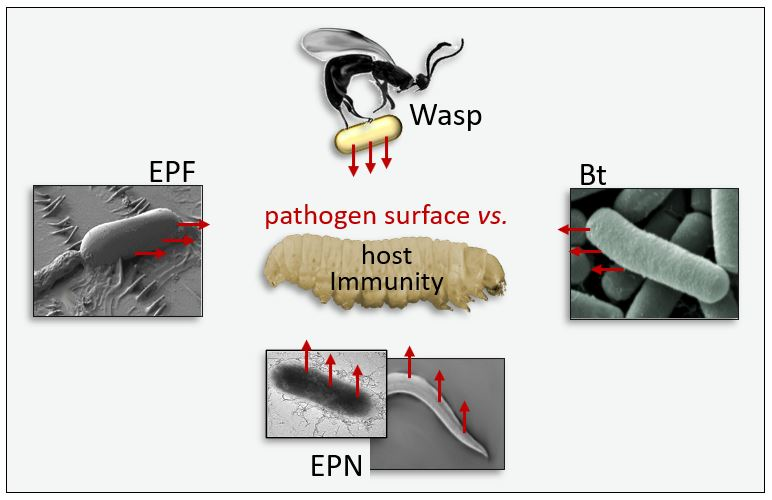
When Appearance Misleads: The Role of the Entomopathogen Surface in the Relationship with Its Host



Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
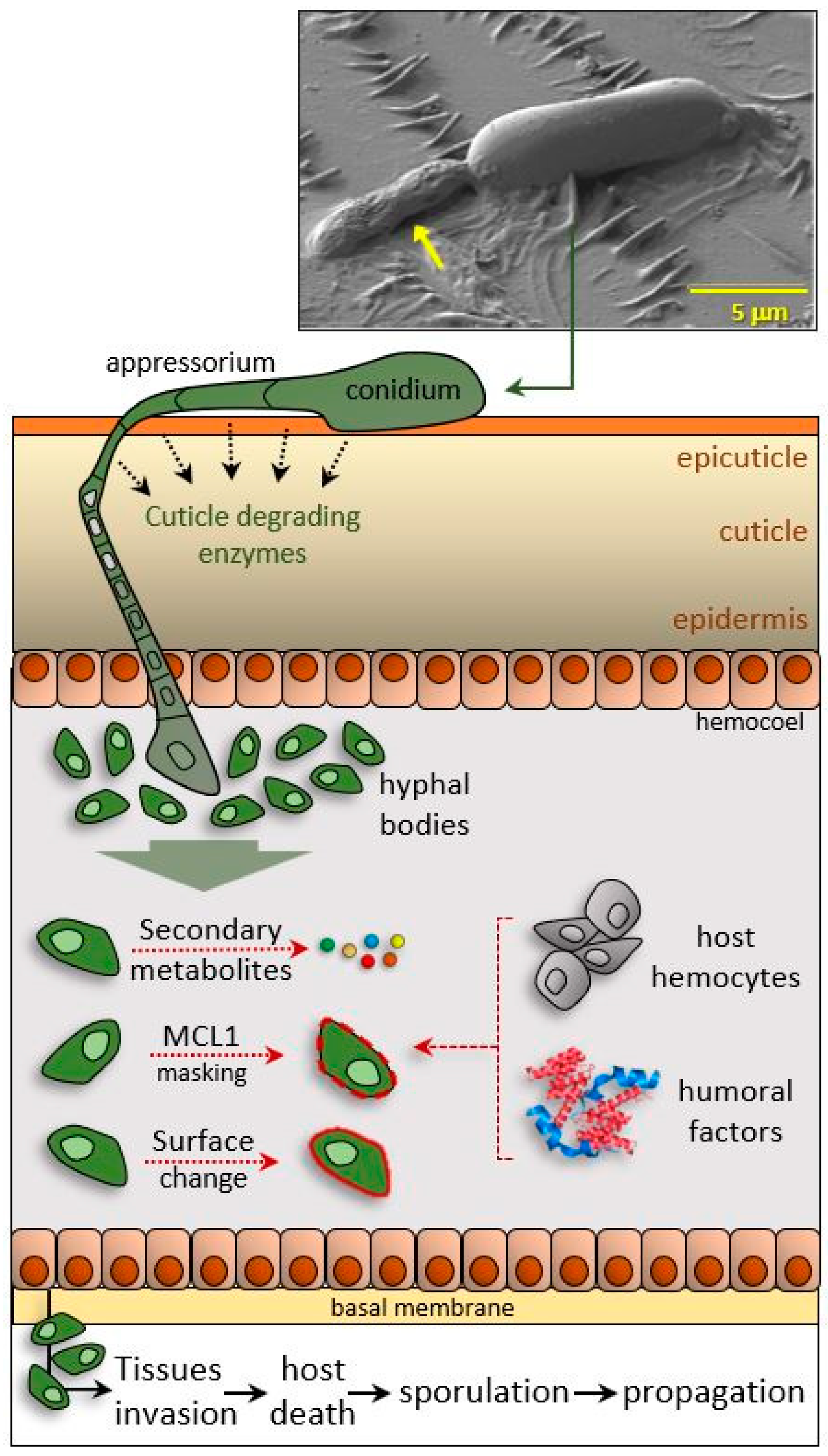{kind=link}
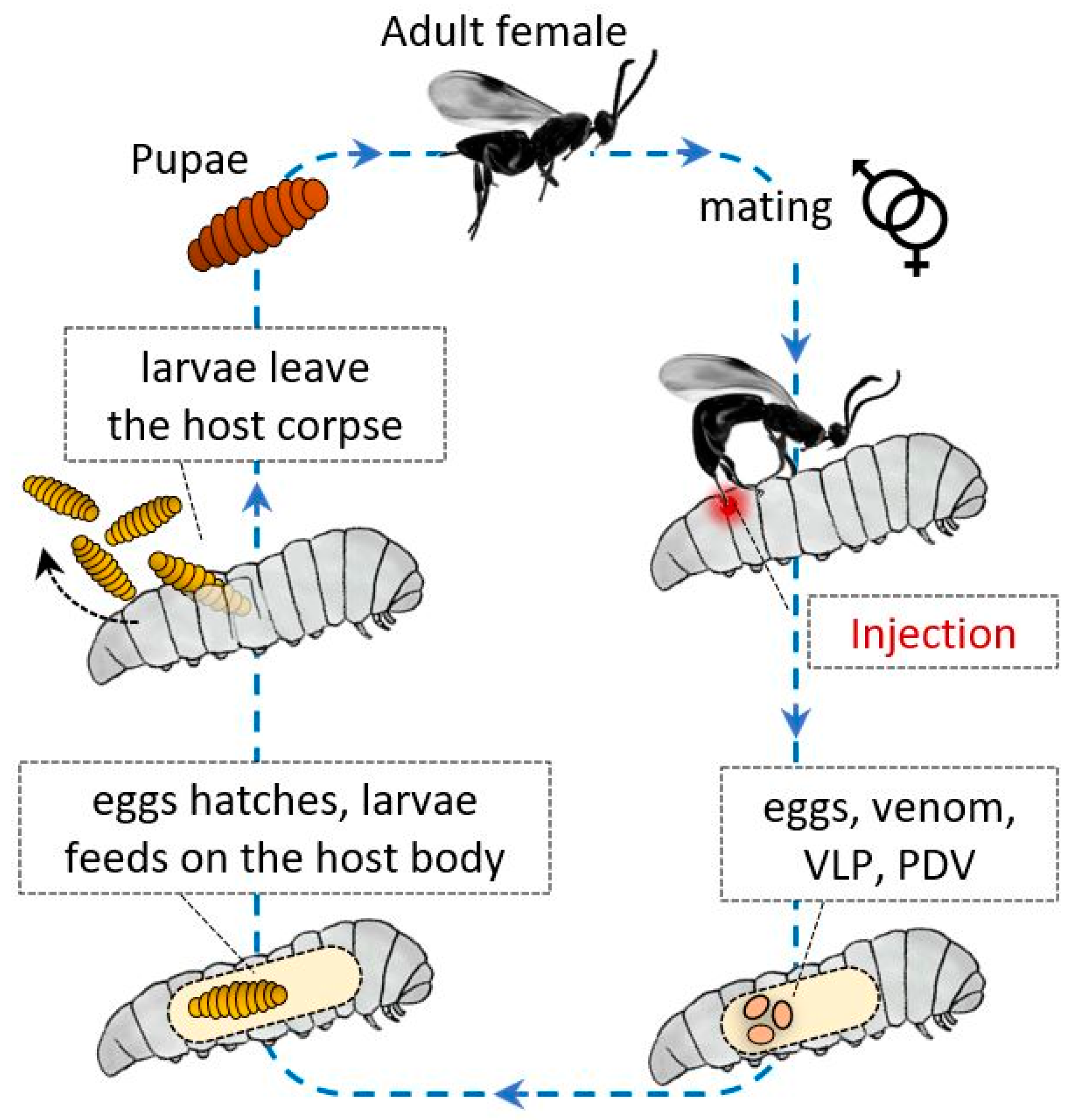{kind=link}
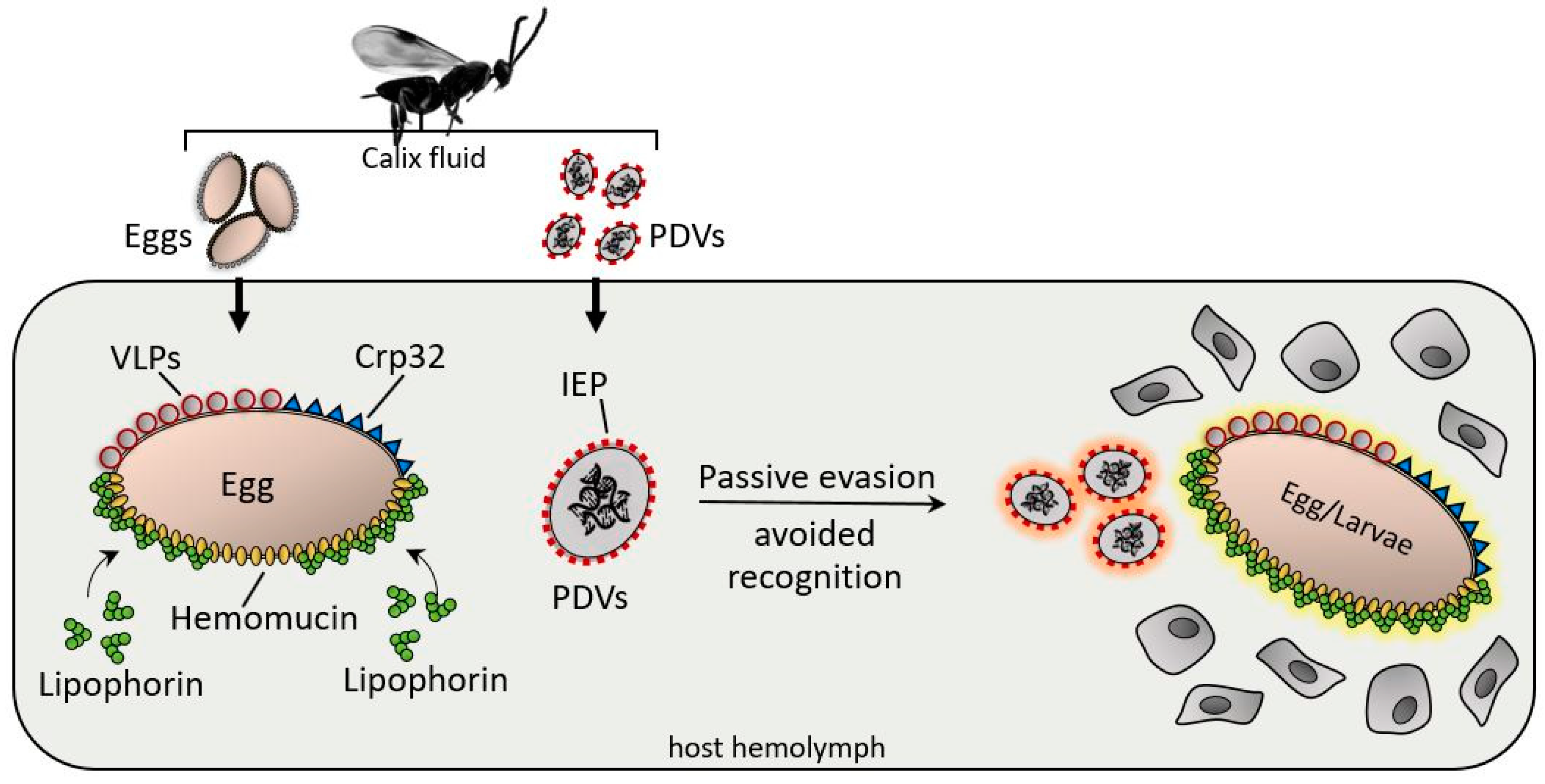{kind=link}
{kind=link}
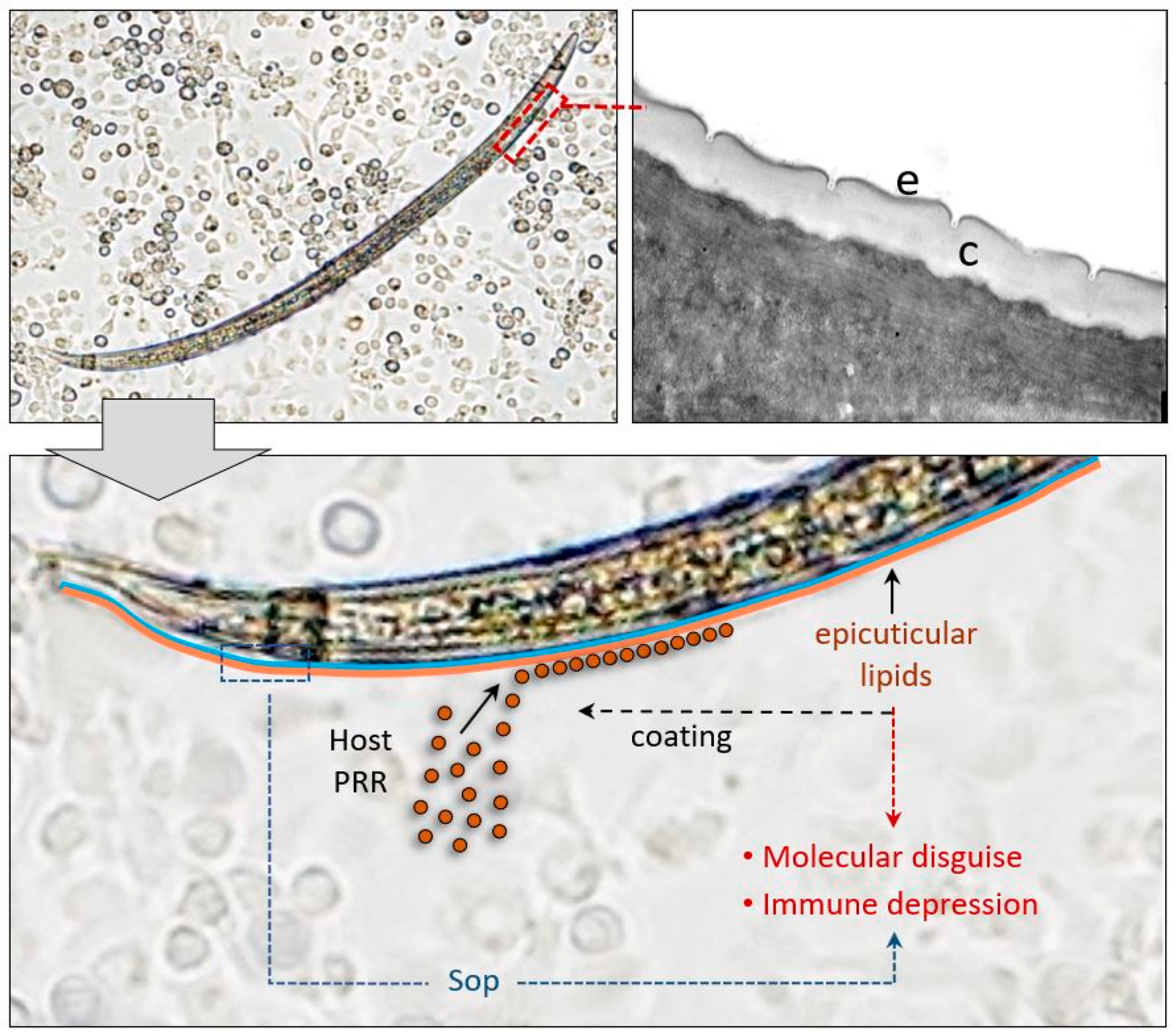{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
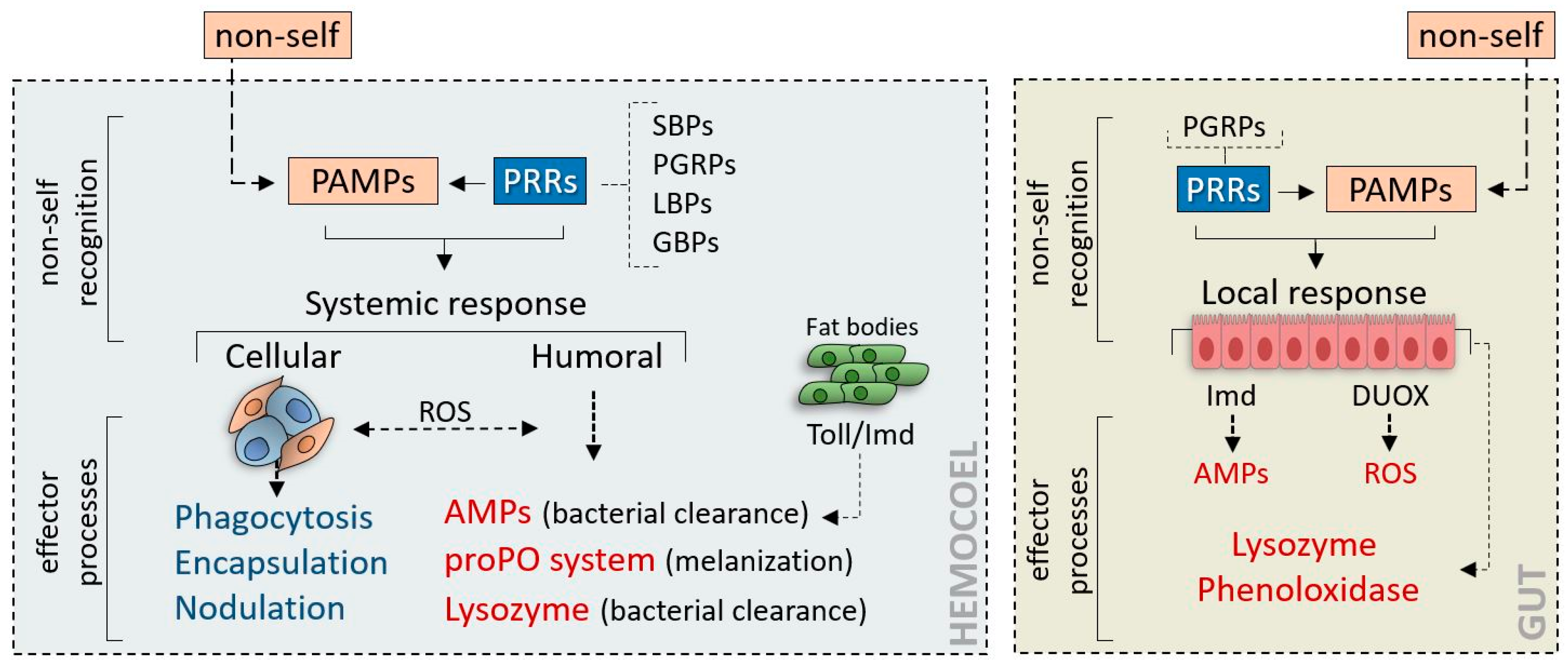
1.1. An Overview of Insect Immunity
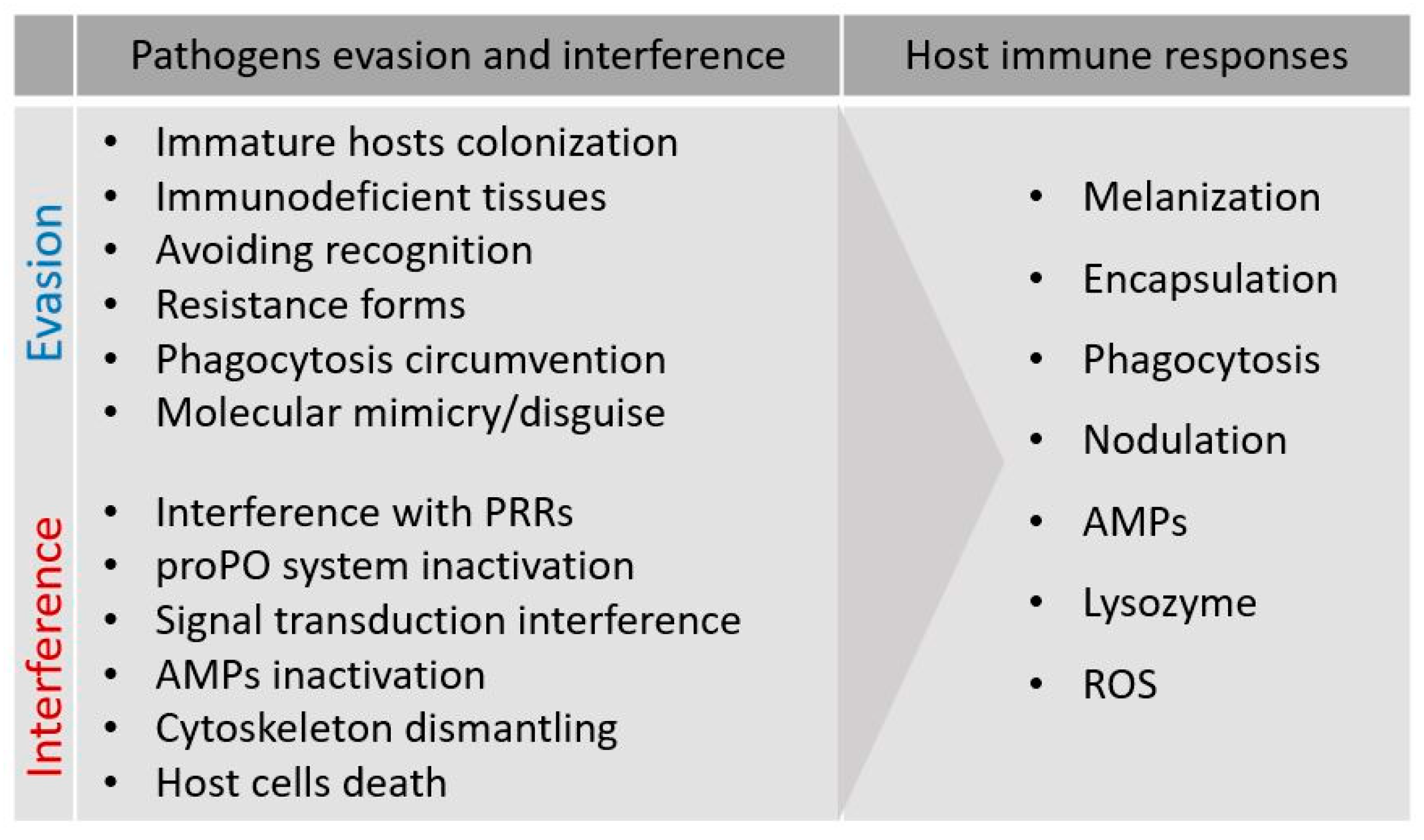
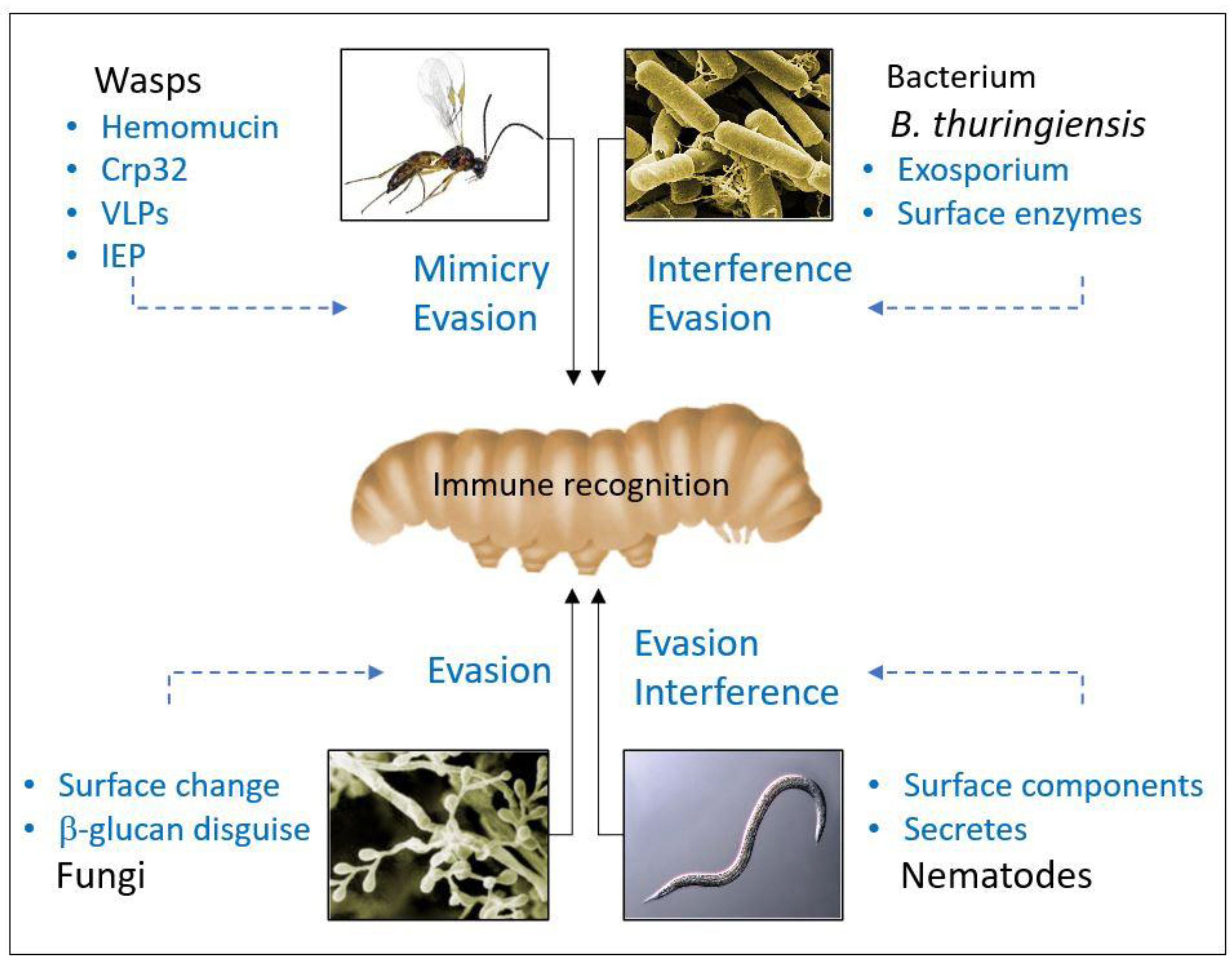
1.2. Strategies of Parasites and Microorganisms
2. Entomopathogens
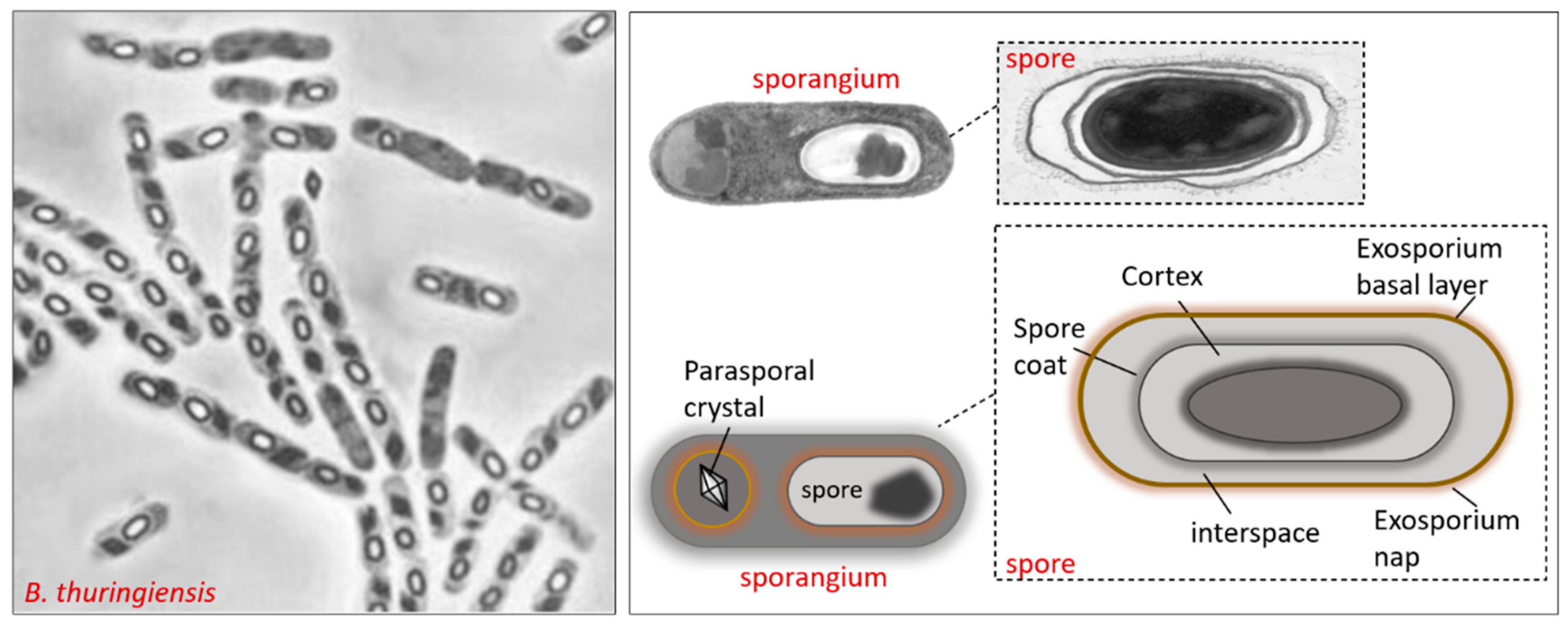
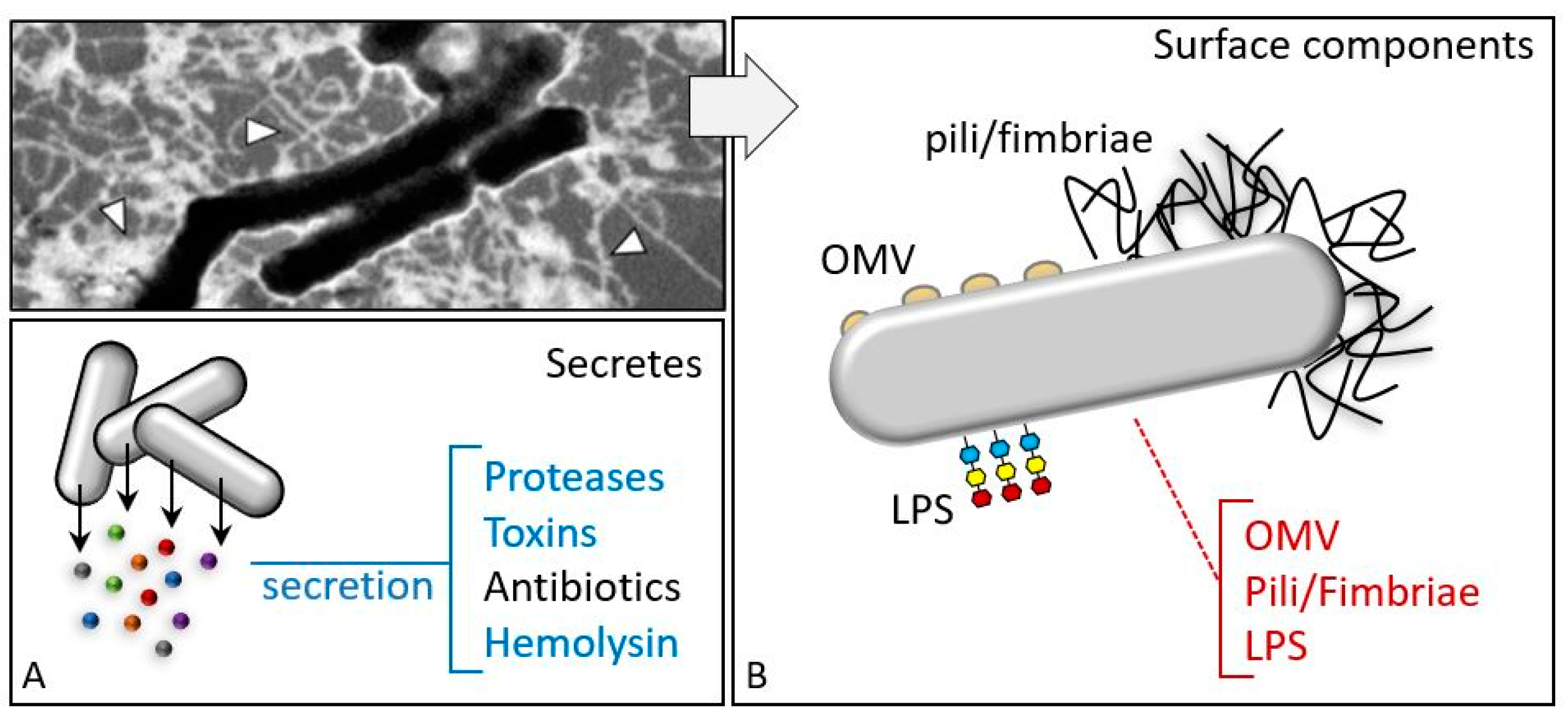
2.1. Bacillus thuringiensis
2.2. Entomopathogenic Fungi
2.3. Entompathogen Wasps
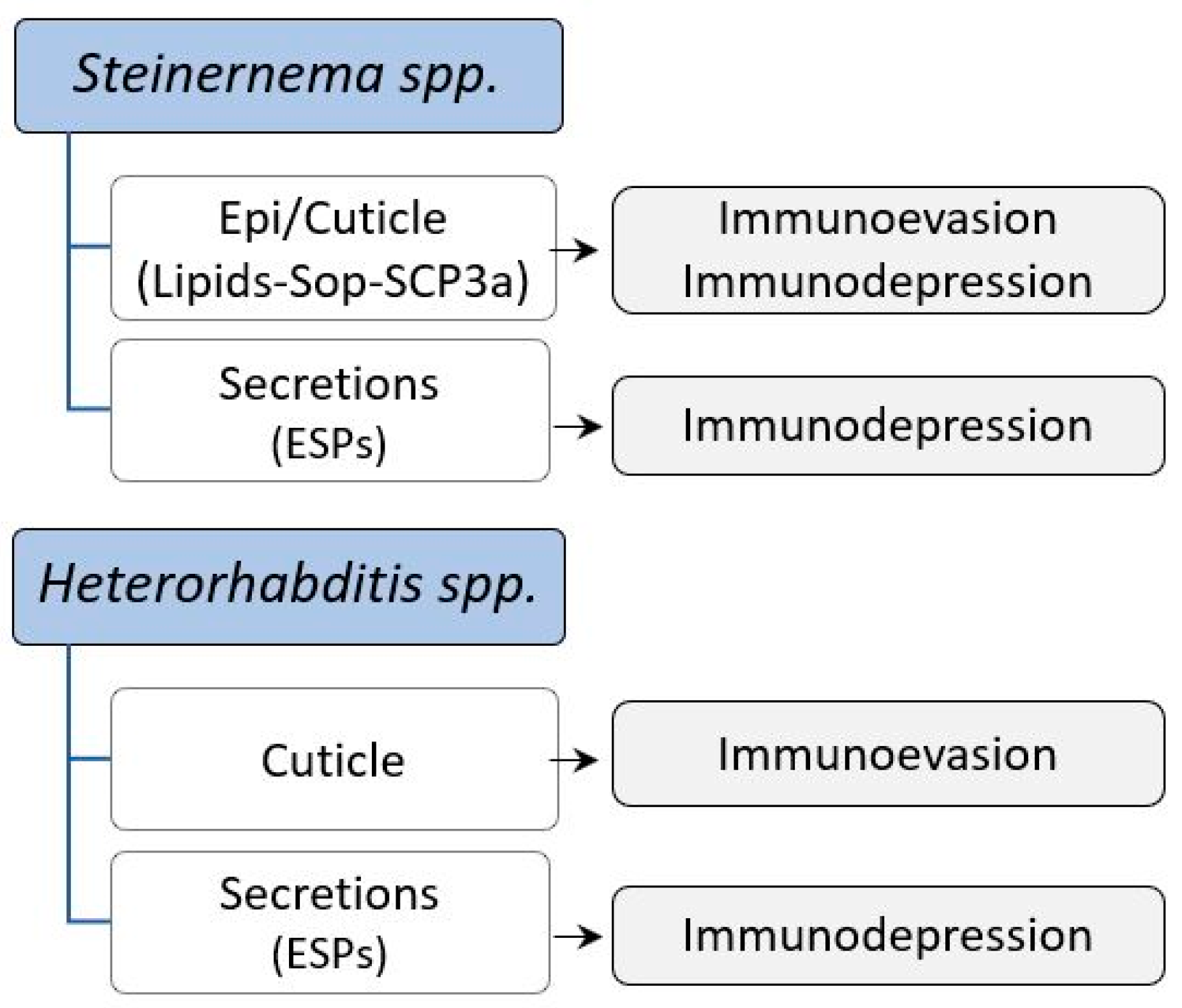
2.4. Entomopathogenic Nematocomplexes
3. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Loker, E.S. On Being a Parasite in an Invertebrate Host: A Short Survival Course. J. Parasitol. 1994, 80, 728–747. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, T.P.; Vasta, G.R. Parasite-Invertebrate Host Immune Interactions. In Invertebrate Immune Responses. Advances in Comparative and Environmental Physiology; Cooper, E.L., Ed.; Springer: Berlin/Heidelberg, Germany, 1996; Volume 24. [Google Scholar]
- Richman, A.; Kafatos, F.C. Immunity to eukaryotic parasites in vector insects. Curr. Opin. Immunol. 1995, 8, 14–19. [Google Scholar] [CrossRef]
- Ratcliffe, N.; Whitten, M. Vector immunity. In Microbe-vector Interactions in Vector-borne Diseases (Society for General Microbiology Symposia; Gillespie, S., Smith, G., Osbourn, A., Eds.; Cambridge University Press: Cambridge, UK, 2004; pp. 199–262. [Google Scholar]
- Sun, S.C.; Lindström, I.; Boman, H.G.; Faye, I.; Schmidt, O. Hemolin: An insect-immune protein belonging to the immunoglobulin superfamily. Science 1990, 250, 1729–1732. [Google Scholar] [CrossRef] [PubMed]
- Lanz-Mendoza, H.; Bettencourt, R.; Fabbri, M.; Faye, I. Regulation of the insect immune response: The effect of hemolin on cellular immune mechanisms. Cell. Immunol. 1996, 169, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Leclerc, V.; Reichhart, J.M. The immune response of Drosophila melanogaster. Immunol. Rev. 2004, 198, 59–71. [Google Scholar] [CrossRef] [PubMed]
- Lemaitre, B.; Hoffmann, J. The Host Defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef] [Green Version]
- Gotz, P.; Boman, H.G. Insect immunity. In Comprehensive Insect Physiology, Biochemistry and Pharmacology; Kerkut, G.A., Gilbert, L.I., Eds.; Pergamon Press: Oxford, UK, 1985; Volume III, pp. 453–485. [Google Scholar]
- Cociancich, S.; Bulet, P.; Hetru, C.; Hoffmann, J.A. The inducible antibacterial peptides of insects. Parasitol. Today 1994, 10, 132–139. [Google Scholar] [CrossRef]
- Gillespie, J.P.; Kanost, M.R.; Trenczek, T. Biological mediators of insect immunity. Annu. Rev. Entomol. 1997, 42, 611–643. [Google Scholar] [CrossRef]
- Hoffmann, J.A. The Immune Response of Drosophila. Nature 2003, 426, 33–38. [Google Scholar] [CrossRef]
- He, X.; Cao, X.; He, Y.; Bhattarai, K.; Rogers, J.; Hartson, S.; Jiang, H. Hemolymph proteins of Anopheles gambiae larvae infected by Escherichia coli. Dev. Comp. Immunol. 2017, 74, 110–124. [Google Scholar] [CrossRef]
- Medzhitov, R.; Janeway, C.A., Jr. Innate immunity: The virtues of a nonclonal system of recognition. Cell 1997, 91, 295–298. [Google Scholar] [CrossRef] [Green Version]
- Medzhitov, R.; Janeway, C.A., Jr. Decoding the patterns of self and nonself by the innate immune system. Science 2002, 296, 298–300. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.Q.; Kanost, M.R. Immulectin-2, a pattern recognition receptor that stimulates hemocytes encapsulation and melanization in the tobacco hornworm, Manduca sexta. Dev. Comp. Immunol. 2004, 28, 891–900. [Google Scholar] [CrossRef] [PubMed]
- Choe, K.M.; Lee, H.; Anderson, K.V. Drosophila peptidoglycan recognition protein LC (PGRP-LC) acts as a signal-transducing innate immune receptor. Proc. Natl. Acad. Sci. USA 2005, 102, 1122–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Chen, T.; Rayaprolu, S.; Zou, Z.; Xia, Q.; Xiang, Z.; Jiang, H. Proteolytic activation of pro-spätzle is required for the induced transcription of antimicrobial peptide genes in lepidopteran insects. Dev. Comp. Immunol. 2007, 31, 1002–1012. [Google Scholar] [CrossRef] [Green Version]
- Stokes, B.A.; Yadav, S.; Shokal, U.; Smith, L.C.; Eleftherianos, I. Bacterial and fungal pattern recognition receptors in homologous innate signaling pathways of insects and mammals. Front. Microbiol. 2015, 6, 19. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, M.; Li, C.F.; He, Z.; Lu, Y.; Liu, X.S.; Wang, Y.F.; Ip, Y.T.; Strand, M.R.; Yu, X.Q. Toll family members bind multiple Spätzle proteins and activate antimicrobial peptide gene expression in Drosophila. J. Biol. Chem. 2019, 294, 10172–10181. [Google Scholar] [CrossRef]
- Jomori, T.; Natori, S. Function of the lipopolysaccharide-binding protein of Periplaneta americana as an opsonin. FEBS Lett. 1992, 296, 283–286. [Google Scholar] [CrossRef] [Green Version]
- Boman, H.G. Antibacterial Peptides: Basic Facts and Emerging Concepts. J. Intern. Med. 2003, 254, 197–215. [Google Scholar] [CrossRef]
- Cerenius, L.; Soderhall, K. The Prophenoloxidase-Activating System in Invertebrates. Immunol. Rev. 2004, 198, 116–126. [Google Scholar] [CrossRef]
- Kanost, M.R.; Gorman, M.J. Phenoloxidase in insect immunity. In Insect Immunology, 1st ed.; Beckage, N.E., Ed.; Academic Press: Cambridge, MA, USA, 2008; pp. 69–96. [Google Scholar]
- De Lerma Barbaro, A.; Gariboldi, M.B.; Mastore, M.; Brivio, M.F.; Giovannardi, S. In Vivo Effects of a pro-PO system Inhibitor on the phagocytosis of Xenorhabdus nematophila in Galleria mellonella Larvae. Insects 2019, 10, 263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hultmark, D. Insect lysozymes. EXS 1996, 75, 87–102. [Google Scholar]
- Bulet, P.; Stöcklin, R.; Menin, L. Anti-microbial peptides: From invertebrates to vertebrates. Immunol. Rev. 2004, 198, 169–184. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.A.; Lemaitre, B. New insights on Drosophila antimicrobial peptide function in host defense and beyond. Curr. Opin. Immunol. 2020, 62, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Götz, P. Encapsulation in arthropods. In Immunity in Invertebrates; Brehélin, M., Ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA; Tokyo, Japan, 1986; pp. 153–170. [Google Scholar]
- Dunn, P.E. Insect antibacterial proteins. In Phylogenesis of Immune Functions; Warr, G.W., Cohen, N., Eds.; CRC Press: Boca Raton, FL, USA, 1991; pp. 19–44. [Google Scholar]
- Lavine, M.D.; Strand, M.R. Insect hemocytes and their role in immunity. Insect Biochem. Mol. Biol. 2002, 32, 1295–1309. [Google Scholar]
- Dubovskiy, I.M.; Kryukova, N.A.; Glupov, V.V.; Ratcliffe, N.A. Encapsulation and nodulation in insects. Invertebr. Surviv. J. 2016, 13, 229–246. [Google Scholar]
- Ryu, J.H.; Ha, E.M.; Oh, C.T.; Seol, J.H.; Brey, P.T.; Jin, I.; Lee, D.G.; Kim, J.; Lee, D.; Lee, W.J. An essential complementary role of NF-kappaB pathway to microbicidal oxidants in Drosophila gut immunity. EMBO J. 2006, 25, 3693–3701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tzou, P.; Ohresser, S.; Ferrandon, D.; Capovilla, M.; Reichhart, J.M.; Lemaitre, B.; Hoffmann, J.A.; Imler, J.L. Tissue-specific inducible expression of antimicrobial peptide genes in Drosophila surface epithelia. Immunity 2000, 13, 737–748. [Google Scholar] [CrossRef] [Green Version]
- Hurford, A.; Day, T. Immune evasion and the evolution of molecular mimicry in parasites. Evolution 2013, 67, 2889–2904. [Google Scholar] [CrossRef] [PubMed]
- Lie, J.K.; Heyneman, D.; Jeong, K.H. Studies on resistance in snails. 4. Induction of ventricular capsules and changes in the amebocyte-producing organ during sensitization of Biomphalaria glabrata snails. J. Parasitol. 1976, 62, 286–291. [Google Scholar] [CrossRef]
- Schmid-Hempel, P. Parasite immune evasion: A momentous molecular war. Trends Ecol. Evol. 2008, 23, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Smithers, S.R.; Terry, R.J.; Hockley, D.J. Host antigens in schistosomiasis. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1969, 171, 483–494. [Google Scholar]
- Damian, R.T. Tropomyosin and molecular mimicry. Parasitol. Today 1991, 7, 96. [Google Scholar] [CrossRef]
- Vellupilai, P.; Harn, D.A. Oligosaccharide specific induction of interleukin to production by B 220 cells from schistosome-infected mice: A mechanism for regulation of CD4+ cell subsets. Proc. Natl. Acad. Sci. USA 1994, 91, 18–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayne, C.J.; Boswell, C.A.; Yui, M.A. Widespread antigenic cross-reactivity between plasma proteins of a gastropod and its trematode parasite. Dev. Comp. Immunol. 1987, 11, 321–329. [Google Scholar] [CrossRef]
- Weston, D.S.; Kemp, W.M. Schistosoma mansoni—Comparison of cloned tropomyosin antigens shared between adult parasites and Biomphalaria glabrata. Exp. Parasitol. 1993, 76, 358–370. [Google Scholar] [CrossRef] [PubMed]
- Ratcliffe, N.A.; Rowley, A.F.; Fitzgerald, S.W.; Rhodes, C.P. Invertebrate immunity: Basic concepts and recent advances. Int. Rev. Cytol. 1985, 97, 183–350. [Google Scholar]
- Strand, M.R.; Pech, L.L. Immunological basis for compatibility in parasitoid-host relationships. Annu. Rev. Entomol. 1995, 40, 31–56. [Google Scholar] [CrossRef]
- Yoshino, T.P.; Bayne, C.J. Mimicry of snail host antigens by miracidia and primary sporocysts of Schistosoma mansoni. Parasite Immunol. 1983, 5, 317–328. [Google Scholar] [CrossRef]
- Dunn, T.S.; Yoshino, T.P. Tegumental surface modulation in Schistosoma mansoni primary sporocysts in response to ligand binding. Parasite Immunol. 1991, 13, 121–135. [Google Scholar] [CrossRef]
- Johnston, L.A.; Yoshino, T.P. Analysis of lectin and snail plasma-binding glycopeptides associated with the tegumental surface of the primary sporocysts of Schistosoma mansoni. Parasitology 1996, 112, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Mastore, M.; Brivio, M.F. Cuticular surface lipids are responsible for disguise properties of an entomoparasite against host cellular responses. Dev. Comp. Immunol. 2008, 32, 1050–1062. [Google Scholar] [CrossRef] [PubMed]
- Brivio, M.F.; Mastore, M. Nematobacterial Complexes and Insect Hosts: Different Weapons for the Same War. Insects 2018, 9, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kathirithamby, J.; Ross, L.D.; Johnston, J.S. Masquerading as self? Endoparasitic Strepsiptera (Insecta) enclose themselves in host-derived epidermal bag. Proc. Natl. Acad. Sci. USA 2003, 100, 7655–7659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khafagi, W.E.; Hegazi, E.M. Is the host or the parasitoid in control? Effects of host age and temperature on pseudoparasitization by Microplitis rufiventris in Spodoptera littoralis. J. Insect Physiol. 2004, 50, 577–584. [Google Scholar] [CrossRef]
- Gardiner, E.; Strand, M. Hematopoiesis in larval Pseudoplusia includens and Spodoptera frugiperda. Arch. Insect Biochem. Physiol. 2000, 43, 147–164. [Google Scholar] [CrossRef]
- Beetz, S.; Holthusen, T.K.; Koolman, J.; Trenczek, T. Correlation of hemocyte counts with different developmental parameters during the last larval instar of the tobacco hornworm, Manduca sexta. Arch. Insect Biochem. Physiol. 2008, 67, 63–75. [Google Scholar] [CrossRef]
- Pereira, S.S.; Trindade, S.; De Niz, M.; Figueiredo, L.M. Tissue tropism in parasitic diseases. Open Biol. 2019, 9, 190036. [Google Scholar] [CrossRef] [Green Version]
- Libersat, F.; Maayan, K.; Stav, E. Mind Control: How Parasites Manipulate Cognitive Functions in Their Insect Hosts. Front. Psychol. 2018, 9, 572. [Google Scholar] [CrossRef]
- Rahatkhah, Z.; Karimi, J.; Ghadamyari, M.; Brivio, M.F. Immune defenses of Agriotes lineatus larvae against entomopathogenic nematodes. BioControl 2015, 60, 641–653. [Google Scholar] [CrossRef]
- Binda-Rossetti, S.; Mastore, M.; Protasoni, M.; Brivio, M.F. Effects of an entomopathogen nematode on the immune response of the insect pest red palm weevil: Focus on the host antimicrobial response. J. Invertebr. Pathol. 2016, 133, 110–119. [Google Scholar] [CrossRef]
- Mastore, M.; Binda Rossetti, S.; Giovannardi, S.; Scarì, G.; Brivio, M.F. Inducible factors with antimicrobial activity after immune challenge in the haemolymph of Red Palm Weevil (Insecta). Innate Immun. 2015, 21, 392–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garriga, A.; Mastore, M.; Morton, A.; Garcia-del-Pino, F.; Brivio, M.F. Immune response of Drosophila suzukii larvae to infection with the nematobacterial complex Steinernema carpocapsae-Xenorhabdus nematophila. Insects 2020, 11, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argôlo-Filho, R.C.; Loguercio, L.L. Bacillus thuringiensis Is an Environmental Pathogen and Host-Specificity Has Developed as an Adaptation to Human-Generated Ecological Niches. Insects 2013, 5, 62–91. [Google Scholar] [CrossRef] [Green Version]
- Gohar, M.; Gilois, N.; Graveline, R.; Garreau, C.; Sanchis, V.; Lereclus, D. A comparative study of Bacillus cereus, Bacillus thuringiensis and Bacillus anthracis extracellular proteomes. Proteomics 2005, 5, 3696–3711. [Google Scholar] [CrossRef] [PubMed]
- Han, C.S.; Xie, G.; Challacombe, J.F.; Altherr, M.R.; Bhotika, S.S.; Brown, N.; Bruce, D.; Campbell, C.S.; Campbell, M.L.; Chen, J.; et al. Pathogenomic sequence analysis of Bacillus cereus and Bacillus thuringiensis isolates closely related to Bacillus anthracis. J. Bacteriol. 2006, 188, 3382–3390. [Google Scholar] [CrossRef] [Green Version]
- Park, S.H.; Kim, H.J.; Kim, J.H.; Kim, T.W.; Kim, H.Y. Simultaneous detection and identification of Bacillus cereus group bacteria using multiplex PCR. J. Microbiol. Biotechnol. 2007, 17, 1177–1182. [Google Scholar]
- Aronson, A.I.; Beckman, W.; Dunn, P. Bacillus thuringiensis and related insect pathogens. Microbiol. Rev. 1986, 50, 1–24. [Google Scholar] [CrossRef]
- Engel, P.; Moran, N.A. The gut microbiota of insects - diversity in structure and function. FEMS Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef]
- Peterson, J.W. Bacterial pathogenesis. In Medical Microbiology; Baron, S., Ed.; University of Texas Medical Branch at Galveston: Galveston, TX, USA, 1996; Chapter 7. [Google Scholar]
- Vallet-Gely, I.; Lemaitre, B.; Boccard, F. Bacterial strategies to overcome insect defences. Nat. Rev. Microbiol. 2008, 6, 302–313. [Google Scholar] [CrossRef] [Green Version]
- Los, F.C.; Randis, T.M.; Aroian, R.V.; Ratner, A.J. Role of pore-forming toxins in bacterial infectious diseases. Microbiol. Mol. Biol. Rev. 2013, 77, 173–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliva, C.; Turnbough, C.L., Jr.; Kearney, J.F. CD14-Mac-1 interactions in Bacillus anthracis spore internalization by macrophages. Proc. Natl. Acad. Sci. USA 2009, 106, 13957–13962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, Q.; Kao, G.; Qu, N.; Zhang, J.; Li, J.; Song, F. The Regulation of Exosporium-Related Genes in Bacillus thuringiensis. Sci. Rep. 2016, 6, 19005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bravo, A.; Gómez, I.; Porta, H.; García-Gómez, B.I.; Rodriguez-Almazan, C.; Pardo, L.; Soberón, M. Evolution of Bacillus thuringiensis Cry toxins insecticidal activity. Microb. Biotechnol. 2013, 6, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y. Detection and Mechanisms of Resistance Evolved in Insects to Cry Toxins from Bacillus thuringiensis. In Insect Midgut and Insecticidal Proteins, Advances in Insect Physiology; Dhadialla, T.S., Gill, S.S., Eds.; Academic Press: Cambridge, MA, USA, 2014; Chapter Six; pp. 297–342. [Google Scholar]
- Henriques, A.O.; Moran, C.P., Jr. Structure, assembly and function of the spore surface layers. Annu. Rev. Microbiol. 2007, 61, 555–588. [Google Scholar] [CrossRef]
- Setlow, P. Germination of spores of Bacillus species: What we know and do not know. J. Bacteriol. 2014, 196, 1297–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giorno, R.; Mallozzi, M.; Bozue, J.; Moody, K.S.; Slack, A.; Qiu, D.; Wang, R.; Friedlander, A.; Welkos, S.; Driks, A. Localization and assembly of proteins comprising the outer structures of the Bacillus anthracis spore. Microbiology 2009, 155, 1133–1145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKenney, P.T.; Driks, A.; Eichenberger, P. The Bacillus subtilis endospore: Assembly and functions of the multilayered coat. Nat. Rev. Microbiol. 2013, 11, 33–44. [Google Scholar] [CrossRef]
- Stewart, G.C. The Exosporium Layer of Bacterial Spores: A Connection to the Environment and the Infected Host. Microbiol. Mol. Biol. Rev. 2015, 79, 437–457. [Google Scholar] [CrossRef] [Green Version]
- Weaver, J.; Kang, T.J.; Raines, K.W.; Cao, G.L.; Hibbs, S.; Tsai, P.; Baillie, L.; Rosen, G.M.; Cross, A.S. Protective role of Bacillus anthracis exosporium in macrophage-mediated killing by nitric oxide. Infect. Immun. 2007, 75, 3894–3901. [Google Scholar] [CrossRef] [Green Version]
- Beaman, T.C.; Pankratz, H.S.; Gerhardt, P. Ultrastructure of the exosporium and underlying inclusions in spores of Bacillus megaterium strains. J. Bacteriol. 1972, 109, 1198–1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steichen, C.T.; Kearney, J.F.; Turnbough, C.L., Jr. Characterization of the exosporium basal layer protein BxpB of Bacillus anthracis. J. Bacteriol. 2005, 187, 5868–5876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faille, C.; Lequette, Y.; Ronse, A.; Slomianny, C.; Garénaux, E.; Guerardel, Y. Morphology and physico-chemical properties of Bacillus spores surrounded or not with an exosporium: Consequences on their ability to adhere to stainless steel. Int. J. Food Microbiol. 2010, 143, 125–135. [Google Scholar] [CrossRef]
- Tan, L.; Turnbough, C.L., Jr. Sequence motifs and proteolytic cleavage of the collagen-like glycoprotein BclA required for its attachment to the exosporium of Bacillus anthracis. J. Bacteriol. 2010, 192, 1259–1268. [Google Scholar] [CrossRef] [Green Version]
- Thompson, B.M.; Hsieh, H.Y.; Spreng, K.A.; Stewart, G.C. The co-dependence of BxpB/ExsFA and BclA for proper incorporation into the exosporium of Bacillus anthracis. Mol. Microbiol. 2011, 79, 799–813. [Google Scholar] [CrossRef] [Green Version]
- Steichen, C.; Chen, P.; Kearney, J.F.; Turnbough, C.L., Jr. Identification of the immunodominant protein and other proteins of the Bacillus anthracis exosporium. J. Bacteriol. 2003, 185, 1903–1910. [Google Scholar] [CrossRef] [Green Version]
- Redmond, C.; Baillie, L.W.; Hibbs, S.; Moir, A.J.; Moir, A. Identification of proteins in the exosporium of Bacillus anthracis. Microbiology 2004, 150, 355–363. [Google Scholar] [CrossRef] [Green Version]
- Thompson, B.M.; Stewart, G.C. Targeting of the BclA and BclB proteins to the Bacillus anthracis spore surface. Mol. Microbiol. 2008, 70, 421–434. [Google Scholar] [CrossRef]
- Thompson, B.M.; Hoelscher, B.C.; Driks, A.; Stewart, G.C. Assembly of the BclB glycoprotein into the exosporium and evidence for its role in the formation of the exosporium ‘cap’ structure in Bacillus anthracis. Mol. Microbiol. 2012, 86, 1073–1084. [Google Scholar] [CrossRef] [Green Version]
- Sylvestre, P.; Couture-Tosi, E.; Mock, M. Contribution of ExsFA and ExsFB proteins to the localization of BclA on the spore surface and to the stability of the bacillus anthracis exosporium. J. Bacteriol. 2005, 187, 5122–5128. [Google Scholar] [CrossRef] [Green Version]
- Fazzini, M.M.; Schuch, R.; Fischetti, V.A. A novel spore protein, ExsM, regulates formation of the exosporium in Bacillus cereus and Bacillus anthracis and affects spore size and shape. J. Bacteriol. 2010, 192, 4012–4021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodenburg, C.M.; McPherson, S.A.; Turnbough, C.L., Jr.; Dokland, T. Cryo-EM analysis of the organization of BclA and BxpB in the Bacillus anthracis exosporium. J. Struct. Biol. 2014, 186, 181–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boydston, J.A.; Chen, P.; Steichen, C.T.; Turnbough, C.L., Jr. Orientation within the exosporium and structural stability of the collagen-like glycoprotein BclA of Bacillus anthracis. J. Bacteriol. 2005, 187, 5310–5317. [Google Scholar] [CrossRef] [Green Version]
- Steichen, C.T.; Kearney, J.F.; Turnbough, C.L., Jr. Non-uniform assembly of the Bacillus anthracis exosporium and a bottle cap model for spore germination and outgrowth. Mol. Microbiol. 2007, 64, 359–367. [Google Scholar] [CrossRef]
- Cybulski, R.J., Jr.; Sanz, P.; Alem, F.; Stibitz, S.; Bull, R.L.; O’Brien, A.D. Four superoxide dismutases contribute to Bacillus anthracis virulence and provide spores with redundant protection from oxidative stress. Infect. Immun. 2009, 77, 274–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey-Smith, K.; Todd, S.J.; Southworth, T.W.; Proctor, J.; Moir, A. The ExsA protein of Bacillus cereus is required for assembly of coat and exosporium onto the spore surface. J. Bacteriol. 2005, 187, 3800–3806. [Google Scholar] [CrossRef] [Green Version]
- Grizanova, E.V.; Dubovskiy, I.M.; Whitten, M.M.; Glupov, V.V. Contributions of cellular and humoral immunity of Galleria mellonella larvae in defence against oral infection by Bacillus thuringiensis. J. Invertebr. Pathol. 2014, 119, 40–46. [Google Scholar] [CrossRef]
- Contreras, E.; Benito-Jardón, M.; López-Galiano, M.J.; Real, M.D.; Rausell, C. (Tribolium castaneum immune defense genes are differentially expressed in response to Bacillus thuringiensis toxins sharing common receptor molecules and exhibiting disparate toxicity. Dev. Comp. Immunol. 2015, 50, 139–145. [Google Scholar] [CrossRef]
- Li, S.; Xu, X.; Shakeel, M.; Xu, J.; Zheng, Z.; Zheng, J.; Yu, X.; Zhao, Q.; Jin, F. Bacillus thuringiensis Suppresses the Humoral Immune System to Overcome Defense Mechanism of Plutella xylostella. Front. Physiol. 2018, 9, 1478. [Google Scholar] [CrossRef] [PubMed]
- Hillyer, J.F. Insect immunology and hematopoiesis. Dev. Comp. Immunol. 2016, 58, 102–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, J.; Jiang, H.; Kanost, M.R.; Wang, Y. Serine proteases and their homologs in the Drosophila melanogaster genome: An initial analysis of sequence conservation and phylogenetic relationships. Gene 2003, 304, 117–131. [Google Scholar] [CrossRef]
- Crava, C.M.; Jakubowska, A.K.; Escriche, B.; Herrero, S.; Bel, Y. Dissimilar Regulation of Antimicrobial Proteins in the Midgut of Spodoptera exigua Larvae Challenged with Bacillus thuringiensis Toxins or Baculovirus. PLoS ONE 2015, 10, e0125991. [Google Scholar] [CrossRef]
- Faria, M.; Wraight, S. Mycoinsecticides and Mycoacaricides: A comprehensive list with worldwide coverage and international classification of formulation types. Biol. Control 2007, 43, 237–256. [Google Scholar] [CrossRef]
- Farenhorst, M.; Mouatcho, J.C.; Kikankie, C.K.; Brooke, B.D.; Hunt, R.H.; Thomas, M.B.; Koekemoer, L.L.; Knols, B.G.; Coetzee, M. Fungal infection counters insecticide resistance in African malaria mosquitoes. Proc. Natl. Acad. Sci. USA 2009, 106, 17443–17447. [Google Scholar] [CrossRef] [Green Version]
- Knols, B.G.; Bukhari, T.; Farenhorst, M. Entomopathogenic fungi as the next-generation control agents against malaria mosquitoes. Future Microbiol. 2010, 5, 339–341. [Google Scholar] [CrossRef]
- Butt, T.M.; Coates, C.J.; Dubovskiy, I.M.; Ratcliffe, N.A. Entomopathogenic Fungi: New Insights into Host-Pathogen Interactions. Adv. Genet. 2016, 94, 307–364. [Google Scholar]
- Qu, S.; Wang, S. Interaction of entomopathogenic fungi with the host immune system. Dev. Comp. Immunol. 2018, 83, 96–103. [Google Scholar] [CrossRef]
- Wanchoo, A.; Lewis, M.W.; Keyhani, N.O. Lectin mapping reveals stage-specific display of surface carbohydrates in in vitro and haemolymph-derived cells of the entomopathogenic fungus Beauveria bassiana. Microbiology 2009, 155, 3121–3133. [Google Scholar] [CrossRef] [Green Version]
- Pendland, J.C.; Boucias, D.G. Variations in the ability of galactose and mannose-specific lectins to bind to cell wall surfaces during growth of the insect pathogenic fungus Paecilomyces farinosus. Eur. J. Cell. Biol. 1993, 60, 322–330. [Google Scholar]
- Pendland, J.C.; Hung, S.Y.; Boucias, D.G. Evasion of host defense by in vivo-produced protoplast-like cells of the insect mycopathogen Beauveria bassiana. J. Bacteriol. 1993, 175, 5962–5969. [Google Scholar] [CrossRef] [Green Version]
- Hou, R.F.; Chang, J. Cellular defense response to Beauveria bassiana in the silkworm, Bombyx mori. Appl. Entomol. Zool. 1985, 20, 118–125. [Google Scholar] [CrossRef] [Green Version]
- Bidochka, M.J.; Khachatourians, G.G. Hemocytic defense response to the entomopathogenic fungus Beauveria bassiana in the migratory grasshopper Melanoplus sanguinipes. Entomol. Exp. Appl. 1987, 45, 151–156. [Google Scholar] [CrossRef]
- Yang, Z.; Jiang, H.; Zhao, X.; Lu, Z.; Luo, Z.; Li, X.; Zhao, J.; Zhang, Y. Correlation of cell surface proteins of distinct Beauveria bassiana cell types and adaption to varied environment and interaction with the host insect. Fungal Genet. Biol. 2017, 99, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Vertyporokh, L.; Hułas-Stasiak, M.; Wojda, I. Host-pathogen interaction after infection of Galleria mellonella with the filamentous fungus Beauveria bassiana. Insect Sci. 2019. [Google Scholar] [CrossRef]
- Li, J.; Ying, S.H.; Shan, L.T.; Feng, M.G. A new non-hydrophobic cell wall protein (CWP10) of Metarhizium anisopliae enhances conidial hydrophobicity when expressed in Beauveria bassiana. Appl. Microbiol. Biotechnol. 2010, 85, 975–984. [Google Scholar] [CrossRef]
- Ying, S.H.; Feng, M.G. A conidial protein (CP15) of Beauveria bassiana contributes to the conidial tolerance of the entomopathogenic fungus to thermal and oxidative stresses. Appl. Microbiol. Biotechnol. 2011, 90, 1711–1720. [Google Scholar] [CrossRef]
- Wang, C.; St Leger, R.J. The MAD1 adhesin of Metarhizium anisopliae links adhesion with blastospore production and virulence to insects, and the MAD2 adhesin enables attachment to plants. Eukaryot. Cell 2007, 6, 808–816. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; St Leger, R.J. A collagenous protective coat enables Metarhizium anisopliae to evade insect immune responses. Proc. Natl. Acad. Sci. USA 2006, 103, 6647–6652. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.L.; St Leger, R.J. Insect Immunity to Entomopathogenic Fungi. Adv. Genet. 2016, 94, 251–285. [Google Scholar]
- Wang, C.; Wang, S. Insect Pathogenic Fungi: Genomics, Molecular Interactions, and Genetic Improvements. Annu. Rev. Entomol. 2017, 62, 73–90. [Google Scholar] [CrossRef]
- Gibson, D.M.; Donzelli, B.G.; Krasnoff, S.B.; Keyhani, N.O. Discovering the secondary metabolite potential encoded within entomopathogenic fungi. Nat. Prod. Rep. 2014, 31, 1287–1305. [Google Scholar] [CrossRef] [PubMed]
- Molnár, I.; Gibson, D.M.; Krasnoff, S.B. Secondary metabolites from entomopathogenic Hypocrealean fungi. Nat. Prod. Rep. 2010, 27, 1241–1275. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Kang, Q.; Lu, Y.; Bai, L.; Wang, C. Unveiling the biosynthetic puzzle of destruxins in Metarhizium species. Proc. Natl. Acad. Sci. USA 2012, 109, 1287–1292. [Google Scholar] [CrossRef] [Green Version]
- Amiri-Besheli, B.; Khambay, B.; Cameron, S.; Deadman, M.L.; Butt, T.M. Inter- and intra-specific variation in destruxin production by insect pathogenic Metarhizium spp., and its significance to pathogenesis. Mycol. Res. 2000, 104, 447–452. [Google Scholar] [CrossRef]
- Kershaw, M.J.; Moorhouse, E.R.; Bateman, R.; Reynolds, S.E.; Charnley, A.K. The role of destruxins in the pathogenicity of metarhizium anisopliae for three species of insect. J. Invertebr. Pathol. 1999, 74, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhu, J.; Ying, S.H.; Feng, M.G. Three mitogen-activated protein kinases required for cell wall integrity contribute greatly to biocontrol potential of a fungal entomopathogen. PLoS ONE 2014, 9, e87948. [Google Scholar] [CrossRef] [PubMed]
- Feng, P.; Shang, Y.; Cen, K.; Wang, C. Fungal biosynthesis of the bibenzoquinone oosporein to evade insect immunity. Proc. Natl. Acad. Sci. USA 2015, 112, 11365–11370. [Google Scholar] [CrossRef] [Green Version]
- Lu, D.; Macchietto, M.; Chang, D.; Barros, M.M.; Baldwin, J.; Mortazavi, A.; Dillman, A.R. Activated entomopathogenic nematode infective juveniles release lethal venom proteins. PLoS Pathog. 2017, 13, e1006302. [Google Scholar] [CrossRef] [Green Version]
- Tzou, P.; De Gregorio, E.; Lemaitre, B. How Drosophila combats microbial infection: A model to study innate immunity and host-pathogen interactions. Curr. Opin. Microbiol. 2002, 5, 102–110. [Google Scholar] [CrossRef]
- Matskevich, A.A.; Quintin, J.; Ferrandon, D. The Drosophila PRR GNBP3 assembles effector complexes involved in antifungal defenses independently of its Toll-pathway activation function. Eur. J. Immunol. 2010, 40, 1244–1254. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, O.; Theopold, U.; Strand, M. Innate immunity and its evasion and suppression by hymenopteran endoparasitoids. Bioessays 2001, 23, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Pennacchio, F.; Strand, M.R. Evolution of developmental strategies in parasitic hymenoptera. Annu. Rev. Entomol. 2006, 51, 233–258. [Google Scholar] [CrossRef] [PubMed]
- Asgari, S.; Rivers, D.B. Venom proteins from endoparasitoid wasps and their role in host-parasite interactions. Annu. Rev. Entomol. 2011, 56, 313–335. [Google Scholar] [CrossRef] [PubMed]
- Falabella, P. The mechanism utilized by Toxoneuron nigriceps in inhibiting the host immune system. Invertebr. Surviv. J. 2018, 15, 240–255. [Google Scholar]
- Yin, C.; Li, M.; Hu, J.; Lang, K.; Chen, Q.; Liu, J.; Guo, D.; He, K.; Dong, Y.; Luo, J.; et al. The genomic features of parasitism, Polyembryony and immune evasion in the endoparasitic wasp Macrocentrus cingulum. BMC Genom. 2018, 19, 420. [Google Scholar] [CrossRef] [Green Version]
- Bedwin, O. An insect glycoprotein: A study of the particles responsible for the resistance of a parasitoid’s egg to the defence reactions of its insect host. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1979, 205, 271–286. [Google Scholar]
- Feddersen, I.; Sander, K.; Schmidt, O. Virus-like particles with host protein-like antigenic determinants protect an insect parasitoid from encapsulation. EXS 1986, 42, 1278–1281. [Google Scholar] [CrossRef]
- Morales, J.; Chiu, H.; Oo, T.; Plaza, R.; Hoskins, S.; Govind, S. Biogenesis, structure, and immune-suppressive effects of virus-like particles of a Drosophila parasitoid, Leptopilina victoriae. J. Insect Physiol. 2005, 51, 181–195. [Google Scholar] [CrossRef]
- Ye, X.Q.; Shi, M.; Huang, J.H.; Chen, X.X. Parasitoid polydnaviruses and immune interaction with secondary hosts. Dev. Comp. Immunol. 2018, 83, 124–129. [Google Scholar] [CrossRef]
- Beck, M.; Theopold, U.; Schmidt, O. Evidence for serine protease inhibitor activity in the ovarian calyx fluid of the endoparasitoid Venturia canescens. J. Insect Physiol. 2000, 46, 1275–1283. [Google Scholar] [CrossRef]
- Zhang, G.; Lu, Z.Q.; Jiang, H.; Asgari, S. Negative regulation of prophenoloxidase (proPO) activation by a clip-domain serine proteinase homolog (SPH) from endoparasitoid venom. Insect Biochem. Mol. Biol. 2004, 34, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Wan, B.; Goguet, E.; Ravallec, M.; Pierre, O.; Lemauf, S.; Volkoff, A.N.; Gatti, J.L.; Poirié, M. Venom Atypical Extracellular Vesicles as Interspecies Vehicles of Virulence Factors Involved in Host Specificity: The Case of a Drosophila Parasitoid Wasp. Front. Immunol. 2019, 10, 1688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotheram, S. Immune surface of eggs of a parasitic insect. Nature 1967, 214, 700. [Google Scholar] [CrossRef] [PubMed]
- Davies, D.H.; Vinson, S.B. Passive evasion by eggs of braconid parasitoid Cardiochiles nigriceps of encapsulation in vitro by haematocytes of host Heliothis virescens. Possible role for fibrous layer in immunity. J. Insect Physiol. 1986, 32, 1003. [Google Scholar] [CrossRef]
- Reineke, A.; Asgari, S.; Schmidt, O. Evolutionary origin of Venturia canescens virus-like particles. Arch. Insect Biochem. Physiol. 2006, 61, 123–133. [Google Scholar] [CrossRef]
- Hellers, M.; Beck, M.; Theopold, U.; Kamei, M.; Schmidt, O. Multiple alleles encoding a virus-like particle protein in the ichneumonid endoparasitoid Venturia canescens. Insect Mol. Biol. 1996, 5, 239–249. [Google Scholar] [CrossRef]
- Theopold, U.; Krause, E.; Schmidt, O. Cloning of a VLP-protein coding gene from a parasitoid wasp Venturia canescens. Arch. Insect Biochem. Physiol. 1994, 26, 137–145. [Google Scholar] [CrossRef]
- Asgari, S.; Reineke, A.; Beck, M.; Schmidt, O. Isolation and characterization of a neprilysin-like protein from Venturia canescens virus-like particles. Insect Mol. Biol. 2002, 11, 477–485. [Google Scholar] [CrossRef]
- Theopold, U.; Samakovlis, C.; Erdjument-Bromage, H.; Dillon, N.; Axelsson, B.; Schmidt, O.; Tempst, P.; Hultmark, D. Helix pomatia lectin, an inducer of Drosophila immune response, binds to hemomucin, a novel surface mucin. J. Biol. Chem. 1996, 271, 12708–12715. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Xu, Q.; Hu, S.; Yu, X.; Liang, Z.; Zhang, W. Hemomucin, an O-glycosylated protein on embryos of the wasp Macrocentrus cingulum that protects it against encapsulation by hemocytes of the host Ostrinia furnacalis. J. Innate Immun. 2014, 6, 663–675. [Google Scholar] [CrossRef]
- Coodin, S.; Caveney, S. Lipophorin inhibits adhesion of cockroach (Periplaneta americana) haemocytes in vitro. J. Insect Physiol. 1992, 38, 853–862. [Google Scholar] [CrossRef]
- Mandato, C.A.; Diehl-Jones, W.L.; Downer, R.G.H. Insect hemocyte adhesion in vitro inhibition by apoliphorin I and an artificial substrate. J. Insect Physiol. 1996, 42, 143–148. [Google Scholar] [CrossRef]
- Davies, D.H.; Burghardt, R.L.; Vinson, S.B. Oogenesis of Cardiochiles nigriceps viereck (Hymenoptera: Braconidae): Histochemistry and development of the chorion with special reference to the fibrous layer. Int. J. Insect Morphol. Embryol. 1986, 15, 363–374. [Google Scholar] [CrossRef]
- Asgari, S.; Schmidt, O. Passive protection of eggs from the parasitoid, Cotesia rubecula, in the host, Pieris rapae. J. Insect Physiol. 1994, 40, 789–795. [Google Scholar] [CrossRef]
- Teng, Z.; Wu, H.; Ye, X.; Xiong, S.; Xu, G.; Wang, F.; Fang, Q.; Ye, G. An Ovarian Protein Involved in Passive Avoidance of an Endoparasitoid to Evade Its Host Immune Response. J. Proteome Res. 2019, 18, 2695–2705. [Google Scholar] [CrossRef]
- Asgari, S.; Theopold, U.; Wellby, C.; Schmidt, O. A protein with protective properties against the cellular defense reactions in insects. Proc. Natl. Acad. Sci. USA 1998, 95, 3690–3695. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Matsumoto, H.; Hayakawa, Y. Detailed characterization of polydnavirus immunoevasive proteins in an endoparasitoid wasp. Eur. J. Biochem. 2002, 269, 2557–2566. [Google Scholar] [CrossRef]
- Furihata, S.; Tanaka, K.; Ryuda, M.; Ochiai, M.; Matsumoto, H.; Csikos, G.; Hayakawa, Y. Immunoevasive protein (IEP)-containing surface layer covering polydnavirus particles is essential for viral infection. J. Invertebr. Pathol. 2014, 115, 26–32. [Google Scholar] [CrossRef]
- Moreau, S.J.; Asgari, S. Venom Proteins from Parasitoid Wasps and Their Biological Functions. Toxins 2015, 7, 2385–2412. [Google Scholar] [CrossRef] [Green Version]
- Zhang, G.; Schmidt, O.; Asgari, S. A calreticulin-like protein from endoparasitoid venom fluid is involved in host hemocyte inactivation. Dev. Comp. Immunol. 2006, 30, 756–764. [Google Scholar] [CrossRef]
- Andrew, N.; Basio, M.; Kim, Y. Additive effect of teratocyte and calyx fluid from Cotesia plutellae on immunosuppression of Plutella xylostella. Physiol. Entomol. 2006, 31, 341–347. [Google Scholar] [CrossRef]
- Ali, M.R.; Seo, J.; Lee, D.; Kim, Y. Teratocyte-secreting proteins of an endoparasitoid wasp, Cotesia plutellae, prevent host metamorphosis by altering endocrine signals. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 166, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Strand, M.R. Teratocytes and their functions in parasitoids. Curr. Opin. Insect Sci. 2014, 6, 68–73. [Google Scholar] [CrossRef]
- Poinar, G.O., Jr. Origins and phylogenetic relationships of the entomophilic rhabditids Heterorhabditis and Steinernema. Fundam. Appl. Nematol. 1993, 16, 333–338. [Google Scholar]
- Poinar, G. Nematode biopesticides. Fundam. Appl. Nematol. 1998, 21, 733–737. [Google Scholar]
- Kaya, H.K.; Gaugler, R. Entomopathogenic nematodes. Annu. Rev. Entomol. 1993, 38, 181–206. [Google Scholar] [CrossRef]
- Schmid-Hempel, P. Immune defence, parasite evasion strategies and their relevance for macroscopic phenomena such as virulence. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2009, 364, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Castillo, J.C.; Reynolds, S.E.; Eleftherianos, I. Insect immune responses to nematode parasites. Trends Parasitol. 2011, 27, 537–547. [Google Scholar] [CrossRef]
- Ciche, T.A.; Darby, C.; Ehlers, R.U.; Forst, S.; Goodrich-Blaire, H. Dangerous liaisons: The symbiosis of entomopathogenic nematodes and bacteria. Biol. Control 2006, 38, 22–46. [Google Scholar] [CrossRef]
- Akhurst, R.J.; Dunphy, G.B. Tripartite interactions between symbiotically associated entomopathogenic bacteria, nematodes, and their insect hosts. In Parasites and Pathogens of Insects; Academic Press: Cambridge, MA, USA, 1993; pp. 1–23. [Google Scholar]
- Cooper, D.; Eleftherianos, I. Parasitic Nematode Immunomodulatory Strategies: Recent Advances and Perspectives. Pathogens 2016, 5, 58. [Google Scholar] [CrossRef] [Green Version]
- Blaxter, M.L.; Page, A.P.; Rudin, W.; Maizels, R.M. Nematode surface coats: Actively evading immunity. Parasitol. Today 1992, 8, 243–247. [Google Scholar] [CrossRef]
- Politz, S.M.; Philipp, M. Caenorhabditis elegans as a model for parasitic nematodes: A focus on the cuticle. Parasitol. Today 1992, 8, 6–12. [Google Scholar] [CrossRef]
- Cox, G.N.; Kusch, M.; Edgar, R.S. Cuticle of Caenorhabditis elegans: Its isolation and partial characterization. J. Cell Biol. 1981, 90, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Fetterer, R.H.; Rhoads, M.L. Biochemistry of the nematode cuticle: Relevance to parasitic nematodes of livestock. Vet. Parasitol. 1993, 46, 103–111. [Google Scholar] [CrossRef]
- Mastore, M.; Arizza, V.; Manachini, B.; Brivio, M.F. Modulation of immune responses of Rhynchophorus ferrugineus (Insecta: Coleoptera) induced by the entomopathogenic nematode Steinernema carpocapsae (Nematoda: Rhabditida). Insect Sci. 2015, 22, 748–760. [Google Scholar] [CrossRef]
- Brivio, M.F.; Mastore, M.; Moro, M. The role of Steinernema feltiae body-surface lipids in host–parasite immunological interactions. Mol. Biochem. Parasitol. 2004, 135, 111–121. [Google Scholar] [CrossRef]
- Brivio, M.F.; Toscano, A.; De Pasquale, S.M.; De Lerma Barbaro, A.; Giovannardi, S.; Finzi, G.; Mastore, M. Surface protein components from entomopathogenic nematodes and their symbiotic bacteria: Effects on immune responses of the greater wax moth, Galleria mellonella (Lepidoptera: Pyralidae). Pest Manag. Sci. 2018, 74, 2089–2099. [Google Scholar] [CrossRef]
- Wang, Y.; Gaugler, R. Steinernema glaseri Surface Coat Protein Suppresses the Immune Response of Popillia japonica (Coleoptera: Scarabaeidae) Larvae. Biol. Control 1999, 14, 45–50. [Google Scholar] [CrossRef]
- Toubarro, D.; Avila, M.M.; Montiel, R.; Simões, N. A pathogenic nematode targets recognition proteins to avoid insect defenses. PLoS ONE 2013, 8, e75691. [Google Scholar] [CrossRef] [Green Version]
- Balasubramanian, N.; Toubarro, D.; Simões, N. Biochemical study and in vitro insect immune suppression by a trypsin-like secreted protease from the nematode Steinernema carpocapsae. Parasite Immunol. 2010, 32, 165–175. [Google Scholar] [CrossRef]
- Chang, D.Z.; Serra, L.; Lu, D.; Mortazavi, A.; Dillman, A.R. A core set of venom proteins is released by entomopathogenic nematodes in the genus Steinernema. PLoS Pathog. 2019, 15, e1007626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, A.; Gouge, D.H.; Ehlers, R.; Hague, N. Avoidance of Encapsulation by Heterorhabditis spp. Infecting Larvae of Tipula oleracea. J. Invertebr. Pathol. 1997, 70, 161–164. [Google Scholar] [CrossRef] [PubMed]
- Armer, C.A.; Rao, S.; Berry, R.E. Insect cellular and chemical limitations to pathogen development: The Colorado potato beetle, the nematode Heterorhabditis marelatus, and its symbiotic bacteria. J. Invertebr. Pathol. 2004, 87, 114–122. [Google Scholar] [CrossRef] [PubMed]
- An, R.; Voss, M.; Jagdale, G.B.; Grewal, P.S. Differences in Immune Defense Evasion of Selected Inbred Lines of Heterorhabditis Bacteriophora in Two White Grub Species. Insects 2012, 3, 378–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebrahimi, L.; Niknam, G.; Dunphy, G.B. Hemocyte responses of the Colorado potato beetle, Leptinotarsa decemlineata, and the greater wax moth, Galleria mellonella, to the entomopathogenic nematodes, Steinernema feltiae and Heterorhabditis bacteriophora. J. Insect Sci. 2011, 11, 75. [Google Scholar] [CrossRef] [Green Version]
- Jarosz, J. Active resistance of entomophagous rhabditid Heterorhabditis bacteriophora to insect immunity. Parasitology 1998, 117, 201–208. [Google Scholar] [CrossRef]
- Kenney, E.; Hawdon, J.M.; O’Halloran, D.; Eleftherianos, I. Heterorhabditis bacteriophora Excreted-Secreted Products Enable Infection by Photorhabdus luminescens Through Suppression of the Imd Pathway. Front. Immunol. 2019, 10, 2372. [Google Scholar] [CrossRef]
- Richards, G.R.; Goodrich-Blair, H. Masters of conquest and pillage: Xenorhabdus nematophila global regulators control transitions from virulence to nutrient acquisition. Cell Microbiol. 2009, 11, 1025–1033. [Google Scholar] [CrossRef] [Green Version]
- Khandelwal, P.; Banerjee-Bhatnagar, N. Insecticidal activity associated with the outer membrane vesicles of Xenorhabdus nematophilus. Appl. Environ. Microbiol. 2003, 69, 2032–2037. [Google Scholar] [CrossRef] [Green Version]
- Ellis, T.N.; Kuehn, M.J. Virulence and immunomodulatory roles of bacterial outer membrane vesicles. Microbiol. Mol. Biol. Rev. 2010, 74, 81–94. [Google Scholar] [CrossRef] [Green Version]
- Givaudan, A.; Lanois, A. Flagellar Regulation and Virulence in the Entomopathogenic Bacteria—Xenorhabdus nematophila and Photorhabdus luminescens. In The Molecular Biology of Photorhabdus Bacteria. Current Topics in Microbiology and Immunology; Ffrench-Constant, R., Ed.; Springer: Cham, Switzerland, 2016; Volume 402. [Google Scholar]
- Darsouei, R.; Karimi, J.; Dunphy, G.B. Functional Characterization of Outer Membrane Proteins (OMPs) in Xenorhabdus nematophila and Photorhabdus luminescens through Insect Immune Defense Reactions. Insects 2019, 10, 352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunphy, G.B.; Webster, J.M. Antihemocytic Surface Components of Xenorhabdus Nematophilus Var. Dutki and Their Modification by Serum of Nonimmune Larvae of Galleria. J. Invertebr. Pathol. 1991, 58, 40–51. [Google Scholar] [CrossRef]
- Giannoulis, P.; Brooks, C.L.; Dunphy, G.B.; Niven, D.F.; Mandato, C.A. Surface antigens of Xenorhabdus nematophila (F. Enterobacteriaceae) and Bacillus subtilis (F. Bacillaceae) react with antibacterial factors of Malacosoma disstria (C. Insecta: O. Lepidoptera) hemolymph. J. Invertebr. Pathol. 2008, 97, 211–222. [Google Scholar] [CrossRef] [PubMed]
- Marokházi, J.; Lengyel, K.; Pekár, S.; Felföldi, G.; Patthy, A.; Gráf, L.; Fodor, A.; Venekei, I. Comparison of proteolytic activities produced by entomopathogenic Photorhabdus bacteria: Strain- and phase-dependent heterogeneity in composition and activity of four enzymes. Appl. Environ. Microbiol. 2004, 70, 7311–7320. [Google Scholar] [CrossRef] [Green Version]
- Goodrich-Blair, H.; Clarke, D.J. Mutualism and pathogenesis in Xenorhabdus and Photorhabdus: Two roads to the same destination. Mol. Microbiol. 2007, 64, 260–268. [Google Scholar] [CrossRef]
- Hinchliffe, S.J.; Hares, M.C.; Dowling, A.J.; Ffrench-Constant, R.H. Insecticidal Toxins from the Photorhabdus and Xenorhabdus Bacteria. Open Toxinol. J. 2010, 3, 83–100. [Google Scholar] [CrossRef]











© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brivio, M.F.; Mastore, M. When Appearance Misleads: The Role of the Entomopathogen Surface in the Relationship with Its Host. Insects 2020, 11, 387. https://doi.org/10.3390/insects11060387
Brivio MF, Mastore M. When Appearance Misleads: The Role of the Entomopathogen Surface in the Relationship with Its Host. Insects. 2020; 11(6):387. https://doi.org/10.3390/insects11060387
Chicago/Turabian StyleBrivio, Maurizio Francesco, and Maristella Mastore. 2020. "When Appearance Misleads: The Role of the Entomopathogen Surface in the Relationship with Its Host" Insects 11, no. 6: 387. https://doi.org/10.3390/insects11060387
APA StyleBrivio, M. F., & Mastore, M. (2020). When Appearance Misleads: The Role of the Entomopathogen Surface in the Relationship with Its Host. Insects, 11(6), 387. https://doi.org/10.3390/insects11060387