Tracking Red Palm Mite Damage in the Western Hemisphere Invasion with Landsat Remote Sensing Data

 ,
,  ,
,  ,
, 

Abstract
:Simple Summary
Abstract
1. Introduction
2. Background
2.1. Taxonomy and Geography
2.2. Remote Sensing with Landsat Satellites
3. Methods
3.1. PROSAIL Simulations
3.2. Change Detection Analysis
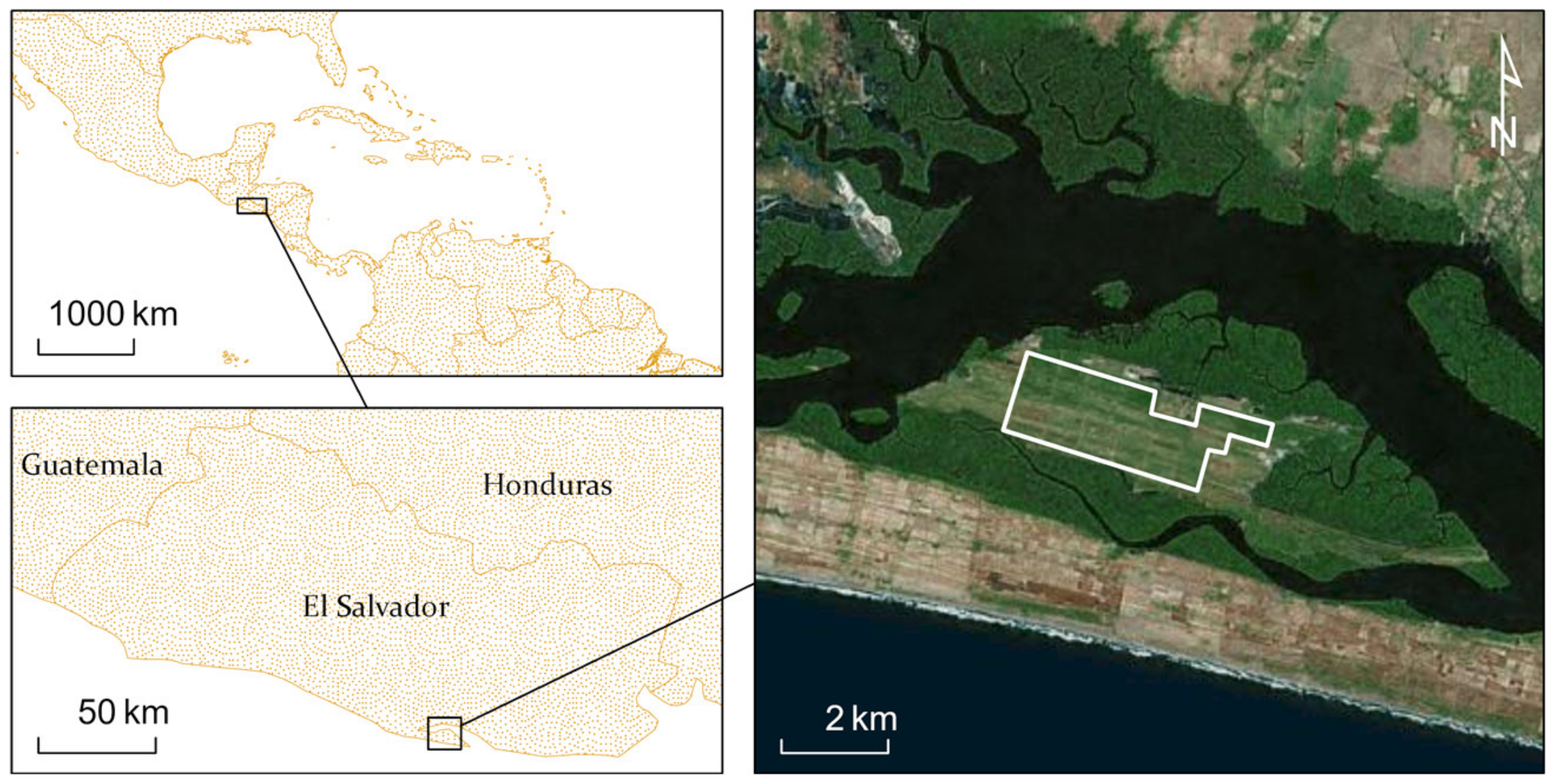
3.3. Study Sites
4. Results
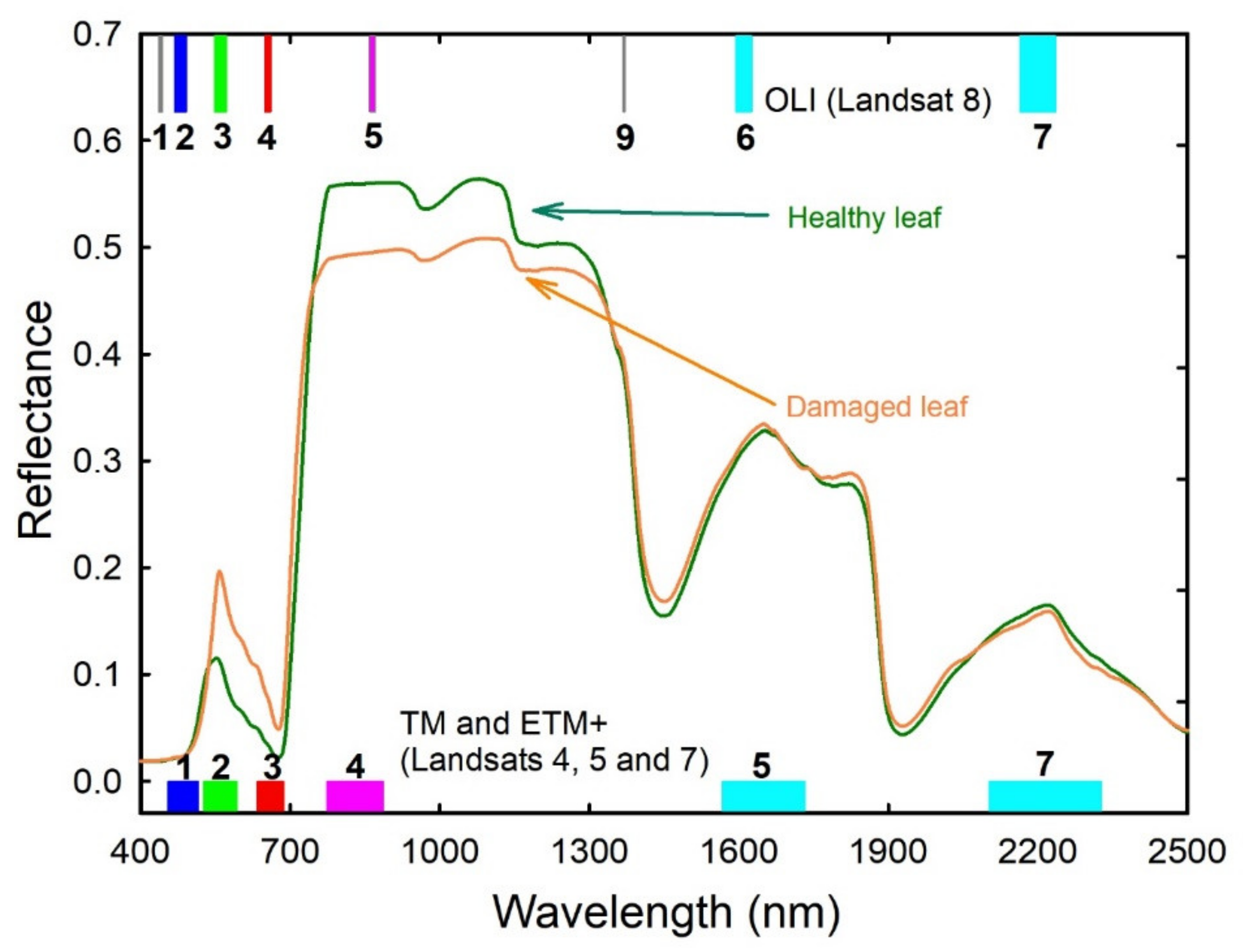
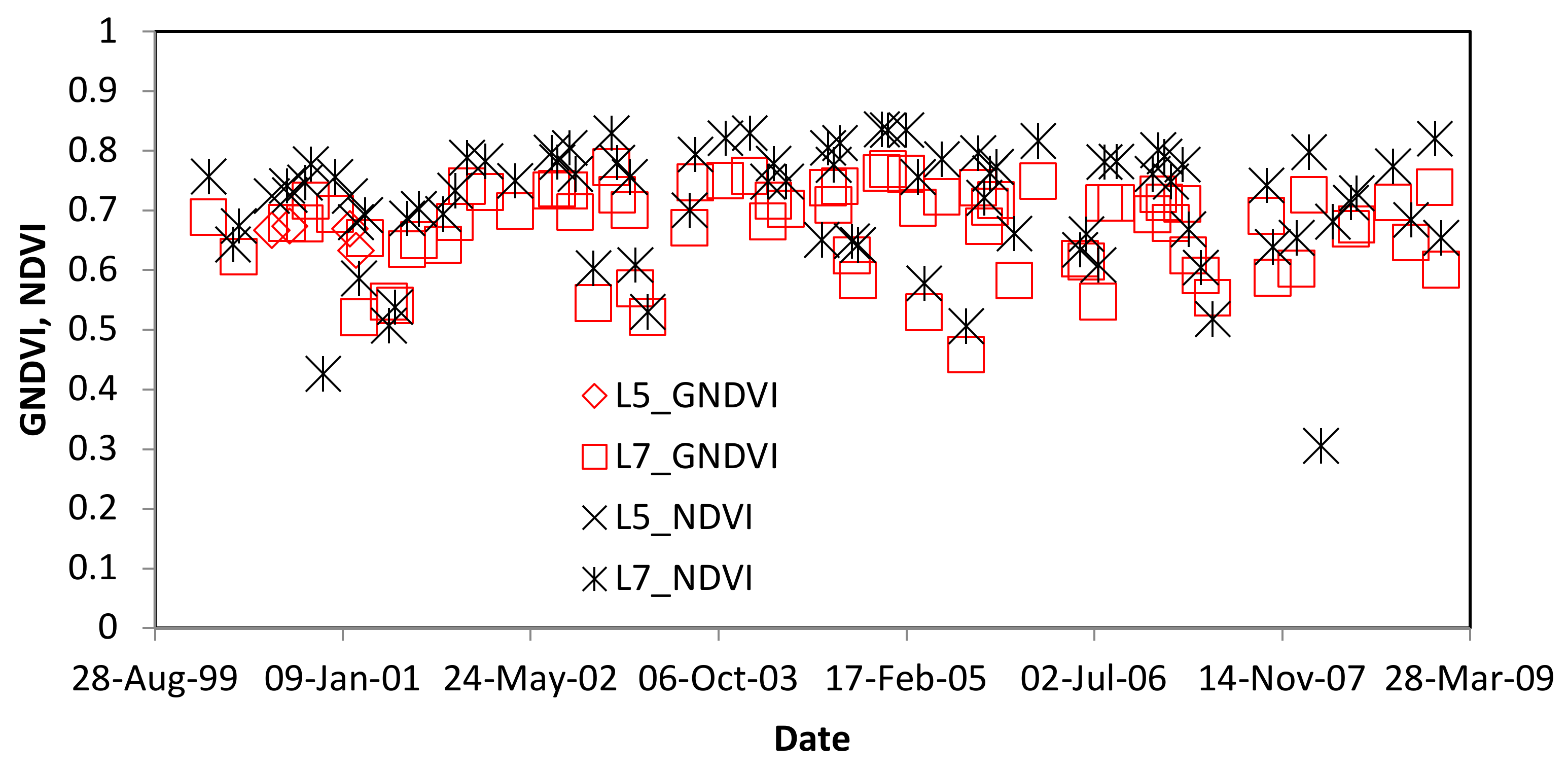
4.1. Predicted Remote Sensing Signal for RPM damage
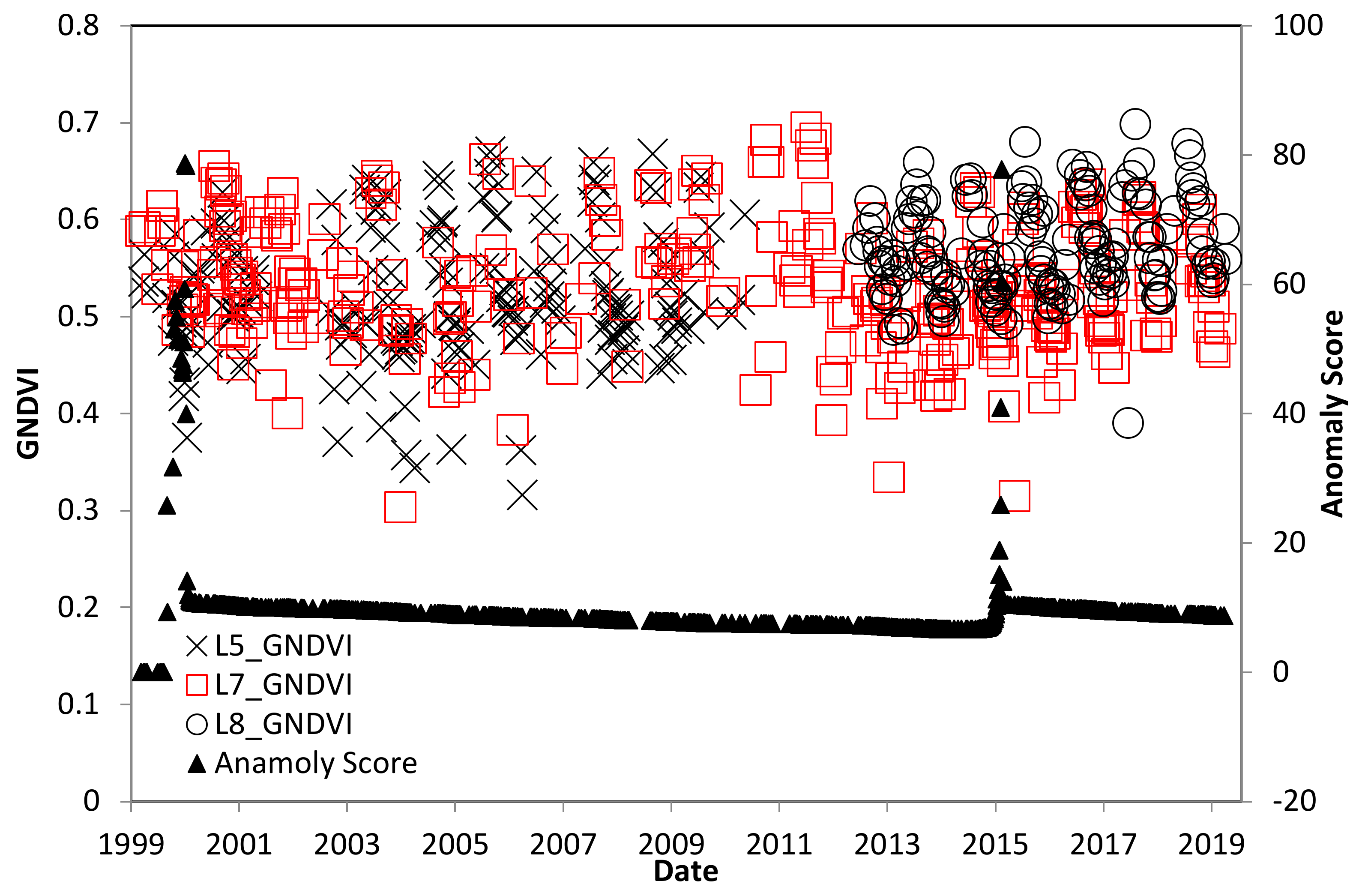
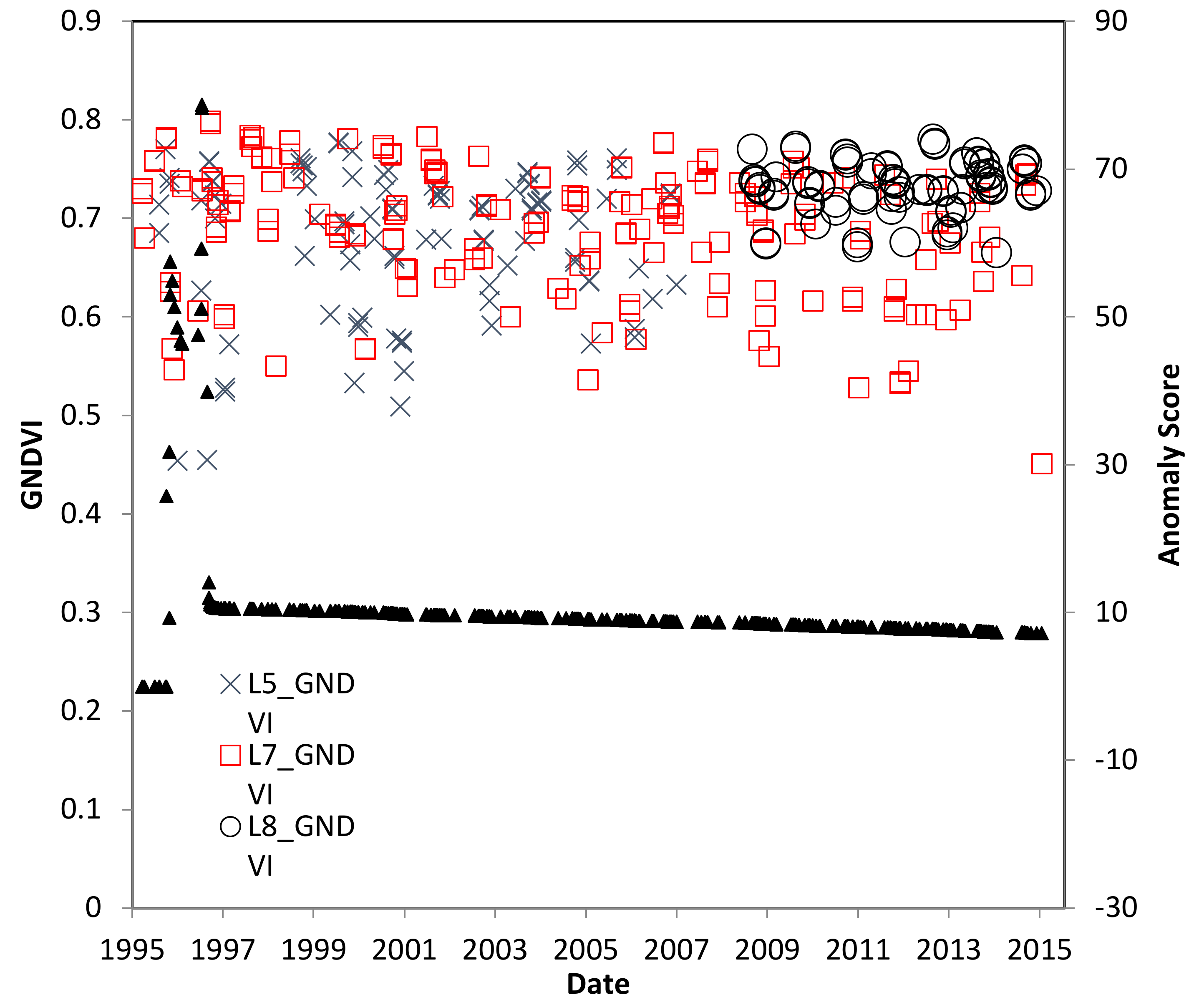
4.2. Trinidad Study Site
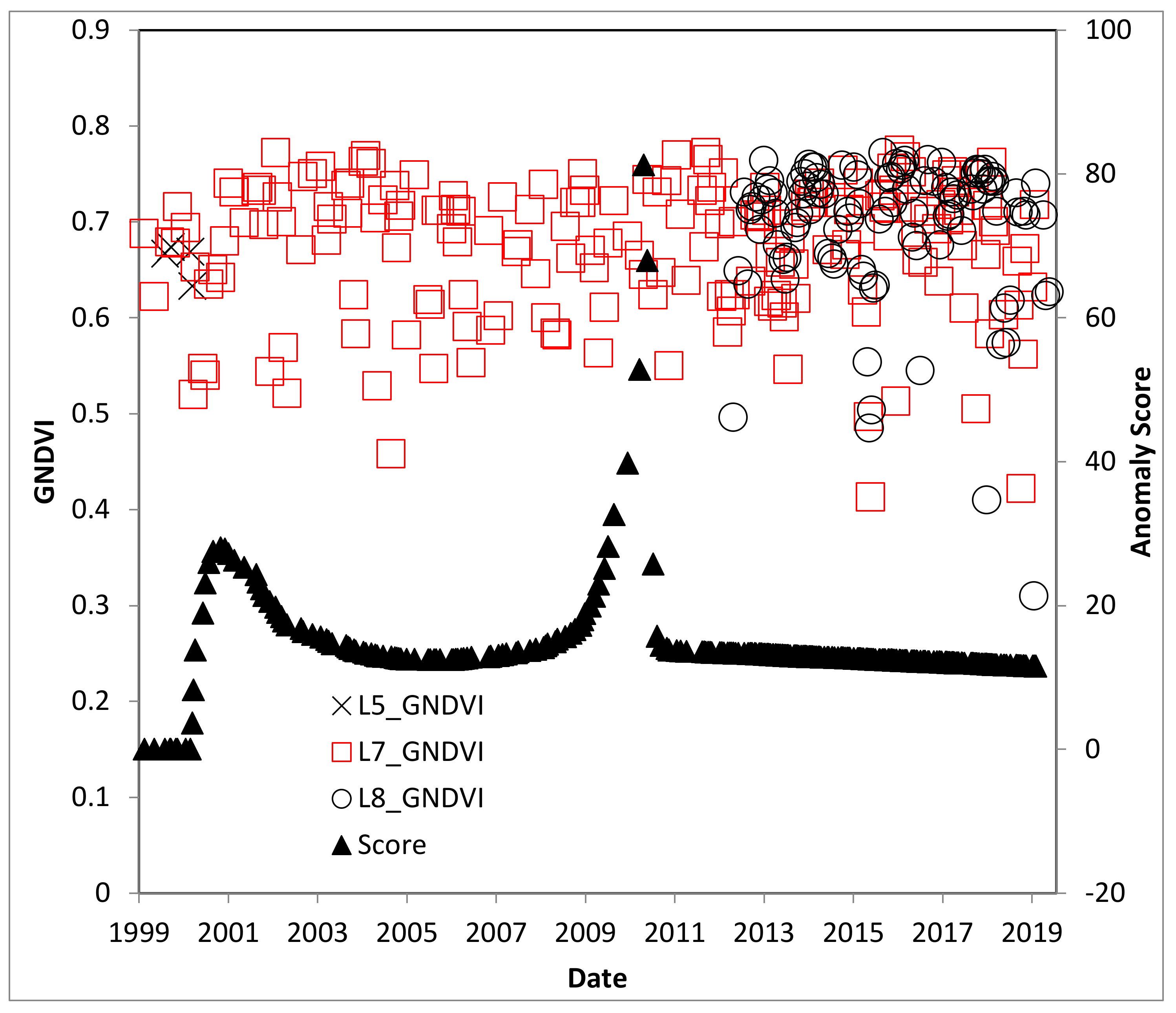
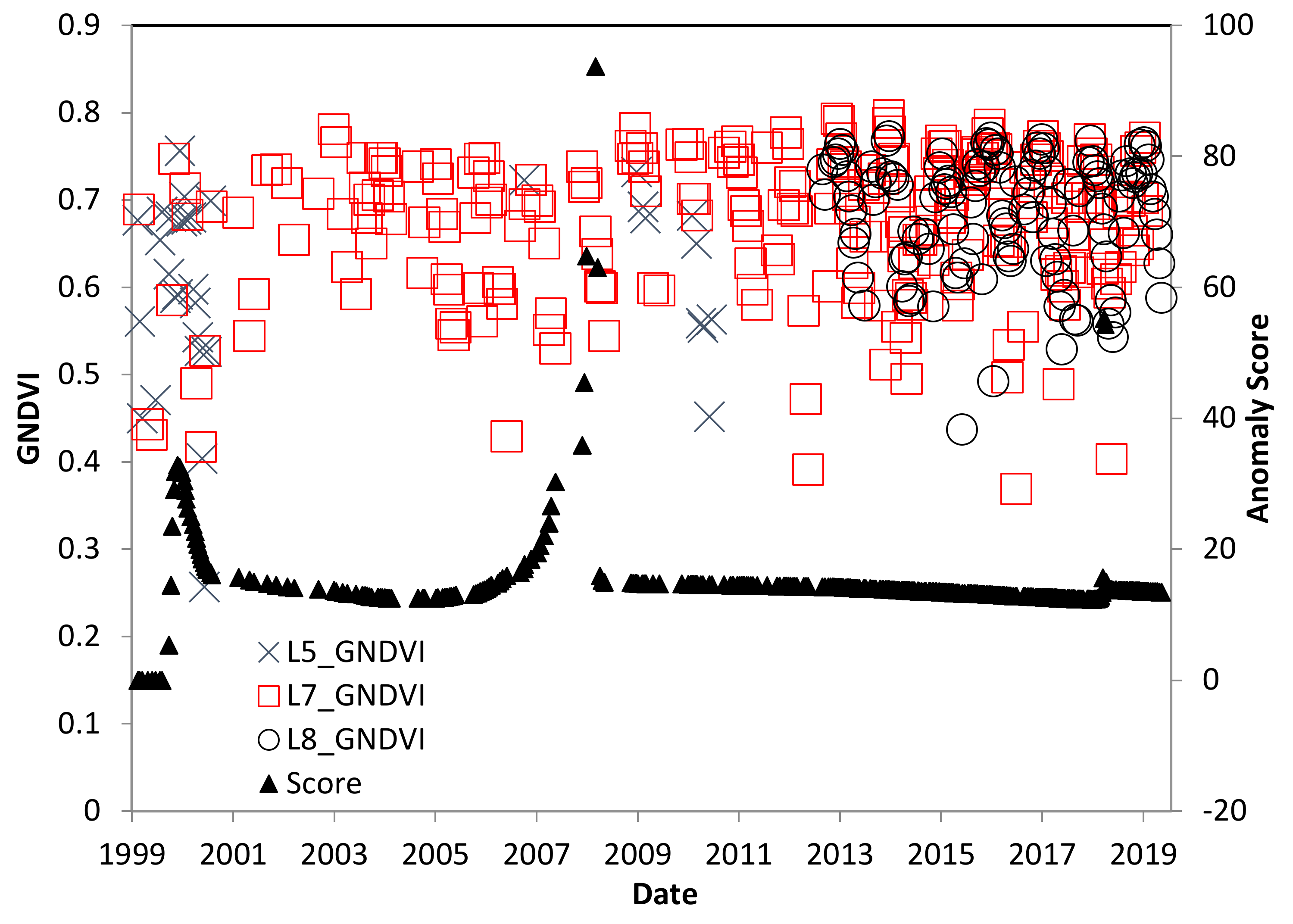
4.3. El Salvador Study Site
4.4. Ceara, Brazil Study Site
4.5. Pará, Brazil Study Site
5. Discussion
5.1. Spectral Information for Detecting Leaf Damage by RPM
5.2. Spatial and Temporal Resolution
5.3. Other Sensor Systems
5.3.1. Off-Nadir Sensors
5.3.2. Synthetic Aperture Radar (SAR)
5.3.3. Light Detection and Ranging (LIDAR)
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Island/Country | Year | Reference |
|---|---|---|
| Martinique | 2004 | Flechtmann and Etienne [69] |
| Saint Lucia | 2004 | Kane et al. [70,71] |
| Dominica | 2005 | Kane et al. [70,71] |
| Dominican Republic | 2006 | Rodriguez et al. [29] |
| Guadeloupe | 2006 | Etienne and Flechtmann [72] |
| Saint Martin | 2006 | Etienne and Flechtmann [72] |
| Trinidad | 2006 | Kane and Ochoa [2] |
| Puerto Rico | 2006 | Rodrigues et al. [29] |
| Florida, United States | 2007 | Welbourn [19]; Feiber and Lemon [73] |
| Haiti | 2007 | Dowling et al. [17] |
| Venezuela | 2007 | Vásquez et al. [26] |
| Cuba | 2007 | Dowling et al. [17] |
| Turks & Caicos | 2008 | Navia et al. [74] |
| Mexico | 2007 | Beard et al. [15] |
| El Salvador | 2015 | Guzman et al. [22] |
| Roraima, Brazil | 2009 | Navia et al. [74] |
| Colombia | 2010 | Carrillo et al. [20] |
| Amazonas, Brazil | 2010 | Rodrigues and Antony [24] |
| Nicaragua | 2016 | Beard et al. [15] |
| Honduras | 2016 | Beard et al. [15] |
| Guatemala | 2017 | Garcia-Ochaeta [75] |
| Pará, Brazil | 2018 | Noronha et al. [25] |
| Ecuador | 2020 | Alcivar et al. [76] |
| Paraguay | 2020 | Ramirez et al. [77] |
Appendix B
| Waveband | L5-TM 1 | L7-ETM+ 2 | L8-OLI 3 | |||
|---|---|---|---|---|---|---|
| Number | Wavelength (μm) | Number | Wavelength (μm) | Number | Wavelength (μm) | |
| Coastal | 1 | 0.43–0.45 | ||||
| Blue | 1 | 0.45–0.52 | 1 | 0.45–0.52 | 2 | 0.45–0.51 |
| Green | 2 | 0.52–0.60 | 2 | 0.52–0.60 | 3 | 0.53–0.59 |
| Red | 3 | 0.63–0.69 | 3 | 0.63–0.69 | 4 | 0.64–0.67 |
| Near Infrared | 4 | 0.76–0.90 | 4 | 0.77–0.90 | 5 | 0.85–0.88 |
| Shortwave Infrared 1 | 5 | 1.55–1.75 | 5 | 1.55–1.75 | 6 | 1.57–1.65 |
| Shortwave Infrared 2 | 7 4 | 2.08–2.35 | 7 4 | 2.08–2.35 | 7 | 2.11–2.29 |
| Panchromatic 5 | 8 | 0.52–0.90 | 8 | 0.50–0.68 | ||
| Cirrus | 9 | 1.36–1.38 | ||||
References
- Hirst, S. On some new species of red spider. Ann. Mag. Nat. Hist. 1924, 14, 522–527. [Google Scholar] [CrossRef]
- Kane, E.C.; Ochoa, R. Detection and Identification of the Red Palm Mite Raoiella indica Hirst (Acari: Tenuipalpidae); USDA: Beltsville, MD, USA, 2006. [Google Scholar]
- Cuenca, M.A.G.; Resende, J.M.; Saggin, O.J., Jr.; Reis, C.S. Mercado Brasileiro do Coco: Situação Atual e Perspectivas. In Coco Pós-Colheita; Aragão, W.M., Ed.; Embrapa Tabuleiros Costeiros: Aracaju, Brazil, 2002; pp. 11–18. [Google Scholar]
- Roda, A.; Dowling, A.; Welbourn, C.; Peña, J.; Rodrigues, J.C.V.; Hoy, M.A.; Ochoa, R.; Duncan, R.A.; de Chi, W. Red palm mite situation in the Caribbean and Florida. Proc. Carib. Food Crop. Soc. 2008, 44, 80–87. [Google Scholar]
- Roda, A.; Nachman, G.; Hosein, F.; Rodrigues, J.C.V.; Peña, J. Spatial distributions of the red palm mite, Raoiella indica (Acari: Tenuipalpidae) on coconut and their implications for development of efficient sampling plans. Exp. Appl. Acarol. 2012, 57, 291–308. [Google Scholar] [CrossRef] [PubMed]
- Nainanayake, A.D.; Gunathilake, J.; Kumarathunga, M.D.P.; Gunawardena, N.P.M.; Wijesekara, H.T.R. Limitation in the use of spectral analysis to detect Weligama Coconut Leaf Wilt Disease affected palms in Southern Sri Lanka. COCOS 2016, 22, 13–24. [Google Scholar] [CrossRef]
- Chong, K.L.; Kanniah, K.D.; Pohl, C.; Tan, K.P. A review of remote sensing applications for oil palm studies. Geo Spat. Inf. Sci. 2017, 20, 184–200. [Google Scholar] [CrossRef] [Green Version]
- Vogelmann, J.E.; Khoa, P.V.; Lan, D.X.; Shermeyer, J.; Shi, H.; Wimberly, M.C.; Duong, H.T.; Huong, L.V. Assessment of forest degradation in Vietnam using Landsat time series data. Forests 2017, 8, 238. [Google Scholar] [CrossRef]
- Lubkin, S.; Marrs, J.; McCartney, S.; Thieme, A. Mites from Space: Tracking a Microscopic Pest in Puerto Rico. DEVELOP, NASA-Goddard Space Flight Center. Available online: https://earthzine.org/mites-from-space-tracking-a-microscopic-pest-in-puerto-rico (accessed on 16 April 2020).
- Meyer, M.K.P. The Tenuipalpidae (Acari) of Africa with Keys to the World Fauna. In Entomology Memoir, Pretoria; Department of Agriculture Republic South Africa: Preporia, South Africa, 1979; Volume 50, pp. 1–133. [Google Scholar]
- Chaudhri, W.M.; Akbar, S. Studies on the Biosystematics and Control of Mites of Field Crops, Vegetables and Fruit Plants in Pakistan; University of Agriculture: Faisalabad, Pakistan, 1985; p. 341. [Google Scholar]
- Akbar, C.; Akbar, S.; Chaudhri, W.M. A new species of genus Raoiella (Acarina: Tenuipalpidae) from date palm. Pak. Entomol. 1987, 9, 41–44. [Google Scholar]
- Akbar, S. New species of genus Raoiella (Acarina: Tenuipalpidae) from Pakistan and their phenetic affinities. Pak. Entomol. 1990, 12, 75–81. [Google Scholar]
- Hasan, M.; Akbar, S. Genus Raoiella (Tenuipalpidae) from date palm in Punjab-Pakistan. Pak. Entomol. 2000, 22, 11–13. [Google Scholar]
- Beard, J.J.; Ochoa, R.; Bauchan, G.R.; Pooley, C.; Dowling, A.P.G. Raoiella of the world (Trombidiformes: Tetranychoisea: Tenuipalpidae). Zootaxa 2018, 4501, 1–301. [Google Scholar] [CrossRef]
- Dowling, A.P.; Ochoa, R.; Beard, J.J.; Welbourn, W.C.; Ueckermann, E.A. Phylogenetic investigation of the genus Raoiella (Prostigmata: Tenuipalpidae): Diversity, distribution, and world invasions. Exp. Appl. Acarol. 2012, 57, 257–269. [Google Scholar] [CrossRef]
- Dowling, A.P.; Ochoa, R.; Beard, J.J. Preliminary Results on Phylogeographic Patterns of the Invasive red Palm Mite, Raoiella indica (Prostigmata: Tenuipalpidae). In Proceedings of the 6th European Congress 2008, Montpellier, France, 21–25 July 2008; Bertrand, M., Kreiter, S., McCoy, K.D., Migeon, A., Navajas, M., Tixier, M.S., Vial, L., Eds.; European Association of Acarologists: Kent, UK, 2008; pp. 147–154. [Google Scholar]
- Jeppson, L.R.; Keifer, H.H.; Baker, E.W. Mites Injurious to Economic Plants; University of California Press: Berkeley, CA, USA, 1975. [Google Scholar]
- Welbourn, C. Pest Alert: Red Palm Mite Raoiella Indica Hirst (Acari: Tenuipalpidae); Florida Department of Agriculture and Consumer Services: Tallahassee, FL, USA, 2007. [Google Scholar]
- Carrillo, D.; Frank, J.H.; Rodrigues, J.C.V.; Peña, J.E. A review on the natural enemies of the red palm mite, Raoiella indica (Acari: Tenuipalpidae). Exp. Appl. Acarol. 2012, 57, 347–360. [Google Scholar] [CrossRef]
- Albinet, C.; Whitehurst, A.S.; Jewell, L.A. A Joint ESA-NASA Multi-mission Algorithm and Analysis Platform (MAAP) for Biomass, NISAR, and GEDI. Surv. Geophys. 2019, 40, 1017–1027. [Google Scholar] [CrossRef] [Green Version]
- Guzmán de Serrano, R.F.; Menjivar-Silis, A.G.; Borja-Melara, C.A. Nueva Amenaza para el Cocotero (Cocos Nucifera) en El Salvador; Report; CENTA, Laboratorio de Parasitología Vegetal, Centro de Tecnologia Agropecuaria: Ciudad Arce La, República de El Salvador, 2020; pp. 1–5. [Google Scholar]
- Gondim, M.G., Jr.; Castro, T.M.; Marsaro, A.L., Jr.; Navia, D.; Melo, J.W.; Demite, P.R.; de Moraes, G.J. Can the red palm mite threaten the Amazon vegetation? Syst. Biodivers. 2012, 10, 527–535. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, J.C.V.; Antony, L.K. First Report of Raoiella Indica Hirst (Acari: Tenuipalpidae) in Amazonas State, Brazil. Florida Entomol. 2011, 94, 1073–1074. Available online: https://www.scielo.br/scielo.php?script=sci_arttext&pid=S1519-566X2011000300018 (accessed on 10 September 2020). [CrossRef]
- Noronha, A.C.S.; Corrêa, C.L.P.S.; Pereira, C.A.B. Distribuição de Raoiella indica (Acari:Tenuipalpidae) na região do Baixo Amazonas, Pará. In Proceedings of the Il Congresso Latinoamericano de Acarologica E VI Simpósio Brazileiro de Acarologia, Pirenópolis, Goiás, Brasil, 29 July–2 August 2018. [Google Scholar]
- Vásquez, C.; Quiros De, G.M.; Aponte, O.; Sandoval, D.M. Primer reporte de Raoiella indica Hirst (Acari: Tenuipalpidae) en Sur América. Neotrop. Entomol. 2008, 37, 739–740. [Google Scholar] [CrossRef] [Green Version]
- Barroso, G.; Rocha, C.M.; Moreira, G.F.; Hata, F.T.; Roggia, S.; Ventura, M.U.; Pasini, A.; da Silva, J.E.P.; Holtz, A.M.; de Moraes, G.J. What is the southern limit of the distribution of red palm mite, Raoiella indica (Acari: Tenuipalpidae), in agricultural lands in Brazil? Florida Entomol. 2009, 102, 581–585. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, J.C.; Peña, J.E. Chemical control of the red palm mite, Raoella indica (Acari: Tenuipalpidae) in banana and coconut. Exp. Appl. Acarol. 2012, 57, 317–329. [Google Scholar] [CrossRef]
- Rodrigues, J.C.V.; Ochoa, R.; Kane, E. First report of Raoiella indica Hirst (Acari: Tenuipalpidae) and its damage to coconut palms in Puerto Rico and Culebra Island. Internat. J. Acarol. 2007, 33, 3–5. [Google Scholar] [CrossRef]
- Loveland, T.R.; Dwyer, J.L. Landsat: Building a strong future. Remote Sens. Environ. 2012, 122, 22–29. [Google Scholar] [CrossRef]
- Dottavio, C.L.; Williams, D.L. Satellite technology: An improved means for monitoring forest insect defoliation. J. For. 1983, 81, 30–34. [Google Scholar]
- Nelson, R.F. Detecting forest canopy change due to insect activity using Landsat MSS. Photogramm. Eng. Remote Sens. 1983, 49, 1303–1314. [Google Scholar]
- Vogelmann, J.E.; Rock, B.N. Assessing forest damage in high-elevation coniferous forests in Vermont and New Hampshire using Thematic Mapper data. Remote Sens. Environ. 1988, 24, 227–246. [Google Scholar] [CrossRef]
- Vogelmann, J.E.; Rock, B.N. Use of Thematic Mapper data for the detection of forest damage caused by pear thrips. Remote Sens. Environ. 1989, 30, 217–225. [Google Scholar] [CrossRef]
- Williams, D.L.; Goward, S.; Arvidson, T. Landsat: Yesterday, today, and tomorrow. Photogramm. Eng. Remote Sens. 2006, 72, 1171–1178. [Google Scholar] [CrossRef]
- Roy, D.; Wulder, M.A.; Loveland, T.; Woodcock, C.E.; Allen, R.; Anderson, M.C.; Helder, D.; Irons, J.; Johnson, D.; Kennedy, R.; et al. Landsat-8: Science and product vision for terrestrial global change research. Remote Sens. Environ. 2014, 145, 154–172. [Google Scholar] [CrossRef] [Green Version]
- Loveland, T.R.; Irons, J.R. Landsat 8: The plans, reality and the legacy. Remote Sens. Environ. 2016, 185, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Goward, S.N.; Masek, J.G.; Gao, F.; Vermote, E.F.; Thomas, N.; Schleeweis, K.; Kennedy, R.E.; Zhu, Z.; Eidenshink, J.C.; et al. Development of time series stacks of Landsat images for reconstructing forest disturbance history. Int. J. Digit. Earth 2009, 2, 195–218. [Google Scholar] [CrossRef]
- Huang, C.; Goward, S.N.; Masek, J.G.; Thomas, N.; Zhu, Z.; Vogelmann, J.E. An automated approach for reconstructing recent forest disturbance history using dense Landsat time series stacks. Remote Sens. Environ. 2010, 114, 183–198. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Lee, J.S.H.; Wich, S.; Widayati, A.; Koh, L.P. Detecting industrial oil palm plantations on Landsat images with Google Earth Engine. Remote Sens. Appl. Soc. Environ. 2016, 4, 219–224. [Google Scholar] [CrossRef] [Green Version]
- Rouse, J.W., Jr.; Haas, R.H.; Schnell, J.A.; Deering, D.W. Monitoring Vegetation Systems in the Great Plains with ERTS. In Third Earth Resources Technology Satellite–1 Symposium; National Aeronautics and Space Administration: Washington, DC, USA, 1974; Volume 1, pp. 309–317. [Google Scholar]
- Tucker, C.J. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef] [Green Version]
- Gitelson, A.A.; Kaufman, Y.J.; Merzlyak, M.N. Use of a green channel in remote sensing of global vegetation from EOS-MODIS. Remote Sens. Environ. 1996, 58, 289–298. [Google Scholar] [CrossRef]
- Jacquemoud, S.; Verhoef, W.; Baret, F.; Bacour, C.; Zarco-Tejada, P.J.; Asner, G.P.; François, C.; Ustin, S. PROSPECT+SAIL models: A review of use for vegetation characterization. Remote. Sens. Environ. 2009, 113, S56–S66. [Google Scholar] [CrossRef]
- Féret, J.-B.; François, C.; Asner, G.P.; Gitelson, A.; Martin, R.E.; Bidel, L.P.; Ustin, S.; Le Maire, G.; Jacquemoud, S. PROSPECT-4 and 5: Advances in the leaf optical properties model separating photosynthetic pigments. Remote Sens. Environ. 2008, 112, 3030–3043. [Google Scholar] [CrossRef]
- Lelong, C.C.; Roger, J.-M.; Brégand, S.; Dubertret, F.; Lanore, M.; Sitorus, N.A.; Raharjo, D.A.; Caliman, J.-P. Evaluation of Oil-Palm Fungal Disease Infestation with Canopy Hyperspectral Reflectance Data. Sensors 2010, 10, 734–747. [Google Scholar] [CrossRef]
- Santoso, H.; Gunawan, T.; Jatmiko, R.H.; Darmosarkoro, W.; Minasny, B. Mapping and identifying basal stem rot disease in oil palms in North Sumatra with QuickBird imagery. Precis. Agric. 2011, 12, 233–248. [Google Scholar] [CrossRef]
- Shafri, H.Z.M.; Anuar, M.I.; Seman, I.A.; Noor, N.M. Spectral discrimination of healthy and Ganoderma-infected oil palms from hyperspectral data. Int. J. Remote Sens. 2011, 32, 7111–7129. [Google Scholar] [CrossRef]
- Liaghat, S.; Ehsani, R.; Mansor, S.; Shafri, H.Z.M.; Meon, S.; Sankaran, S.; Azam, S.H.M.N. Early detection of basal stem rot disease (Ganoderma) in oil palms based on hyperspectral reflectance data using pattern recognition algorithms. Int. J. Remote Sens. 2014, 35, 3427–3439. [Google Scholar] [CrossRef]
- Ahmadi, P.; Muharam, F.M.; Ahmad, K.; Mansor, S.; Seman, I.A. Early detection of Ganoderma basal stem rot of oil palms using artificial neural network spectral analysis. Plant Dis. 2017, 101, 1009–1016. [Google Scholar] [CrossRef] [Green Version]
- Gapor, A.; Kato, A.; Ong, A.S.H. α-Tocopherol content in oil palm leaflet. J. Amer. Oil Chem. Soc. 1986, 63, 330–331. [Google Scholar] [CrossRef]
- Awal, M.A.; Ishak, W.; Endan, J.; Haniff, M. Determination of specific leaf area and leaf area-mass relationship in oil palm plantation. Asian J. Plant. Sci. 2004, 3, 264–268. [Google Scholar]
- Tan, K.P.; Kanniah, K.D.; Cracknell, A.P. On the upstream inputs into the MODIS primary productivity products using biometric data from oil palm plantations. Int. J. Remote Sens. 2014, 35, 2215–2246. [Google Scholar] [CrossRef]
- Barsi, J.A.; Lee, K.; Kvaran, G.; Markham, B.L.; Pedelty, J.A. The spectral response of the Landsat-8 Operational Land Imager spectral response and spectral uniformity. Remote Sens. 2014, 6, 10232–10251. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, J.; Yamanishi, K. A unifying framework for detecting outliers and change points from time series. IEEE Trans. Knowl. Data Eng. 2006, 18, 482–492. [Google Scholar] [CrossRef]
- Urabe, Y.; Yamanishi, K.; Tomioka, R.; Iwai, H. Real-time change-point detection using sequentially discounting normalized maximum likelihood coding. Adv. Knowl. Discov. Data Min. 2011, 6635, 185–197. [Google Scholar] [CrossRef]
- Lawhern, V.; Scott, K.; Kay, A.R. Detecting Alpha Spindle Events in EEG Time Series Using Adaptive Autoregressive Models. BMC Neurosci. 2013, 14, 101. [Google Scholar] [CrossRef] [Green Version]
- Vogelmann, J.E. Comparison between two vegetation indices for measuring different types of forest damage in the north-eastern United States. Int. J. Remote Sens. 1990, 11, 2281–2297. [Google Scholar] [CrossRef]
- Daughtry, C.S.T.; Walthall, C.L.; Kim, M.S.; Brown de Colstoun, E.; McMurtrey, J.E., III. Estimating corn leaf chlorophyll concentration from leaf and canopy reflectance. Remote Sens. Environ. 2000, 74, 229–239. [Google Scholar] [CrossRef]
- Hunt, E.R., Jr.; Horneck, D.A.; Spinelli, C.B.; Turner, R.W.; Bruce, A.E.; Gadler, D.J.; Brungardt, J.J.; Hamm, P.B. Monitoring nitrogen status of potatoes using small unmanned aerial vehicles. Precis. Agric. 2018, 19, 314–333. [Google Scholar] [CrossRef]
- Immitzer, M.; Vuolo, F.; Atzberger, C. First experience with Sentinel-2 data for crop and tree species classifications in central Europe. Remote Sens. 2016, 8, 166. [Google Scholar] [CrossRef]
- Claverie, M.; Ju, J.; Masek, J.G.; Dungan, J.L.; Vermote, E.F.; Roger, J.C.; Skakun, S.V.; Justice, C. The harmonized Landsat and Sentinel-2 surface reflectance data set. Remote Sens. Environ. 2018, 219, 145–161. [Google Scholar] [CrossRef]
- Filho, F.H.I.; Heldens, W.B.; Kong, Z.; de Lange, E.S. Drones: Innovative technology for use in precision pest management. J. Econ. Entomol. 2020, 113, 1–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morel, A.C.; Saatchi, S.S.; Malhi, Y.; Berry, N.J.; Banin, L.; Burslem, D.; Nilus, R.; Ong, R.C. Estimating aboveground biomass in forest and oil palm plantation in Sabah, Malaysian Borneo using ALOS PALSAR data. For. Ecol. Manag. 2011, 262, 1786–1798. [Google Scholar] [CrossRef]
- Tan, K.P.; Kanniah, K.D.; Cracknell, A.P. Use of UK-DMC 2 and ALOS PALSAR for studying the age of oil palm trees in southern peninsular Malaysia. Int. J. Remote Sens. 2013, 34, 7424–7446. [Google Scholar] [CrossRef]
- Cheng, Y.; Yu, L.; Xu, Y.; Lu, H.; Cracknell, A.P.; Kanniah, K.; Gong, P. Mapping oil palm extent in Malaysia using ALOS-2 PALSAR-2 data. Int. J. Remote Sens. 2018, 39, 432–452. [Google Scholar] [CrossRef]
- Boudreau, J.; Nelson, R.F.; Margolis, H.A.; Beaudoin, A.; Guindon, L.; Kimes, D.S. Regional aboveground forest biomass using airborne and spaceborne LiDAR in Québec. Remote Sens. Environ. 2008, 112, 3876–3890. [Google Scholar] [CrossRef]
- Flechtmann, C.H.W.; Etienne, J. The red palm mite, Raoiella indica Hirst, a threat to palms in the Americas (Acari: Prostigmata: Tenuipalpidae). Syst. Appl. Acarol. 2004, 9, 109–110. [Google Scholar] [CrossRef]
- Kane, E.C.; Ochoa, R.; Mathurin, G.; Erbe, E.F. Raoiella Indica Hirst (Acari: Tenuipalpidae): An Island-Hopping Mite Pest in the Caribbean. Entomological Society of America Meeting. Available online: http://www.doc-developpement-durable.org/file/Arbres-Fruitiers/FICHES_ARBRES/Palmier-dattier/maladies/Raoiella%20indica%20Hirst_Trinidad.pdf (accessed on 10 September 2020).
- Kane, E.C.; Ochoa, R.; Mathurin, G.; Erbe, E.F.; Beard, J.J. Raoiella indica Hirst (Acari: Tenuipalpidae): An island-hopping mite pest in the Caribbean. Exp. Appl. Acarol. 2012, 57, 215–225. [Google Scholar] [CrossRef]
- Etienne, J.; Flechtmann, C.H.W. First record of Raoiella indica (Hirst, 1924) (Acari: Tenuipalpidae) in Guadeloupe and Saint Martin, West Indies. Internat. J. Acarol. 2006, 32, 331–332. [Google Scholar] [CrossRef]
- Feiber, D.; Lemon, N. Red Palm Mite Infestation Detected in Palm gardens. Florida Department of Agriculture, Department Press Release. 2007. Available online: http://www.doacs.state.fl.us/press/2007/12052007_2.html (accessed on 5 April 2010).
- Navia, D.; Marsaro, A.K., Jr.; Da Silva, F.R.; Gondim, M.G., Jr.; De Moraes, G.J. First report of the red palm mite, Raoiella indica Hirst (Acari: Tenuipalpidae) in Brazil. Neotrop. Entomol. 2011, 40, 409–411. [Google Scholar] [CrossRef] [Green Version]
- García Ochaeta, J.F. Primer Registro De Raoiella Indica Hirst, 1924 (Acari: Tenuipalpidae) En Guatemala. 2018. Available online: https://journals.flvc.org/mundi/article/view/0607/101213 (accessed on 10 September 2020).
- Alcivar, J.; Mesa, N.C.; Vasquez, C. First report of Raoiella indica Hisrt (Acari: Tenuipalpidae) in Province of Manabi, Ecuador. Int. J. Acarol. 2020, 46, 120–122. [Google Scholar] [CrossRef]
- Ramírez, M.B.; Sarubbi, H.J.; Arias, O.; De Azevedo, L.H.; Flechtmann, C.H.W. First report of Raoiella indica Hirst (Acari: Tenuipalpidae) in Paraguay. J. Plant Dis. Prot. 2020, 1–3. [Google Scholar] [CrossRef]













| Study Site | Latitude | Longitude | Area of Study Site (hectares) | Date First RPM Observation |
|---|---|---|---|---|
| Trinidad | 10.057° N | 61.900° W | ~700 (220) | October 2006 |
| El Salvador | 13.241° N | 88.606° W | ~1000 (500) | January 2015 |
| Ceara, Brazil | 2.974° S | 39.793° W | ~5000 (16) | May 2016 |
| Pará, Brazil | 2.141° S | 48.630° W | ~3800 (11) | None |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodrigues, J.C.V.; Cosh, M.H.; Hunt, E.R., Jr.; de Moraes, G.J.; Barroso, G.; White, W.A.; Ochoa, R. Tracking Red Palm Mite Damage in the Western Hemisphere Invasion with Landsat Remote Sensing Data. Insects 2020, 11, 627. https://doi.org/10.3390/insects11090627
Rodrigues JCV, Cosh MH, Hunt ER Jr., de Moraes GJ, Barroso G, White WA, Ochoa R. Tracking Red Palm Mite Damage in the Western Hemisphere Invasion with Landsat Remote Sensing Data. Insects. 2020; 11(9):627. https://doi.org/10.3390/insects11090627
Chicago/Turabian StyleRodrigues, Jose Carlos Verle, Michael H. Cosh, E. Raymond Hunt, Jr., Gilberto J. de Moraes, Geovanny Barroso, William A. White, and Ronald Ochoa. 2020. "Tracking Red Palm Mite Damage in the Western Hemisphere Invasion with Landsat Remote Sensing Data" Insects 11, no. 9: 627. https://doi.org/10.3390/insects11090627
APA StyleRodrigues, J. C. V., Cosh, M. H., Hunt, E. R., Jr., de Moraes, G. J., Barroso, G., White, W. A., & Ochoa, R. (2020). Tracking Red Palm Mite Damage in the Western Hemisphere Invasion with Landsat Remote Sensing Data. Insects, 11(9), 627. https://doi.org/10.3390/insects11090627