hymenoptera: Apiformes) for Pollinator Community Studies: Pitfalls of Pan-Trapping. J. Kans. Entomol. Soc. 2000, 73, 225–231. [Google Scholar]
- Roulston, T.A.H.; Smith, S.A.; Brewster, A.L. A Comparison of Pan Trap and Intensive Net Sampling Techniques for Documenting a Bee (Hymenoptera: Apiformes) Fauna. J. Kans. Èntomol. Soc. 2007, 80, 179–181. [Google Scholar] [CrossRef]
- Wilson, J.S.; Griswold, T.; Messinger, O.J. Sampling Bee Communities (Hymenoptera: Apiformes) in a Desert Landscape: Are Pan Traps Sufficient? J. Kans. Èntomol. Soc. 2008, 81, 288–300. [Google Scholar] [CrossRef]
- Portman, Z.M.; Bruninga-Socolar, B.; Cariveau, D.P. The State of Bee Monitoring in the United States: A Call to Refocus Away From Bowl Traps and Towards More Effective Methods. Ann. Èntomol. Soc. Am. 2020, 1–6. [Google Scholar] [CrossRef]
- Parys, K.A.; Esquivel, I.L.; Wright, K.W.; Griswold, T.; Brewer, M.J. Native Pollinators (Hymenoptera: Anthophila) in Cotton Grown in the Gulf South, United States. Agronomy 2020, 10, 698. [Google Scholar] [CrossRef]
- Wood, T.; Holland, J.; Goulson, D. A comparison of techniques for assessing farmland bumblebee populations. Oecologia 2015, 177, 1093–1102. [Google Scholar] [CrossRef] [PubMed]
- Prendergast, K.S.; Menz, M.H.M.; Dixon, K.W.; Bateman, P.W. The relative performance of sampling methods for native bees: An empirical test and review of the literature. Ecosphere 2020, 11, 1–22. [Google Scholar] [CrossRef]
- Toler, T.R.; Evans, E.W.; Tepedino, V.J. Pan-trapping for bees (Hymenoptera: Apiformes) in Utah’s West Desert: The importance of color diversity. Pan Pac. Entomol. 2005, 81, 103–113. [Google Scholar]
- Droege, S.; Engler, J.; Sellers, E.; O’Brien, L. National Protocol Framework for the Inventory and Monitoring of Bees; U.S. Fish and Wildlife Service: Fort Collins, CO, USA, 2016; pp. 1–89.
- Grundel, R.; Frohnapple, K.J.; Jean, R.P.; Pavlovic, N.B. Effectiveness of Bowl Trapping and Netting for Inventory of a Bee Community. Environ. Èntomol. 2011, 40, 374–380. [Google Scholar] [CrossRef] [Green Version]
- Seeley, T.D. The Wisdom of the Hive: The Social Physiology of Honey Bee Colonies; Harvard University Press: Cambridge, MA, USA, 1995. [Google Scholar]
- Delaplane, K.S.; van Der Steen, J.; Guzman-Novoa, E. Standard methods for estimating strength parameters of Apis mellifera colonies. J. Apic. Res. 2013, 52, 1–12. [Google Scholar] [CrossRef]
- Kulhanek, K.; Steinhauer, N.; Rennich, K.; Caron, D.M.; Sagili, R.R.; Pettis, J.S.; Ellis, J.D.; Wilson, M.E.; Wilkes, J.T.; Tarpy, D.R.; et al. A national survey of managed honey bee 2015–2016 annual colony losses in the USA. J. Apic. Res. 2017, 56, 328–340. [Google Scholar] [CrossRef] [Green Version]
- Otto, C.R.V.; Zheng, H.; Gallant, A.L.; Iovanna, R.; Carlson, B.L.; Smart, M.D.; Hyberg, S. Past role and future outlook of the Conservation Reserve Program for supporting honey bees in the Great Plains. Proc. Natl. Acad. Sci. USA. 2018, 115, 7629. [Google Scholar] [CrossRef] [Green Version]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee declines driven by combined stress from parasites, pesticides, and lack of flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Sponsler, D.B.; Grozinger, C.M.; Hitaj, C.; Rundlöf, M.; Botías, C.; Code, A.; Lonsdorf, E.V.; Melathopoulos, A.P.; Smith, D.J.; Suryanarayanan, S.; et al. Pesticides and pollinators: A socioecological synthesis. Sci. Total. Environ. 2019, 662, 1012–1027. [Google Scholar] [CrossRef] [PubMed]
- Thapa, R.B. Honeybees and other Insect Pollinators of Cultivated Plants: A Review. J. Inst. Agric. Anim. Sci. 2006, 27, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Rucker, R.R.; Thurman, W.N.; Burgett, M. Honey Bee Pollination Markets and the Internalization of Reciprocal Benefits. Am. J. Agric. Econ. 2012, 94, 956–977. [Google Scholar] [CrossRef]
- Todd, F.E.; McGregor, S.E. The use of Honey Bees in the Production of Crops. Annu. Rev. Entomol. 1960, 5, 265–278. [Google Scholar] [CrossRef]
- Nicodemo, D.; Couto, R.H.N.; Malheiros, E.B.; De Jong, D. Honey bee as an effective pollinating agent of pumpkin. Sci. Agricola 2009, 66, 476–480. [Google Scholar] [CrossRef] [Green Version]
- Artz, D.R.; Nault, B.A. Performance of Apis mellifera, Bombus impatiens, and Peponapis pruinosa (Hymenoptera: Apidae) as pollinators of pumpkin. J. Econ. Entomol. 2011, 104, 1153–1161. [Google Scholar] [CrossRef]
- Carr-Markell, M.K.; Demler, C.M.; Couvillon, M.J.; Schürch, R.; Spivak, M. Do honey bee (Apis mellifera) foragers recruit their nestmates to native forbs in reconstructed prairie habitats? PLoS ONE 2020, 15, e0228169. [Google Scholar] [CrossRef] [Green Version]
- Couvillon, M.J.; Schurch, R.; Ratnieks, F.L.W. Waggle dance distances as integrative indicators of seasonal foraging challenges. PLoS ONE 2014, 9, e93495. [Google Scholar] [CrossRef] [Green Version]
- Couvillon, M.; Riddell Pearce, F.; Accleton, C.; Fensome, K.; Quah, S.; Taylor, E.; Ratnieks, F. Honey bee foraging distance depends on month and forage type. Apidologie 2015, 46, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.W.; Tautz, J.; Grünewald, B.; Fuchs, S. RFID Tracking of Sublethal Effects of Two Neonicotinoid Insecticides on the Foraging Behavior of Apis mellifera (RFID Tracking of Sublethal Insecticide Effects). PLoS ONE 2012, 7, e30023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenczar, P.; Lutz, C.C.; Rao, V.D.; Goldenfeld, N.; Robinson, G.E. Automated monitoring reveals extreme interindividual variation and plasticity in honeybee foraging activity levels. Anim. Behav. 2014, 95, 41–48. [Google Scholar] [CrossRef]
- Yang, E.C.; Chuang, Y.; Chen, Y.; Chang, L.H. Abnormal Foraging Behavior Induced by Sublethal Dosage of Imidacloprid in the Honey Bee (Hymenoptera: Apidae). J. Econ. Entomol. 2008, 101, 1743–1748. [Google Scholar] [CrossRef]
- Dukku, H. Identification of plants visited by the honeybee, Apis mellifera L. in the Sudan Savanna zone of northeastern Nigeria. Afr. J. Plant Sci. 2013, 7, 273–284. [Google Scholar] [CrossRef] [Green Version]
- Baum, K.A.; Wallen, K.E. Potential Bias in Pan Trapping as a Function of Floral Abundance. J. Kans. Entomol. Soc. 2011, 84, 155–159. [Google Scholar] [CrossRef]
- Gonçalves, R.; Santos, E.; Scott-Santos, C. Bees (Hymenoptera: Apoidea: Apidae s.l.) captured with Malaise and pan traps along an altitudinal gradient in the Parque Estadual da Serra do Mar, Ubatuba, São Paulo, Brazil. Check List 2012, 8, 53–56. [Google Scholar] [CrossRef] [Green Version]
- Aizen, M.A.; Feinsinger, P. Habitat Fragmentation, Native Insect Pollinators, and Feral Honey Bees in Argentine ‘Chaco Serrano’. Ecol. Appl. 1994, 4, 378–392. [Google Scholar] [CrossRef]
- Baum, K.A.; Rubink, W.L.; Coulson, R.N. Trapping of feral honey bee workers (Hymenoptera: Apidae) in a coastal prairie landscape: Effects of season and vegetation type. Can. Entomol. 2006, 138, 228–234. [Google Scholar] [CrossRef]
- Westphal, C.; Bommarco, R.; Carré, G.; Lamborn, E.; Morison, N.; Petanidou, T.; Potts, S.G.; Roberts, S.P.M.; Szentgyörgyi, H.; Tscheulin, T.; et al. Measuring Bee Diversity in Different European Habitats and Biogeographical Regions. Ecol. Monogr. 2008, 78, 653–671. [Google Scholar] [CrossRef] [Green Version]
- Gill, K.A.; O’Neal, M.E. Survey of Soybean Insect Pollinators: Community Identification and Sampling Method Analysis. Environ. Entomol. 2015, 44, 488–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wheelock, M.J.; Rey, K.P.; O’Neal, M.E. Defining the Insect Pollinator Community Found in Iowa Corn and Soybean Fields: Implications for Pollinator Conservation. Environ. Entomol. 2016, 45, 1099–1106. [Google Scholar] [CrossRef] [PubMed]
- Wheelock, M.J.; O’Neal, M.E. Insect Pollinators in Iowa Cornfields: Community Identification and Trapping Method Analysis. PLoS ONE 2016, 11, e0143479. [Google Scholar] [CrossRef] [Green Version]
- Popic, T.J.; Davila, Y.C.; Wardle, G.M. Evaluation of common methods for sampling invertebrate pollinator assemblages: Net sampling out-perform pan traps. PLoS ONE 2013, 8, e66665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brosi, B.J.; Daily, G.C.; Ehrlich, P.R. Bee community shifts with landscape context in a tropical countryside. Ecol. Appl. 2007, 17, 418–430. [Google Scholar] [CrossRef]
- Hokkanen, H.; Holopainen, J.K. Carabid species and activity densities in biologically and conventionally managed cabbage fields. J. Appl. Entomol. 1986, 102, 353–363. [Google Scholar] [CrossRef]
- Nuttman, C.V.; Otieno, M.; Kwapong, P.K.; Combey, R.; Willmer, P.; Potts, S.G. The Utility of Aerial Pan-Trapping for Assessing Insect Pollinators Across Vertical Strata. J. Kans. Entomol. Soc. 2011, 84, 260–270. [Google Scholar] [CrossRef]
- NASS-USDA. State Agricultural Overview. Available online: https://www.nass.usda.gov/Statistics_by_State/Ag_Overview/ (accessed on 17 April 2020).
- Brown, P.W.; Schulte, L.A. Agricultural landscape change (1937–2002) in three townships in Iowa, USA. Landsc. Urban Plan. 2011, 100, 202–212. [Google Scholar] [CrossRef]
- Klopatek, J.M.; Olson, R.J.; Emerson, C.J.; Joness, J.L. Land-use conflicts with natural vegetation in the United States. Environ. Conserv. 2009, 6, 191–199. [Google Scholar] [CrossRef]
- Seeley, T.D. The nest of the honey bee (Apis mellifera). Insectes Sociaux 1976, 23, 495–512. [Google Scholar] [CrossRef]
- USDA. Attractiveness of Agricultural Crops to Pollinating Bees for the Collection of Nectar and/or Pollen. 2017. Available online: https://www.usda.gov/oce/opmp/Attractiveness%20of%20Agriculture%20Crops%20to%20Pollinating%20Bees%20Report-FINAL_Web%20Version_Jan%203_2018.pdf (accessed on 11 October 2019).
- Ellis, M.; Baxendale, F.P.; Keith, D.L. Protecting Bees When Using Insecticides; Cooperative Extension; Institute of Agriculture and Natural Resources-University of Nebraska-Lincoln: Lincoln, NE, USA, 1998. [Google Scholar]
- Westphal, C.; Steffan-Dewenter, I.; Tscharntke, T. Mass flowering crops enhance pollinator densities at a landscape scale. Ecol. Lett. 2003, 6, 961–965. [Google Scholar] [CrossRef]
- Dolezal, A.G.; St Clair, A.L.; Zhang, G.; Toth, A.L.; O’Neal, M.E. Native habitat mitigates feast-famine conditions faced by honey bees in an agricultural landscape. Proc. Natl. Acad. Sci. USA 2019, 116, 25147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiari, W.C.; Toledo, V.d.A.A.d.; Ruvolo-Takasusuki, M.C.C.; Oliveira, A.J.B.d.; Sakaguti, E.S.; Attencia, V.M.; Costa, F.M.; Mitsui, M.H. Pollination of soybean (Glycine max L. Merril) by honeybees (Apis mellifera L.). Braz. Arch. Boil. Technol. 2005, 48, 31–36. [Google Scholar] [CrossRef]
- Milfont, M.; Rocha, E.E.M.; Lima, A.O.N.; Freitas, B.M. Higher soybean production using honeybee and wild pollinators, a sustainable alternative to pesticides and autopollination. Environ. Chem. Lett. 2013, 11, 335. [Google Scholar] [CrossRef]
- Blettler, D.; FagÚNdez, G.; Caviglia, O. Contribution of honeybees to soybean yield. Apidologie 2018, 49, 101–111. [Google Scholar] [CrossRef] [Green Version]
- Colin, T.; Meikle, W.G.; Wu, X.; Barron, A.B. Traces of a Neonicotinoid Induce Precocious Foraging and Reduce Foraging Performance in Honey Bees. Environ. Sci. Technol. 2019, 53, 8252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tosi, S.; Burgio, G.; Nieh, J.C. A common neonicotinoid pesticide, thiamethoxam, impairs honey bee flight ability. Sci. Rep. 2017, 7, 1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pedersen, P. Soybean Growth and Development; Iowa State University, University Extension: Ames, IA, USA, 2004. [Google Scholar]
- Hodgson, E.W.; McCornack, B.P.; Tilmon, K.; Knodel, J.J. Management Recommendations for Soybean Aphid (Hemiptera: Aphididae) in the United States. J. Integr. Pest Manag. 2012, 3, E1–E10. [Google Scholar] [CrossRef]
- Samson-Robert, O.; Labrie, G.; Chagnon, M.; Fournier, V. Planting of neonicotinoid-coated corn raises honey bee mortality and sets back colony development. PeerJ 2017, 5, e3670. [Google Scholar] [CrossRef] [Green Version]
- Krupke, C.H.; Long, E.Y. Intersections between neonicotinoid seed treatments and honey bees. Curr. Opin. Insect Sci. 2015, 10, 8–13. [Google Scholar] [CrossRef]
- USDA-NASS. United State Department of Agriculture-National Agricultural Statisitcs Survey; Upper Midwest Region—Iowa Field Office 2015 Iowa Crop Progress Review. 2015. Available online: https://www.nass.usda.gov/Statistics_by_State/Iowa/Publications/Crop_Progress_&_Condition/2015/IA%20Annual%20Crop%20Progress%202015.pdf (accessed on 23 November 2019).
- USDA-NASS. United States Department of Agriculture-National Agricultural Statistics Survey: Upper Midwest Region—Iowa Field Office 2016 Iowa Crop Progress Review. 2016. Available online: https://www.nass.usda.gov/Statistics_by_State/Iowa/Publications/Crop_Progress_&_Condition/2016/IA_Annual_Crop_Progress_2016.pdf (accessed on 23 November 2019).
- Mueller, U.; Wolf-Mueller, B. A method for estimating the age of bees: Age-dependent wing wear and coloration in the Wool-Carder bee Anthidium manicatum (hymenoptera: Megachilidae). J. Insect Behav. 1993, 6, 529–537. [Google Scholar] [CrossRef]
- Adhikari, S.; Burkle, L.A.; O’Neill, K.M.; Weaver, D.K.; Delphia, C.M.; Menalled, F.D. Dryland Organic Farming Partially Offsets Negative Effects of Highly Simplified Agricultural Landscapes on Forbs, Bees, and Bee-Flower Networks. Environ. Entomol. 2019, 48, 826–835. [Google Scholar] [CrossRef]
- Giurfa, M.; Núñez, J.; Chittka, L.; Menzel, R. Colour preferences of flower-naive honeybees. J. Comp. Physiol. A 1995, 177, 247–259. [Google Scholar] [CrossRef]
- Lunau, K.; Maier, E. Innate colour preferences of flower visitors. J. Comp. Physiol. A 1995, 177, 1–19. [Google Scholar] [CrossRef]
- St. Clair, A.L.; Dolezal, A.G.; Zhang, G.; O’Neal, M.E.; Toth, A.L. Diversified farming in a monoculture landscape: Effects on honey bee health and wild bee communities. Environ. Entomol. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Stein, D.S.; Debinski, D.M.; Pleasants, J.M.; Toth, A.L. Evaluating Native Bee Communities and Nutrition in Managed Grasslands. Environ. Entomol. 2020, 1–9. [Google Scholar] [CrossRef]
- Zurbuchen, A.; Cheesman, S.; Klaiber, J.; Müller, A.; Hein, S.; Dorn, S. Long foraging distances impose high costs on offspring production in solitary bees. J. Anim. Ecol. 2010, 79, 674. [Google Scholar] [CrossRef]
- Zurbuchen, A.; Landert, L.; Klaiber, J.; Müller, A.; Hein, S.; Dorn, S. Maximum foraging ranges in solitary bees: Only few individuals have the capability to cover long foraging distances. Biol. Conserv. 2010, 143, 669–676. [Google Scholar] [CrossRef]
- Visscher, P.K.; Seeley, T.D. Foraging strategy of honeybee colonies in a temperate deciduous forest. Ecology 1982, 63, 1790–1801. [Google Scholar] [CrossRef]
- Capaldi, E.A.; Dyer, F.C. The role of orientation flights on homing performance in honeybees. J. Exp. Biol. 1999, 202, 1655–1666. [Google Scholar]
- Higginson, A.D.; Gilbert, F. Paying for nectar with wingbeats: A new model of honeybee foraging. Proc. Biol. Sci. 2004, 271, 2595–2603. [Google Scholar] [CrossRef] [Green Version]
- Higginson, A.D.; Barnard, C.J.; Tofilski, A.; Medina, L.; Ratnieks, F. Experimental Wing Damage Affects Foraging Effort and Foraging Distance in Honeybees. Psyche 2011, 2011, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Klein, S.; Pasquaretta, C.; He, X.J.; Perry, C.; Søvik, E.; Devaud, J.-M.; Barron, A.B.; Lihoreau, M. Honey bees increase their foraging performance and frequency of pollen trips through experience. Sci. Rep. 2019, 9, 6778. [Google Scholar] [CrossRef] [PubMed]
- Eckert, C.; Winston, M.; Ydenberg, R. The relationship between population size, amount of brood, and individual foraging behaviour in the honey bee, Apis mellifera L. Oecologia 1994, 97, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Traynor, K.S.; Le Conte, Y.; Page, R.E. Age matters: Pheromone profiles of larvae differentially influence foraging behaviour in the honeybee, Apis mellifera. Anim. Behav. 2015, 99, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Michener, C.D.; McGinley, R.J.; Danforth, B.N. The Bee Genera of North. and Central America (Hymenoptera:Apoidea); Smithsonian Institution Press: Washington, WA, USA, 1994. [Google Scholar]
- Michener, C.D. The Bees of the World, 2nd ed.; Johns Hopkins University Press: Baltimore, MD, USA, 2007. [Google Scholar]
- Wenner, A.M.; Wells, P.H.; Johnson, D.L. Honey bee recruitment to food sources: Olfaction or language? Science 1969, 164, 84. [Google Scholar] [CrossRef]
- Tautz, J. Honeybee waggle dance: Recruitment success depends on the dance floor. J. Exp. Biol. 1996, 199, 1375. [Google Scholar]
- Riley, J.R.; Greggers, U.; Smith, A.D.; Reynolds, D.R.; Menzel, R. Flight paths of honeybees recruited by the waggle dance. Nature 2005, 435, 205–207. [Google Scholar] [CrossRef]
- Von Frisch, K. The Dance Language and Orientation of Bees. Cambridge University Press 1967. Sci. Educ. 1969, 53, 181–182. [Google Scholar] [CrossRef]
- Gould, J.L. Honey bee communication. Nature 1974, 252, 300–301. [Google Scholar] [CrossRef]
- Wilson, J.; Jahner, J.; Starley, L.; Calvin, C.; Ikerd, H.; Griswold, T. Sampling bee communities using pan traps: Alternative methods increase sample size. J. Insect Conserv. 2016, 20, 919–922. [Google Scholar] [CrossRef]
- Ahrent, D.K.; Caviness, C.E. Natural cross-pollination of twelve soybean cultivars in Arkansas. Crop. Sci. 1994, 34, 376–378. [Google Scholar] [CrossRef]
- Zhang, G.; St. Clair, A.L.; Dolezal, A.G.; Toth, A.L.; O’Neal, M.E. Honey Bee (Hymenoptera: Apidea) Pollen Forage in a Highly Cultivated Agroecosystem: Limited Diet Diversity and Its Relationship to Virus Resistance. J. Econ. Entomol. 2020, 113, 1062–1072. [Google Scholar] [CrossRef] [PubMed]
- Aizen, M.A.; Harder, L.D. The global stock of domesticated honey bees is growing slower than agricultural demand for pollination. Curr. Biol. 2009, 19, 915–918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seitz, N.; Traynor, K.S.; Steinhauer, N.; Rennich, K.; Wilson, M.E.; Ellis, J.D.; Rose, R.; Tarpy, D.; Sagili, R.; Caron, D.M.; et al. A national survey of managed honey bee 2014–2015 annual colony losses in the USA. J. Apic. Res. 2016, 54, 292–304. [Google Scholar] [CrossRef]
- Steinhauer, N.; Rennich, K.; Wilson, M.E.; Caron, D.M.; Lengerich, E.; Pettis, J.; Rose, R.; Skinner, J.; Tarpy, D.; Wilkes, J.; et al. A national survey of managed honey bee 2012–2013 annual colony losses in the USA: Results from the Bee Informed Partnership. J. Apic. Res. 2014, 53, 1–18. [Google Scholar] [CrossRef]
- Ragsdale, D.W.; McCornack, B.P.; Venette, R.C.; Potter, B.; MacRae, I.V.; Hodgson, E.W.; O’Neal, M.E.; Johnson, K.D.; O’Neil, J.; Difonzo, C.D.; et al. Economic Threshold for Soybean Aphid (Hemiptera: Aphididae). J. Apic. Res. 2007, 100, 1258–1267. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Question # | Pan Trap Research Objective | Figure | Result a |
|---|---|---|---|
| 1 | Does activity-density vary with pan trap color? | 2A | ✔ |
| 2 | Does activity-density vary with colony presence/absence? | 2B | ✘ |
| 3 | Does activity-density vary with placement in the interior vs. exterior of crop field? | 2C | ✔ |
| 4 | Does activity-density vary across the season? | 2D | ✔/✘ |
| 5 | When colonies are present, does activity-density correlate with bee population? | 3 | ✘ |
| 6 | Does activity-density vary with surrounding landscape composition? | 4 | ✘ |
| 7 | Does activity-density vary with distance from colonies? | 5 | ✘ |
| 8 | Does activity-density vary with number of colonies present? | 6 | ✔ |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
St. Clair, A.L.; Dolezal, A.G.; O’Neal, M.E.; Toth, A.L. Pan Traps for Tracking Honey Bee Activity-Density: A Case Study in Soybeans. Insects 2020, 11, 366. https://doi.org/10.3390/insects11060366
St. Clair AL, Dolezal AG, O’Neal ME, Toth AL. Pan Traps for Tracking Honey Bee Activity-Density: A Case Study in Soybeans. Insects. 2020; 11(6):366. https://doi.org/10.3390/insects11060366
Chicago/Turabian StyleSt. Clair, Ashley L., Adam G. Dolezal, Matthew E. O’Neal, and Amy L. Toth. 2020. "Pan Traps for Tracking Honey Bee Activity-Density: A Case Study in Soybeans" Insects 11, no. 6: 366. https://doi.org/10.3390/insects11060366
APA StyleSt. Clair, A. L., Dolezal, A. G., O’Neal, M. E., & Toth, A. L. (2020). Pan Traps for Tracking Honey Bee Activity-Density: A Case Study in Soybeans. Insects, 11(6), 366. https://doi.org/10.3390/insects11060366