How Crucial is the Functional Pit Organ for the Varroa Mite?


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Material
2.2. Manipulation of Female Mites for Bioassays
2.2.1. Host Orientation Behavior
2.2.2. Host Identification and Feeding
2.2.3. Mite Reproduction
2.3. Data Analyses
3. Results
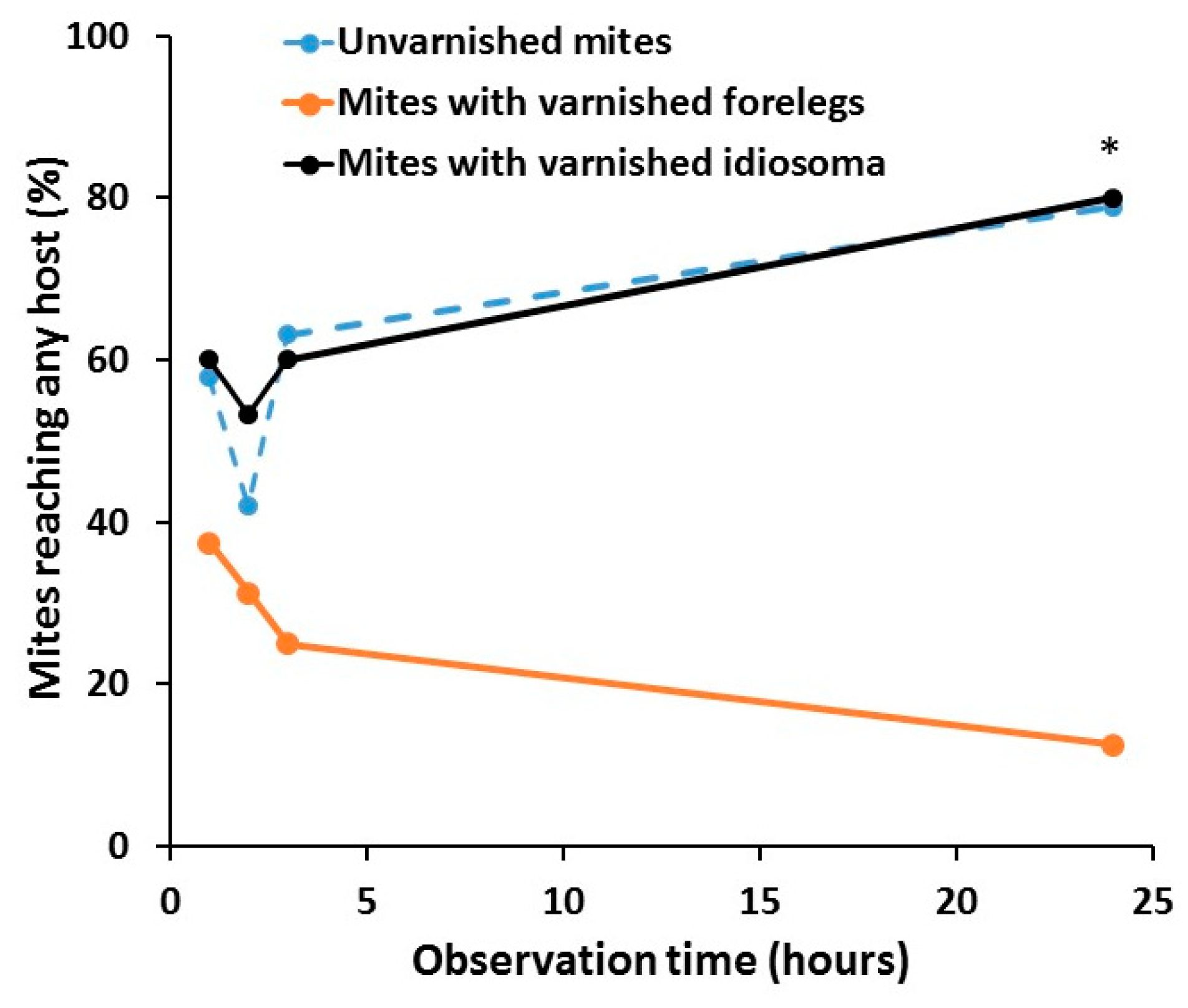
3.1. Host Orientation Behavior
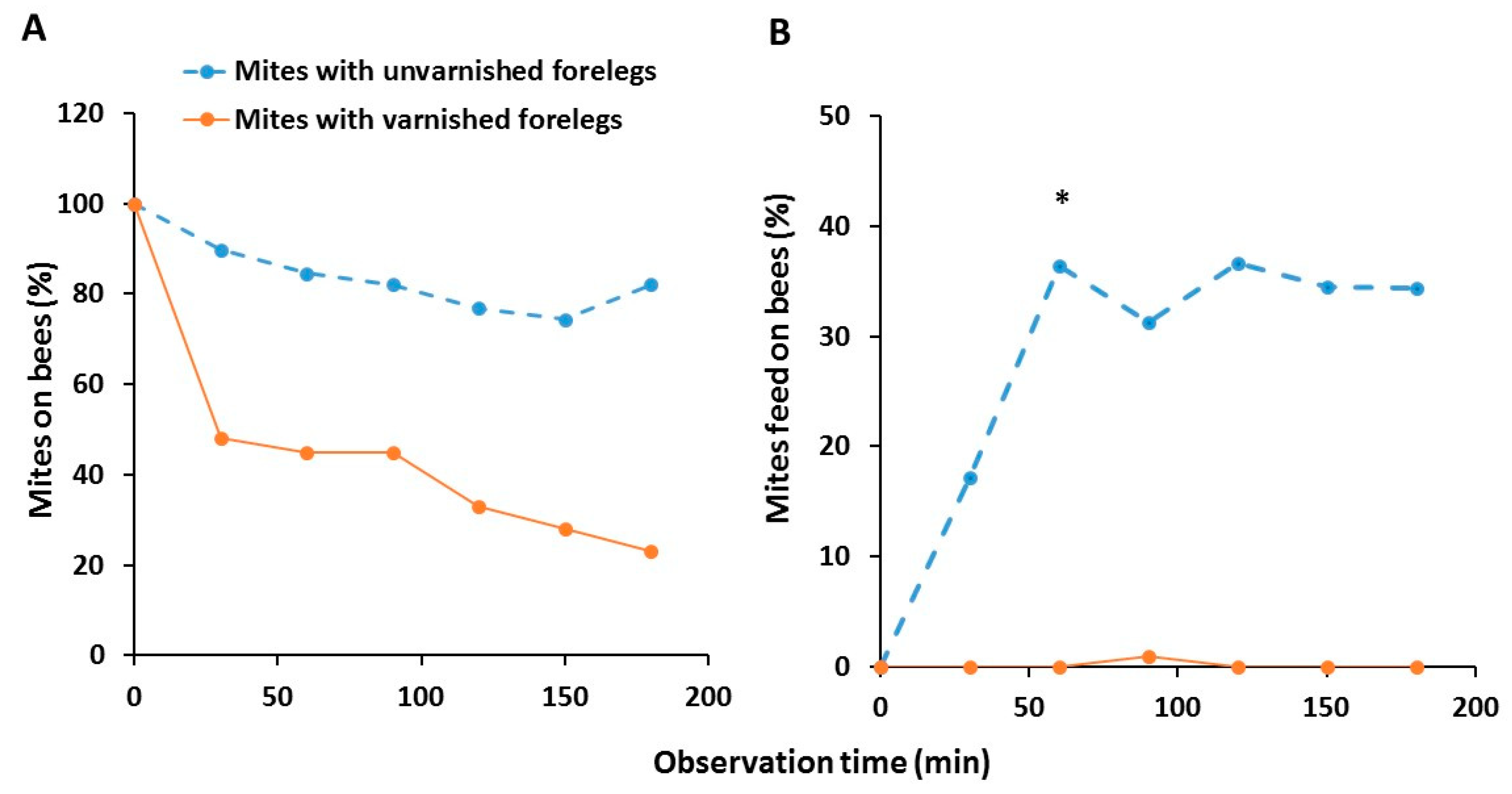
3.2. Host Identification and Feeding
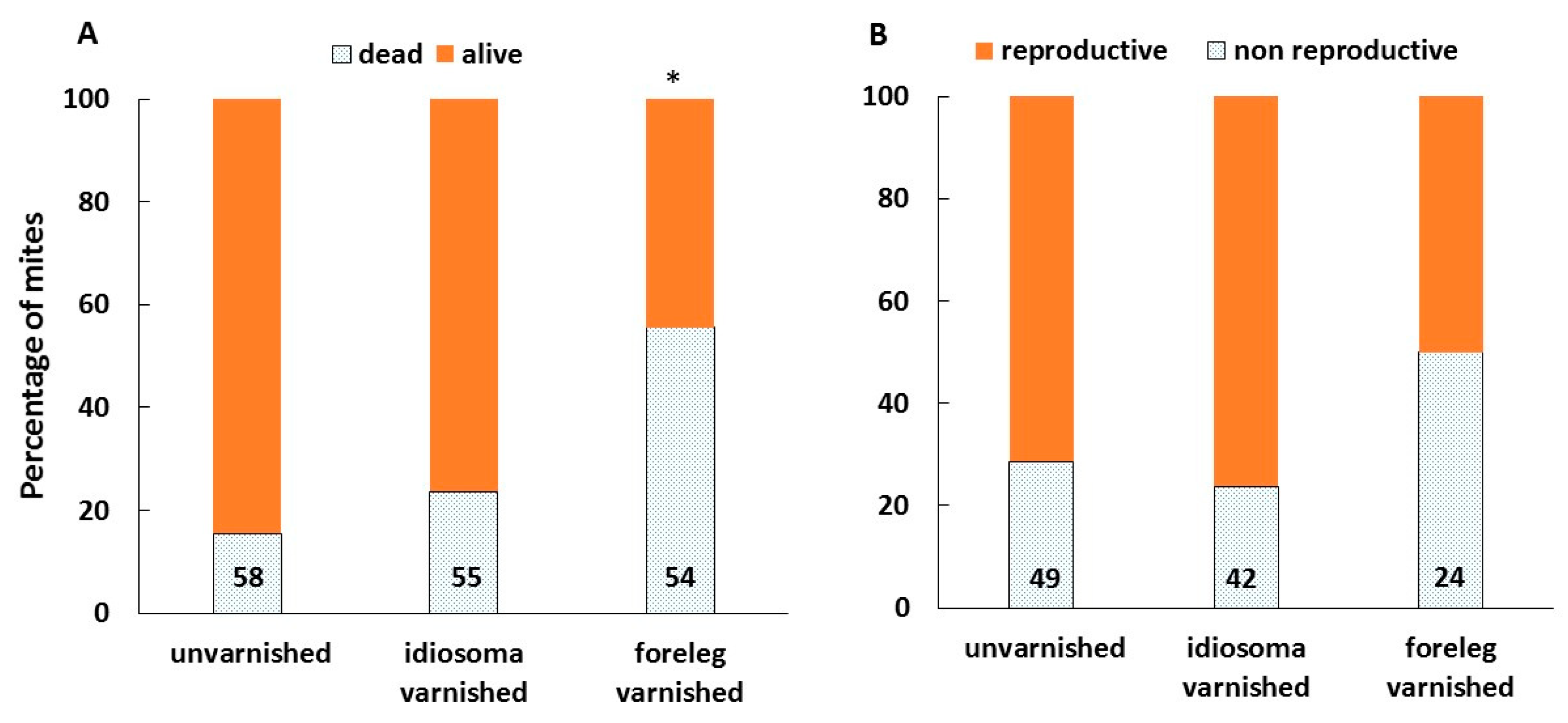
3.3. Mite Reproduction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Leal, W.S. Odorant Reception in Insects: Roles of Receptors, Binding Proteins, and Degrading Enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef] [PubMed]
- Carr, A.L.; Roe, M. Acarine Attractants: Chemoreception, Bioassay, Chemistry and Control. Pestic. Biochem. Physiol. 2016, 131, 60–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozaki, K.; Utoguchi, A.; Yamada, A.; Yoshikawa, H. Identification and Genomic Structure of Chemosensory Proteins (CSP) and Odorant Binding Proteins (OBP) Genes Expressed in Foreleg Tarsi of the Swallowtail Butterfly Papilio Xuthus. Insect Biochem. Mol. Biol. 2008, 38, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Francis, R.M.; Nielsen, S.L.; Kryger, P. Varroa-Virus Interaction in Collapsing Honey Bee Colonies. PLoS ONE 2013, 8, e57540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenkranz, P.; Aumeier, P.; Ziegelmann, B. Biology and Control of Varroa destructor. J. Invertebr. Pathol. 2010, 103, S96–S119. [Google Scholar] [CrossRef]
- Martin, S. Biology and Life History of Varroa Mites. In Mites of the Honey Bee; Webster, T.C., Delaplane, K.S., Eds.; Dadant & Sons: Hamilton, IL, USA, 2001. [Google Scholar]
- Pernal, S.F.; Baird, D.S.; Birmingham, A.L.; Higo, H.A.; Slessor, K.N.; Winston, M.L. Semiochemicals Influencing the Host-Finding Behaviour of Varroa destructor. Exp. Appl. Acarol. 2005, 37, 1–26. [Google Scholar] [CrossRef]
- Singh, N.K.; Eliash, N.; Stein, I.; Kamer, Y.; Ilia, Z. Identification and Gene-Silencing of a Putative Odorant Receptor Transcription Factor in Varroa destructor: Possible Role in Olfaction. Insect Mol. Biol. 2016, 25, 181–190. [Google Scholar] [CrossRef]
- Frey, E.; Odemer, R.; Blum, T.; Rosenkranz, P. Activation and Interruption of the Reproduction of Varroa destructor Is Triggered by Host Signals (Apis mellifera). J. Invertebr. Pathol. 2013, 113, 56–62. [Google Scholar] [CrossRef]
- Nazzi, F.; Le Conte, Y. Ecology of Varroa destructor, the Major Ectoparasite of the Western Honeybee, Apis mellifera. Annu. Rev. Entomol. 2016, 61, 417–432. [Google Scholar] [CrossRef] [Green Version]
- Plettner, E.; Eliash, N.; Singh, N.K.; Pinnelli, G.R.; Soroker, V. The Chemical Ecology of Host-Parasite Interaction as a Target of Varroa destructor Control Agents. Apidologie 2017, 48, 78–92. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.S.; Peng, T.P. Palpal Tarsal Sensilla of the Female Mite, Varroa jacobsoni Oudemans (Acari: Varroidae). Can. Entomol. 1990, 122, 295–300. [Google Scholar] [CrossRef]
- Dillier, F.X.; Fluri, P.; Imdorf, A. Review of the Orientation Behaviour in the Bee Parasitic Mite Varroa destructor: Sensory Equipment and Cell Invasion Behaviour. Rev. Suisse Zool. 2006, 113, 857–877. [Google Scholar] [CrossRef]
- Singh, N.K.; Eliash, N.; Kamer, Y.; Zaidman, I.; Plettner, E.; Soroker, V. The effect of DEET on Chemosensing of the Honey Bee and Its Parasite Varroa destructor. Apidologie 2015, 46, 380–391. [Google Scholar] [CrossRef]
- Eliash, N.; Singh, N.K.; Kamer, Y.; Pinnelli, G.R.; Plettner, E.; Soroker, V. Can We Disrupt the Sensing of Honey Bees by the Bee Parasite Varroa destructor? PLoS ONE 2014, 9, e106889. [Google Scholar] [CrossRef] [PubMed]
- Renthal, R.; Manghnani, L.; Bernal, S.; Qu, Y.; Griffith, W.P.; Lohmeyer, K.; Guerrero, F.D.; Borges, L.M.; Pérez de León, A. The Chemosensory Appendage Proteome of Amblyomma americanum (Acari: Ixodidae) Reveals Putative Odorant-Binding and Other Chemoreception-Related Proteins. Insect Sci. 2017, 24, 730–742. [Google Scholar] [CrossRef] [PubMed]
- Häußermann, C.K.; Ziegelmann, B.; Bergmann, P.; Rosenkranz, P. Male Mites (Varroa destructor) Perceive the Female Sex Pheromone with the Sensory Pit Organ On the Front Leg Tarsi. Apidologie 2015, 46, 771–778. [Google Scholar] [CrossRef] [Green Version]
- Ramsey, S.D.; Ochoa, R.; Bauchan, G.; Gulbronson, C.; Mowery, J.D.; Cohen, A.; Lim, D.; Joklik, J.; Cicero, J.M.; Ellis, J.D.; et al. Varroa destructor Feeds Primarily on Honey Bee Fat Body Tissue and Not Hemolymph. Proc. Natl. Acad. Sci. USA 2019, 116, 1792–1801. [Google Scholar] [CrossRef] [Green Version]
- Dietemann, V.; Nazzi, F.; Martin, S.J.; Anderson, D.L.; Locke, B.; Delaplane, K.S.; Wauquiez, Q.; Tannahill, C.; Frey, E.; Ziegelmann, B.; et al. Standard Methods for Varroa Research. J. Apic. Res. 2013, 52, 1–54. [Google Scholar] [CrossRef] [Green Version]
- Cabrera Cordon, A.R.; Shirk, P.D.; Duehl, A.J.; Evans, J.D.; Teal, P.E.A. Variable Induction of Vitellogenin Genes in the Varroa Mite, Varroa destructor (Anderson & Trueman), by the honeybee, Apis mellifera L., host and its environment. Insect Mol. Biol. 2013, 22, 88–103. [Google Scholar] [CrossRef]
- Finn, R.D.; Attwood, T.K.; Babbitt, P.C.; Bateman, A.; Bork, P.; Bridge, A.J.; Chang, H.Y.; Dosztányi, Z.; El-Gebali, S.; Fraser, M.; et al. InterPro in 2017-Beyond Protein Family and Domain Annotations. Nucleic Acids Res. 2017, 45, D190–D199. [Google Scholar] [CrossRef]
- Campbell, E.M.; Mcintosh, C.H.; Bowman, A.S. A Toolbox for Quantitative Gene Expression in Varroa destructor: RNA Degradation in Field Samples and Systematic Analysis of Reference Gene Stability. PLoS ONE 2016, 11, e0155640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, T.D.; Schmittgen, K.J. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Soroker, V.; Singh, N.K.; Eliash, N.; Plettner, E. Olfaction as a Target for Control of Honeybee Parasite Mite Varroa destructor. In Olfactory Concepts of Insect Control-Alternative to Insecticides; Springer: Cham, Switzerland, 2019; pp. 117–134. [Google Scholar]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nganso, B.T.; Mani, K.; Altman, Y.; Rafaeli, A.; Soroker, V. How Crucial is the Functional Pit Organ for the Varroa Mite? Insects 2020, 11, 395. https://doi.org/10.3390/insects11060395
Nganso BT, Mani K, Altman Y, Rafaeli A, Soroker V. How Crucial is the Functional Pit Organ for the Varroa Mite? Insects. 2020; 11(6):395. https://doi.org/10.3390/insects11060395
Chicago/Turabian StyleNganso, Beatrice T., Kannan Mani, Yam Altman, Ada Rafaeli, and Victoria Soroker. 2020. "How Crucial is the Functional Pit Organ for the Varroa Mite?" Insects 11, no. 6: 395. https://doi.org/10.3390/insects11060395
APA StyleNganso, B. T., Mani, K., Altman, Y., Rafaeli, A., & Soroker, V. (2020). How Crucial is the Functional Pit Organ for the Varroa Mite? Insects, 11(6), 395. https://doi.org/10.3390/insects11060395