Unique Fine Morphology of Mouthparts in Haematoloecha nigrorufa (Stål) (Hemiptera: Reduviidae) Adapted to Millipede Feeding


Abstract
:1. Introduction
2. Material and Methods
2.1. Insect Collecting
2.2. Samples for SEM
2.3. Feeding Behavior on Millipedes
2.4. Image Processing and Morphometric Measurement
2.5. Data Analysis
2.6. Terminology
3. Results
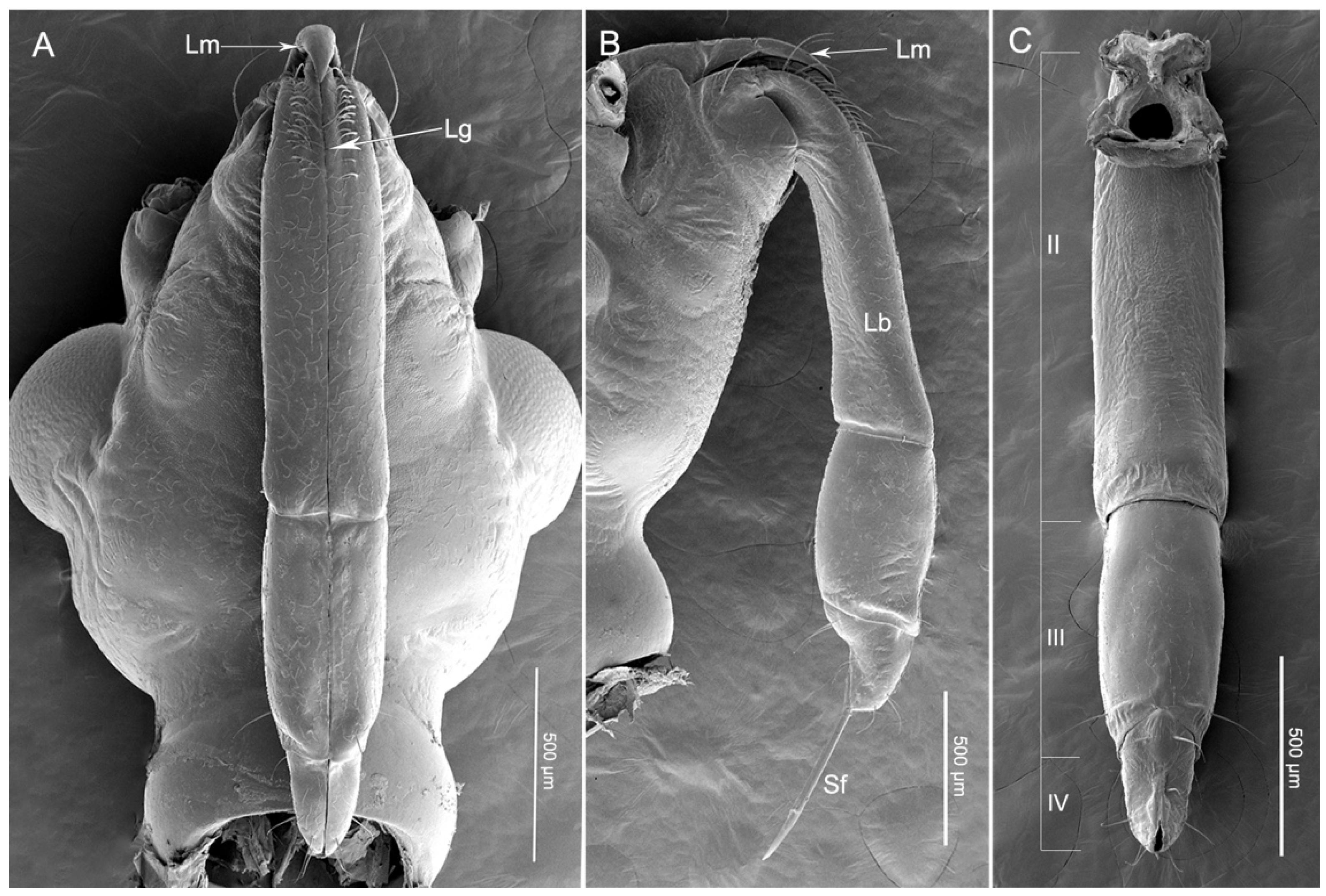
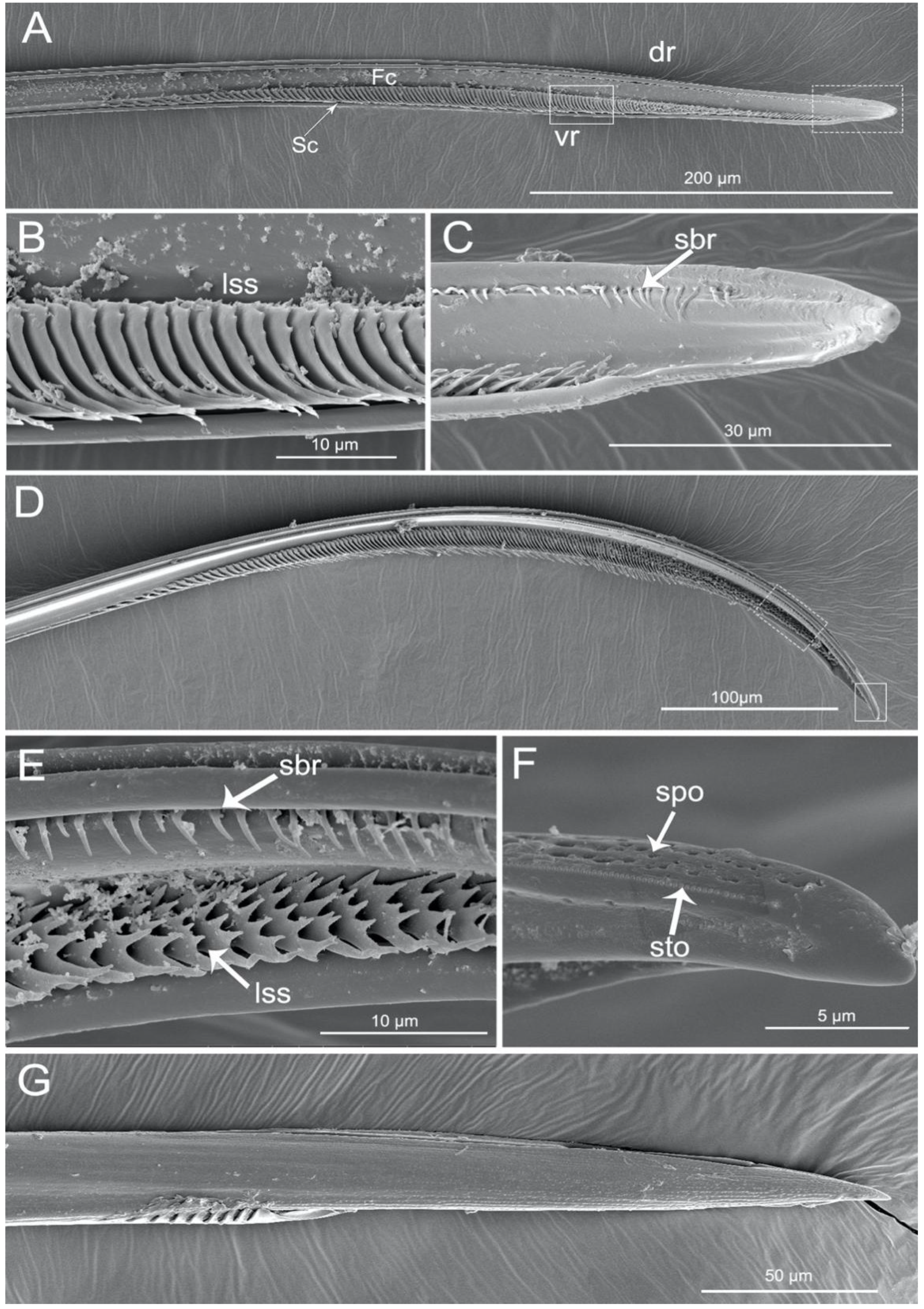
3.1. General Morphology of Mouthparts
3.2. Labrum
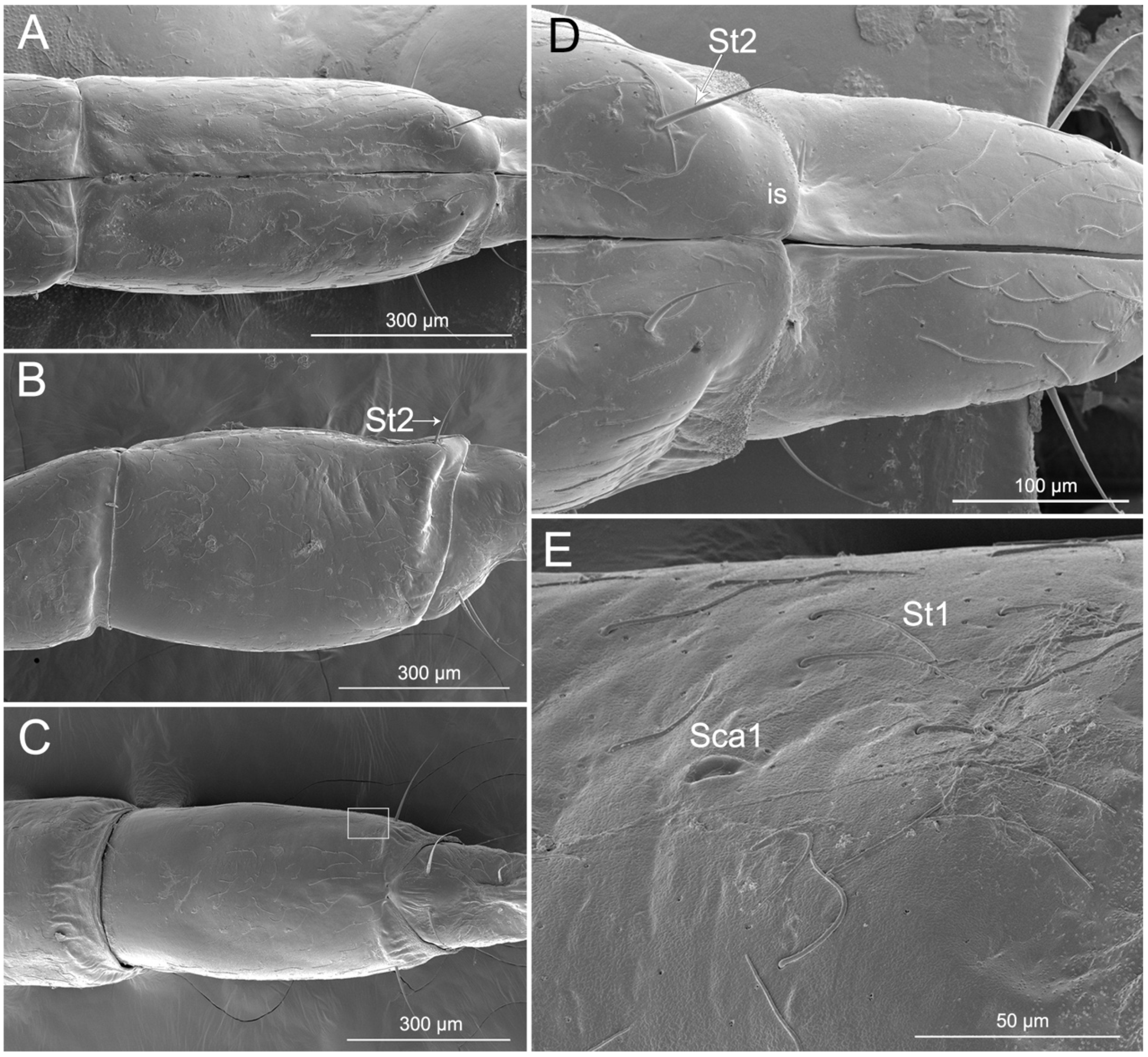
3.3. Labium
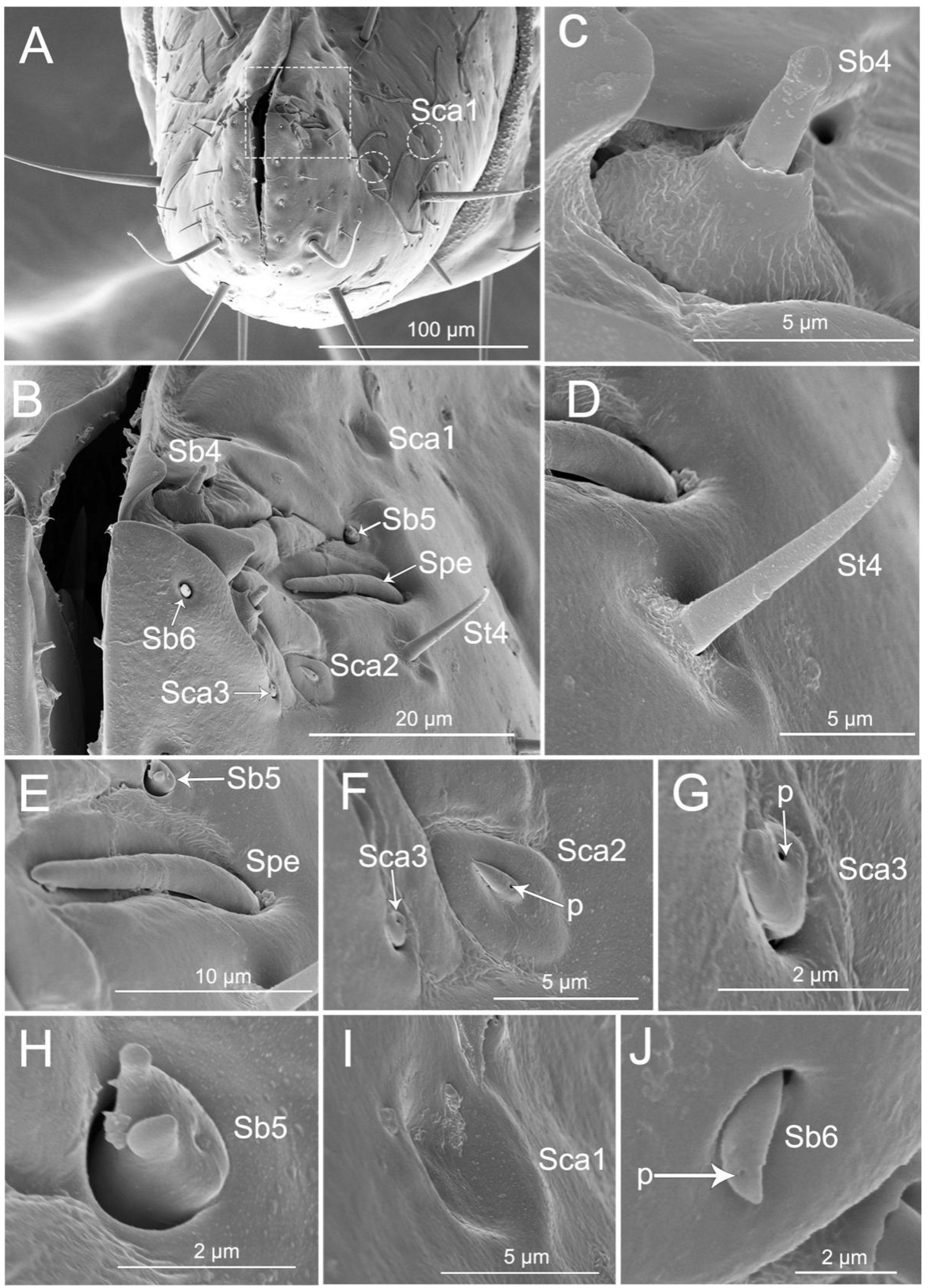
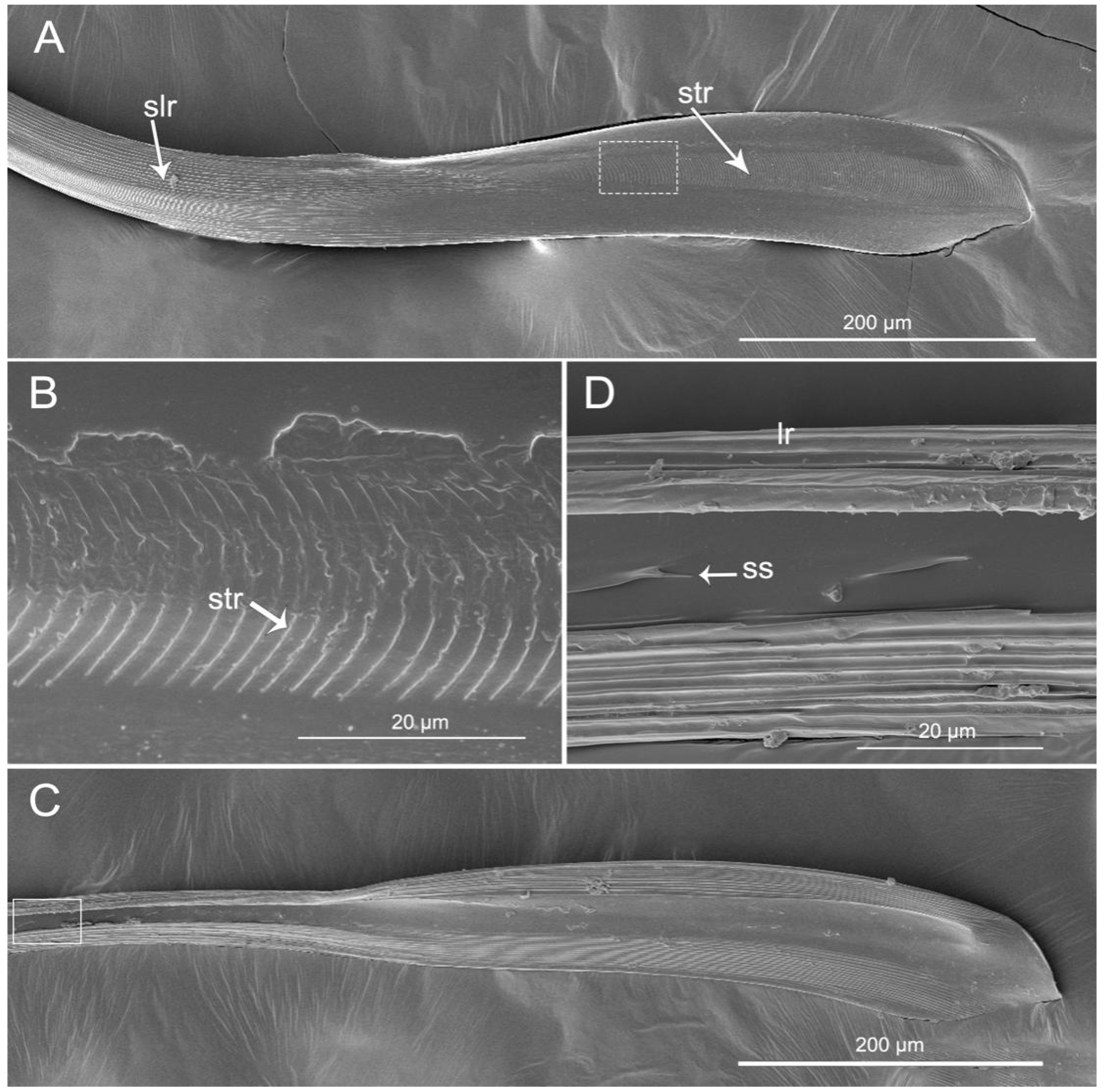
3.4. Stylet Fascicle
3.5. The Process of Feeding by Haematoloecha Nigrorufa
4. Discussion
4.1. The Mouthparts of A Millipede Specialist
4.2. Labial Sensillar System
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Snodgrass, R.E. Principles of Insect Morphology; McGraw Hill Book Co.: New York, NY, USA, 1935. [Google Scholar]
- Chapman, R.F. Communication. In The Insects: Structure and Function, 4th ed.; Chapman, R.F., Ed.; Cambridge University Press: Cambridge, UK, 1998; pp. 610–652. [Google Scholar]
- Chapman, R.F. Mouthparts. In Encyclopedia of Insects; Resh, V.H., Cardé, R.T., Eds.; Academic Press: San Diego, CA, USA, 2003; pp. 750–755. [Google Scholar]
- Cummins, K.W. Trophic relations of aquatic insects. Annu. Rev. Entomol. 1973, 1, 183–206. [Google Scholar] [CrossRef]
- Cummins, K.W. Structure and function of stream ecosystems. Bioscience 1974, 2, 631–641. [Google Scholar] [CrossRef]
- Backus, E.A. Sensory systems and behaviours which mediate hemipteran plant-feeding: A taxonomic overview. J. Insect Physiol. 1988, 34, 151–165. [Google Scholar] [CrossRef]
- Krenn, H.W. Feeding mechanisms of adult Lepidoptera: Structure, function, and evolution of the mouthparts. Annu. Rev. Entomol. 2010, 55, 307–327. [Google Scholar] [CrossRef] [Green Version]
- Schneider, D. Insect antennae. Annu. Rev. Entomol. 1964, 9, 103–122. [Google Scholar] [CrossRef]
- Zacharuk, R.Y. Antennae and sensilla. In Comprehensive Insect Physiology, Biochemistry and Pharmacology; Kerkut, G.A., Gilbert, L.I., Eds.; Pergamon Press: Oxford, UK, 1985; pp. 1–69. [Google Scholar]
- McGavin, G.C. Food and Feeding. In Bugs of the World; Blandford: London, UK, 1993. [Google Scholar]
- Butt, F.H. Comparative study of mouth parts of representative Hemiptera-Heteroptera. Ann. Entomol. Soc. Am. 1943, 38, 124. [Google Scholar] [CrossRef]
- Cobben, R.H. Evolutionary Trends in Heteroptera. In Part II Mouthpart-Structures and Feeding Strategies; Mededelingen Landbouwhogeschool: Wageningen, The Netherlands, 1978; pp. 1–407. [Google Scholar]
- Muir, F.; Kershaw, J.C. On the Homologies and Mechanism of the Mouth-Parts of Hemiptera. A. J. Entomol. 1911, 18, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Gullan, P.J.; Cranston, P.S. (Eds.) The Insects: An Outline of Entomology. In Sensory Systems and Behavior, 5th ed.; Ch4. Wiley Blackwell: Oxford, UK, 2014. [Google Scholar]
- Wang, Y.; Brożek, J.; Dai, W. Sensory armature and stylets of the mouthparts of Stephanitis nashi (Hemiptera: Cimicomorpha: Tingidae), their morphology and function. Micron 2020, 132, 102840. [Google Scholar] [CrossRef]
- Wang, Y.; Brożek, J.; Dai, W. Morphological Disparity of the Mouthparts in Polyphagous Species of Largidae (Heteroptera: Pentatomomorpha: Pyrrhocoroidea) Reveals Feeding Specialization. Insects 2020, 11, 145. [Google Scholar] [CrossRef] [Green Version]
- Engel, M.S. A new interpretation of the oldest fossil bee (Hymenoptera: Apidae). Am. Mus. Novit. 2000, 3296, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Brożek, J.; Chłond, D. Morphology, arrangement and classification of sensilla on the apical segment of labium in Peiratinae (Hemiptera: Heteroptera: Reduviidae). Zootaxa 2010, 2476, 39–52. [Google Scholar] [CrossRef]
- Brożek, J. A comparison of external and internal maxilla and mandible morphology of water bugs (Hemiptera: Heteroptera: Nepomorpha). Zootaxa 2013, 3635, 340–378. [Google Scholar] [CrossRef] [PubMed]
- Brożek, J. Phylogenetic signals from Nepomorpha (Insecta: Hemiptera: Heteroptera) mouthparts: Stylets bundle, sense organs, and labial segments. Sci. World J. 2014, 2014, 237854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brożek, J.; Zettel, H. A comparison of the external morphology and functions of labial tip sensilla in semiaquatic bugs (Hemiptera: Heteroptera: Gerromorpha). Eur. J. Entomol. 2014, 111, 275–297. [Google Scholar] [CrossRef] [Green Version]
- Parveen, S.; Ahmad, A.; Brożek, J.; Ramamurthy, V.V. Morphological diversity of the labial sensilla of phytophagous and predatory Pentatomidae (Hemiptera: Heteroptera), with reference to their possible functions. Zootaxa 2015, 4039, 246–253. [Google Scholar] [CrossRef]
- Wang, Y.; Dai, W. Fine structure of mouthparts and feeding performance of Pyrrhocoris sibiricus Kuschakevich with remarks on the specialization of sensilla and stylets for seed feeding. PLoS ONE 2017, 12, e0177209. [Google Scholar] [CrossRef]
- Wang, Y.; Li, L.F.; Dai, W. Fine Morphology of the Mouthparts in Cheilocapsus nigrescens (Hemiptera: Heteroptera: Miridae) Reflects Adaptation for Phytophagous Habits. Insects 2019, 10, 17. [Google Scholar] [CrossRef] [Green Version]
- Taszakowski, A.; Nowińska, A.; Brożek, J. Morphological study of the labial sensilla in Nabidae (Hemiptera: Heteroptera: Cimicomorpha). Zoomorphology 2019, 138, 483–492. [Google Scholar] [CrossRef] [Green Version]
- Peregrine, D.J. Fine structure of sensilla basiconica on the labium of the cotton stainer, Dysdercus fasciatus (Signoret) (Heteroptera: Pyrrhocoridae). Int. J. Insect Morphol. Embryol. 1972, 1, 241–251. [Google Scholar] [CrossRef]
- Schoonhoven, L.M.; Henstra, S. Morphology of some rostrum receptors in Dysdercus spp. Neth. J. Zool. 1972, 22, 343–346. [Google Scholar] [CrossRef] [Green Version]
- Avé, D.; Frazier, J.L.; Hatfield, L.D. Contact chemoreception in the tarnished plant bug Lygus lineolaris. Ent. Exp. Appl. 1978, 24, 217–227. [Google Scholar] [CrossRef]
- Hatfield, L.D.; Frazier, J.L. Ultrastructure of the labial tip sensilla of the tarnished plant bug, Lygus lineolaris (P. De Beauvois) (Hemiptera: Miridae). Int. J. Insect Morphol. Embryol. 1980, 9, 59–66. [Google Scholar] [CrossRef]
- Gaffal, K.P. Terminal sensilla on the labium of Dysdercus intermedius Distant (Heteroptera: Pyrrhocoridae). Int. J. Insect Morphol. Embryol. 1981, 10, 1–6. [Google Scholar] [CrossRef]
- Rani, P.U.; Madhavendra, S.S. Morphology and distribution of antennal sense organs and diversity of mouthpart structures in Odontopus nigricornis (Stål) and Nezara viridula L. (Hemiptera). Int. J. Insect Morphol. Embryol. 1995, 24, 119–132. [Google Scholar] [CrossRef]
- Rani, P.U.; Madhavendra, S.S. External morphology of antennal and rostral sensillae in four hemipteran insects and their possible role in host plant selection. Int. J. Trop. Insect Sci. 2005, 25, 198–207. [Google Scholar] [CrossRef]
- Anderson, W.G.; Hengmoss, T.M.; Baxendale, F.P.; Baird, L.M.; Sarath, G.; Higley, L. Chinch bug (Hemiptera: Blissidae) mouthpart morphology, probing frequencies, and locations on resistant and susceptible germplasm. J. Econ. Entomol. 2006, 99, 212–221. [Google Scholar] [CrossRef] [PubMed]
- Brożek, J. Morphology and arrangement of the labial sensilla of the water bugs. Bull. Insectol. 2008, 61, 67–168. [Google Scholar]
- Depieri, R.A.; Panizzi, A.R. Rostrum length, mandible serration, and food and salivary canals areas of selected species of stink bugs (Heteroptera, Pentatomidae). Rev. Brasileira De Entomol. 2010, 54, 584–587. [Google Scholar] [CrossRef] [Green Version]
- Depieri, R.A.; Siqueia, F.; Panizzi, A.R. Aging and food source effects on mandibular stylets teeth wear of Phytophagous Stink Bug (Heteroptera: Pentatomidae). Neotrop. Entomol. 2010, 39, 952–956. [Google Scholar] [CrossRef] [Green Version]
- Lucini, T.; Panizzi, A.R.; Backus, E.A. Characterization of an EPG Waveform Library for Redbanded Stink Bug, Piezodorus guildinii (Hemiptera: Pentatomidae), on Soybean Plants. Ann. Entomol. Soc. Am. 2016, 109, 198–210. [Google Scholar] [CrossRef] [Green Version]
- Backus, E.A.; Serrano, M.S.; Ranger, C.M. Mechanisms of Hopperburn: An overview of insect taxonomy, behavior, and physiology. Annu. Rev. Entomol. 2005, 50, 125–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Backus, E.A. Anatomical and sensory mechanisms of leafhopper and planthopper feeding behavior. In The leafhoppers and Planthoppers; Nault, L.R., Rodriguez, J.G., Eds.; John Wiley & Sons: New York, NY, USA, 1985; pp. 163–194. [Google Scholar]
- Feir, D.; Beck, S.D. Feeding behavior of the large milkweed bug, Oncopeltus fasciatus. Ann. Entomol. Soc. Am. 1963, 56, 224–229. [Google Scholar] [CrossRef]
- Saxena, K.N. Mode of ingestion in a heteropteran insect Dysdercus koenigii (F.) (Pyrrhocoridae). J. Insect Physiol. 1963, 9, 47–71. [Google Scholar] [CrossRef]
- Weirauch, C. Cladistic analysis of Reduviidae (Heteroptera: Cimicomorpha) based on morphological characters. Syst. Entomol. 2008, 33, 229–274. [Google Scholar] [CrossRef]
- Weirauch, C. From four- to three-segmented labium in Reduviidae (Hemiptera: Heteroptera). Acta Entomol. Musei Nationalis Pragae 2008, 48, 331–344. [Google Scholar]
- Catalá, S. Sensilla associated with the Rostrum of eight species of Triatominae. J. Morpho. 1996, 228, 195–201. [Google Scholar] [CrossRef]
- Cohen, A.C. Feeding adaptations of some predaceous Hemiptera. Ann. Entomol. Soc. Am. 1990, 83, 1215–1223. [Google Scholar] [CrossRef]
- Forthman, M.; Weirauch, C. Millipede assassins and allies (Heteroptera: Reduviidae: Ectrichodiinae, Tribelocephalinae): Total evidence phylogeny, revised classification and evolution of sexual dimorphism. Syst. Entomol. 2017, 42, 575–595. [Google Scholar] [CrossRef]
- Brożek, J.; Herczek, A. Internal structure of the mouthparts of true bugs (Hemiptera: Heteroptera). Pol. J. Entomol. 2004, 73, 79–106. [Google Scholar]
- Spooner, C.S. The phylogeny of the Hemiptera based on a study of the head capsule. Illinois Biol. Monogr. 1938, 16, 1–102. [Google Scholar]
- Gupta, A.P. A consideration of the systematic position of the Saldidae and the Mesoveliidae (Hemiptera: Heteroptera). Proc. Ent. Soc. Washington 1963, 65, 31–38. [Google Scholar]
- Štys, P. On the morphology of the labrum in Heteroptera. Acta Ent. Bohemoslov. 1969, 66, 150–158. [Google Scholar]
- Kumar, M.; Sahayaraj, K. Gross morphology and histology of head and salivary apparatus of the predatory bug, Rhynocoris marginatus. J. Insect Sci. 2012, 12, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forthman, M.; Weirauch, C. Phylogenetics and biogeography of the endemic Madagascan millipede assassin bugs (Hemiptera: Reduviidae: Ectrichodiinae). Mol. Phylogenet Evol. 2016, 100, 219–233. [Google Scholar] [CrossRef] [Green Version]
- Forthman, M.; Weirauch, C. Toxic associations: A review of the predatory behaviors of millipede assassin bugs (Hemiptera: Reduviidae: Ectrichodiinae). Eur. J. Entomol. 2012, 109, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Altner, H.; Prillinger, L. Ultrastructure of invertebrate chemo-, thermo-, and hygroreceptors and its functional significance. Int. Rev. Cytol. 1980, 67, 69–139. [Google Scholar]
- Frazier, J.L. Nervous system: Sensory system. In Fundamentals of Insect Physiology; Blum, M.S., Ed.; John Wiley & Sons: New York, NY, USA, 1985; pp. 287–356. [Google Scholar]
- Sahayaraj, K.; Kanna, A.V.; Kumar, S.M. Gross morphology of feeding canal, salivary apparatus and digestive enzymes of salivary gland of Catamirus brevipennis (Serville) (Hemiptera: Reduviidae). J. Entomol. Res. Soc. 2010, 12, 37–50. [Google Scholar]
- Hwang, W.S.; Weirauch, C. Evolutionary history of assassin bugs (Insecta: Hemiptera: Reduviidae): Insights from divergence dating and ancestral state reconstruction. PLoS ONE 2012, 7, e45523. [Google Scholar] [CrossRef] [Green Version]
- Schuh, R.T.; Štys, P. Phylogenetic analysis of cimicomorphan family relationships (Heteroptera). J. N. Y. Entomol. Soc. 1991, 99, 298–350. [Google Scholar]
- Schuh, R.T.; Weirauch, C.H.; Wheeler, W.C. Phylogenetic relationships within the Cimicomorpha (Hemiptera: Heteroptera): A total-evidence analysis. Syst. Entomol. 2009, 34, 15–48. [Google Scholar] [CrossRef]
- Weirauch, C.; Bérenger, J.-M.; Berniker, L.; Forero, D.; Forthman, M.; Frankenberg, S.; Freedman, A.; Gordon, E.; Hoey-Chamberlain, R.; Hwang, W.S.; et al. An illustrated identification key to assassin bug subfamilies and tribes (Hemiptera: Reduviidae). Can. J. Arthropod Identif. 2014. [Google Scholar] [CrossRef]
- Schuh, R.T.; Slater, J.A. True Bugs of the World (Hemiptera: Heteroptera). In Classification and Natural History; Cornell University Press: New York, NY, USA, 1995. [Google Scholar]
- Green, W.E. The President’s address. Proc. Entomol. Soc. Lond. 1925, 1924, clxi–ccii. [Google Scholar]
- Cachan, P. Etude de la prédation chez les Réduvides de larégion Éthiopienne. 1. La prédationengroupe chez Ectrichodiagigas H-Sch. Phys. Comp. Oecol. 1952, 2, 378–385. [Google Scholar]
- Miller, N.C.E. The Biology of the Heteroptera; E. W. Classey: Hampton, VA, USA, 1971; p. 206. [Google Scholar]
- Haridass, E.T. Biological and Ethological Studies on some South Indian Reduviids (Hemiptera–Reduviidae). Ph.D. Thesis, University of Madras, Madras, India, 1978. [Google Scholar]
- Haridass, E.T.; Ananthakrishnan, T.N. Models for the predatory behavior of some reduviids from southern India (Insecta–Heteroptera–Reduviidae). Proc. Indian AS Anim. Sci. 1980, 89, 387–402. [Google Scholar] [CrossRef]
- Rathore, Y.K. Studies on the mouth-parts and feeding mechanism in Dysdercus cingulatus Fabr. (Pyrrhocoridae: Heteroptera). Indian J. Entomol. 1961, 23, 163–185. [Google Scholar]
- Faucheux, M.M. Relations entre l’ultrastructure des stylets manibulaires et maxillaires et la prise de nourriture chez les insects Hemipteres. CR Acad. Sci. Paris (Ser. D) 1975, 281, 41–44. [Google Scholar]
- Boyd, D.W. Digestive enzymes and stylet morphology of Deraeocoris nigritulus (Uhler) (Hemiptera: Miridae) reflect adaptations for predatory habits. Ann. Entomol. Soc. Am. 2003, 96, 667–671. [Google Scholar] [CrossRef] [Green Version]
- Swart, C.C.; Felgenhauer, B.E. Structure and function of the mouthparts and salivary gland complex of the giant waterbug, Belostoma lutarium (Stål) (Hemiptera: Belostomatidae). Ann. Entomol. Soc. Am. 2003, 96, 870–882. [Google Scholar] [CrossRef]
- Roitberg, B.D.; Gillespie, D.R.; Quiring, D.M.J.; Alma, C.R.; Jenner, W.H.; Perry, J.; Peterson, J.H.; Salomon, M.; Van Laerhoven, S. The cost of being anomnivore: Mandible wear from plant feeding in a true bug. Naturwissenschaften 2005, 92, 431–434. [Google Scholar] [CrossRef]
- Romani, R.; Salerno, G.; Frati, F.; Conti, E.; Isidoro, N.; Bin, F. Oviposition behaviour in Lygus rugulipennis: Amorpho-functional study. Entomol. Exp. Appl. 2005, 115, 17–25. [Google Scholar] [CrossRef]
- Bérenger, J.; Pluot-Sigwalt, D. Notes sur Micrauchenus lineola (Fabricius 1787), espѐcetermitophile et termitophage (Heteroptera: Reduviidae: Harpactorinae, Apiomerini). Ann. la Soc. Entomol. Fr. 2009, 45, 129–133. [Google Scholar] [CrossRef] [Green Version]
- Stubbins, F.L.; Mitchell, P.L.; Turnbull, M.W.; Reay-Jones, F.P.F.; Greene, J.K. Mouthpart morphology and feeding behavior of the invasive kudzu bug, Megacopta cribraria (Hemiptera: Plataspidae). Invertebr. Biol. 2017, 136, 309–320. [Google Scholar] [CrossRef]
- Ambrose, D.P. Economic importance of Heteroptera. In Heteroptera of Economic Importance; Schaefer, C.W., Panizzi, A.R., Eds.; CRC Press: Boca Raton, FL, USA, 2000; pp. 695–712. [Google Scholar]
- Rosa, J.A.D.; Barata, J.M.S.; Cilense, M.; Neto, F.M.B. Head morphology of 1st and 5th instar nymphs of Triatoma circummaculata and Triatoma rubrovaria (Hemiptera, Reduviidae). Int. J. Insect Morphol. Embryol. 1999, 28, 363–375. [Google Scholar] [CrossRef]
- Hildebrand, J.G.; Shepherd, G.M. Mechanisms of olfactory discrimination: Converging evidence for common principles across phyla. Annu. Rev. Neurosci. 1997, 20, 595–631. [Google Scholar] [CrossRef] [PubMed]
- Baker, G.T.; Chen, X.; Ma, P.W.K. Labial tip sensilla of Blissus leucopterus leucopterus (Hemiptera: Blissidae): Ultrastructure and behavior. Insect Sci. 2008, 15, 271–275. [Google Scholar] [CrossRef]
- Zacharuk, R.Y. Ultrastructure and function of insect chemosensilla. Annu. Rev. Entomol. 1980, 25, 27–47. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | Position | Length (μm) | Width (μm) | N |
|---|---|---|---|---|
| Female | Lb | 2058.0 ± 25.8 | - | 12 |
| L2 | 1174.8 ± 35.9 | 215.3 ± 4.4 | 12 | |
| L3 | 587.4 ± 15.7 | 301.7 ± 5.8 | 12 | |
| L4 | 298 ± 18.3 | 194 ± 2.8 | 12 | |
| Male | Lm | 476.5 ± 8 | - | 4 |
| Lb | 1875.2 ± 113.5 | - | 3 | |
| Lb2 | 1011 ± 131.2 | 219 ± 8.7 | 3 | |
| Lb3 | 583.6 ± 14.2 | 288.2 ± 1.2 | 3 | |
| Lb4 | 285.8 ± 18.9 | 183 ± 4.4 | 3 | |
| Md (spatulate apex) | 507.4 ± 15.3 | - | 4 | |
| RMx (dorsal short bristles) | 104.3 ± 6.5 | - | 3 | |
| RMx (ventral lamellate structures) | 424.2 ± 13.7 | - | 3 | |
| LMx (dorsal short spines) | 15.8 ± 0.5 | - | 3 | |
| LMx (ventral short bristles) | 76.4 ± 3.6 | - | 3 |
| Type | Distribution and Number | Length (μm) | Basal Diameter (μm) | Shape | Socket | Surface | Pore | Category | Function | N |
|---|---|---|---|---|---|---|---|---|---|---|
| St1 | Lm, Lb2-Lb4 | 42.8 ± 1.7 | 1.45 ± 0.1 | Hair in pit | Fnflexible | Smooth | Wp (Multiporous) | Chemoreceptive sensilla | Olfactory | 20 |
| St2 | Lb2 (1 pair), Lb3 (2pairs), Lb4 (5pairs) | 142.8 ± 6 | 5.3 ± 0.2 | Hair, peg | Flexible | Grooved | No | Mechanoreceptive sensilla | Tactile | 11 |
| St3 | Lb2 (about 16 pairs) | 103.8 ± 5.8 | 5.2 ± 0.1 | Hair | Flexible | Grooved | No | Mechanoreceptive sensilla | Tactile | 10 |
| St4 | Lb3 (6 pairs) | 9.4 ± 0.6 | 1.6 ± 0.1 | Hair, peg | Flexible | Smooth | No | Mechanoreceptive sensilla | Tactile | 14 |
| Sb1 | 3 pairs at the base of the second segment, 1 pair on the junction between the third and fourth segment | 32.7 ± 2.5 | 4.9 ± 0.1 | Peg | Flexible | Smooth | Wp (Uniporous) | Proprioceptive sensilla | Perceive the degree of flexion of the joint | 10 |
| Sb2 | Lb2 (2 pairs) | 9.5 ± 0.7 | 1.4 ± 0.1 | Peg in pit | Inflexible | Smooth | No | Thermo-hygroreceptive sensilla | Temperature/humidity | 6 |
| Sb3 | Lb4 (4 pairs) | 6.0 ± 0.4 | 1.1 ± 0.1 | Peg | Flexible | Smooth | No | Mechanoreceptive sensilla | Tactile | 14 |
| Sb4 | SF (2 pairs) | 5.1 ± 0.4 | 4.2 ± 0.2 | Peg (sensillum coleoconicum) | Inflexible | Smooth | Tp | Chemoreceptive sensilla | Gustatory | 8 |
| Sb5 | SF (1 pair) | 1.3 ± 0.1 | 1.2 ± 0.1 | Peg | Flexible | Smooth | Tp | Chemoreceptive sensilla | Gustatory | 4 |
| Sb6 | Tip (1 pair) | 1.7 ± 0.1 | 1.2 ± 0.1 | Peg in pit | Flexible | Smooth | Tp | Chemoreceptive sensilla | Gustatory | 4 |
| Sca1 | Lb2-Lb4 | - | 7.2 ± 0.7 | Oval plate | Inflexible | Smooth | Tp | Proprioceptive sensilla | Perceive the degree of flexion of the joint | 8 |
| Sca2 | SF (1 pair) | - | 5.5 ± 0.3 | Dome-like structures | Inflexible | Smooth | Tp | Proprioceptive sensilla | Perceive the degree of flexion of the joint | 4 |
| Sca3 | SF (1 pair) | - | 1.2 ± 0.1 | Oval plate | Inflexible | Smooth | Tp | Proprioceptive sensilla | Perceive the degree of flexion of the joint | 4 |
| Spe | SF (1 pair) | 16.8 ± 2.1 | 2.2 ± 0.1 | Dome-elongated | Inflexible | Smooth | Tp | Chemoreceptive sensilla | Gustatory | 8 |
| Sm | Lm, Lb2-Lb4 | - | 1.5 ± 0.1 | Pegs in cavity surrounded by fingerlike structures | Inflexible | Smooth | No | Thermo-hygroreceptive sensilla | Temperature/humidity | 20 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhang, J.; Wang, W.; Brożek, J.; Dai, W. Unique Fine Morphology of Mouthparts in Haematoloecha nigrorufa (Stål) (Hemiptera: Reduviidae) Adapted to Millipede Feeding. Insects 2020, 11, 386. https://doi.org/10.3390/insects11060386
Wang Y, Zhang J, Wang W, Brożek J, Dai W. Unique Fine Morphology of Mouthparts in Haematoloecha nigrorufa (Stål) (Hemiptera: Reduviidae) Adapted to Millipede Feeding. Insects. 2020; 11(6):386. https://doi.org/10.3390/insects11060386
Chicago/Turabian StyleWang, Yan, Junru Zhang, Wanshan Wang, Jolanta Brożek, and Wu Dai. 2020. "Unique Fine Morphology of Mouthparts in Haematoloecha nigrorufa (Stål) (Hemiptera: Reduviidae) Adapted to Millipede Feeding" Insects 11, no. 6: 386. https://doi.org/10.3390/insects11060386
APA StyleWang, Y., Zhang, J., Wang, W., Brożek, J., & Dai, W. (2020). Unique Fine Morphology of Mouthparts in Haematoloecha nigrorufa (Stål) (Hemiptera: Reduviidae) Adapted to Millipede Feeding. Insects, 11(6), 386. https://doi.org/10.3390/insects11060386