Potato Tuberworm Phthorimaea operculella (Zeller) (Lepidoptera: Gelechioidea) Leaf Infestation Affects Performance of Conspecific Larvae on Harvested Tubers by Inducing Chemical Defenses


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plants
2.2. Insects
2.3. Leaf Herbivory by P. operculella Larvae and Plant Responses
2.4. Effects of Aboveground Herbivory on the Performance of Larvae Feeding on Tubers
2.5. Analysis of Plant Chemistry
2.6. Data Analysis
3. Results
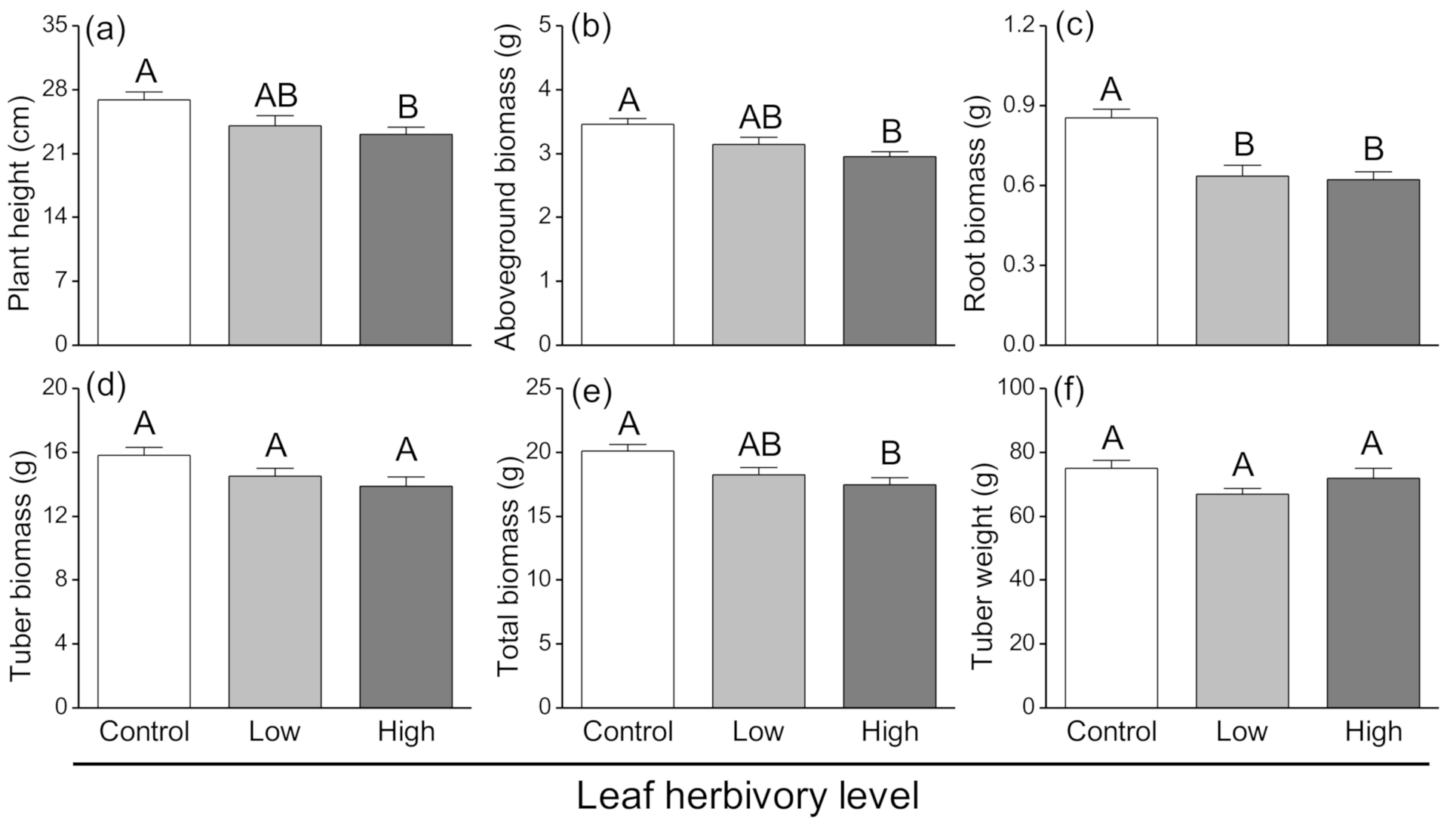
3.1. Plant Growth Response to Aboveground Herbivory
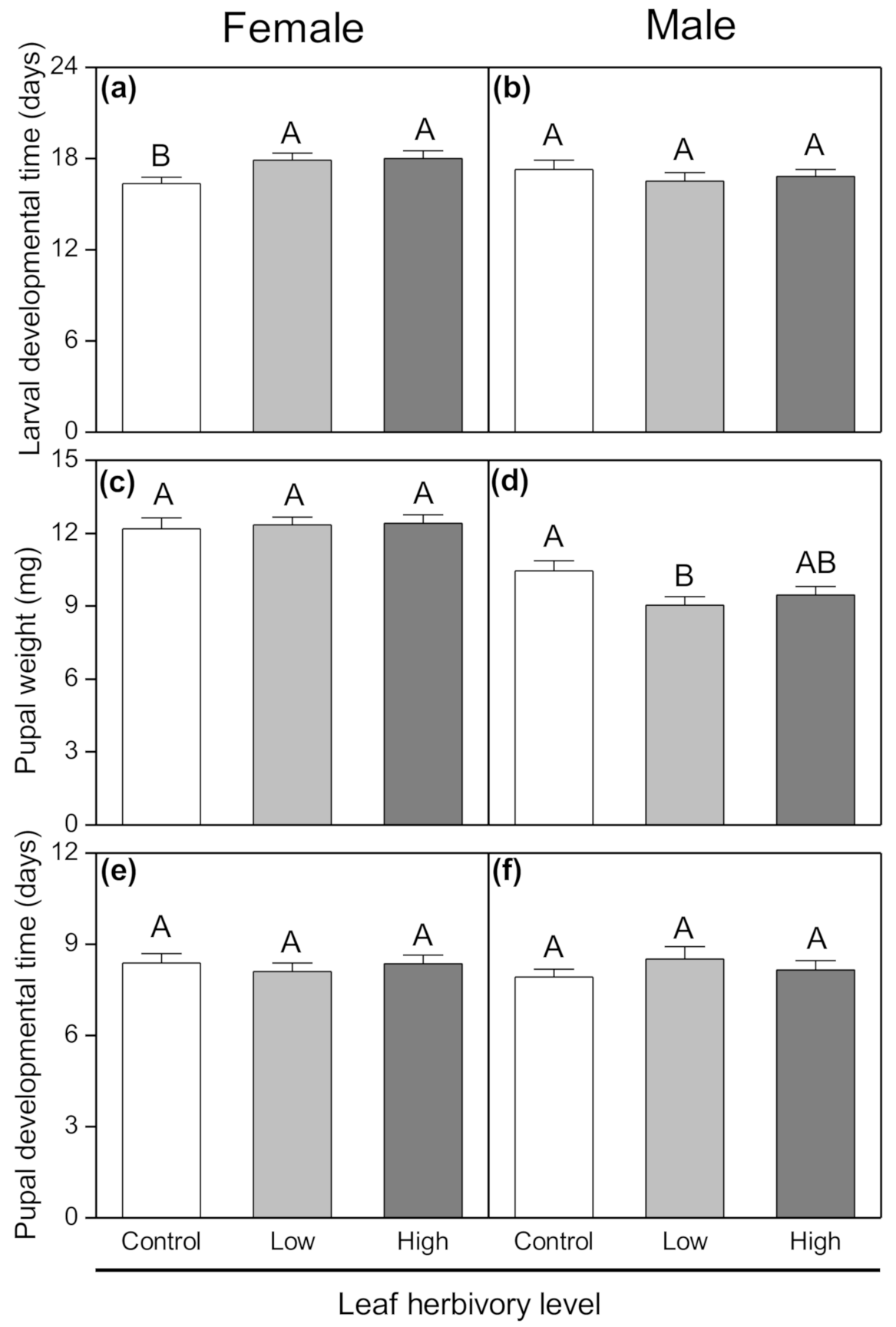
3.2. Development Performance of Tuber-Feeding Larvae
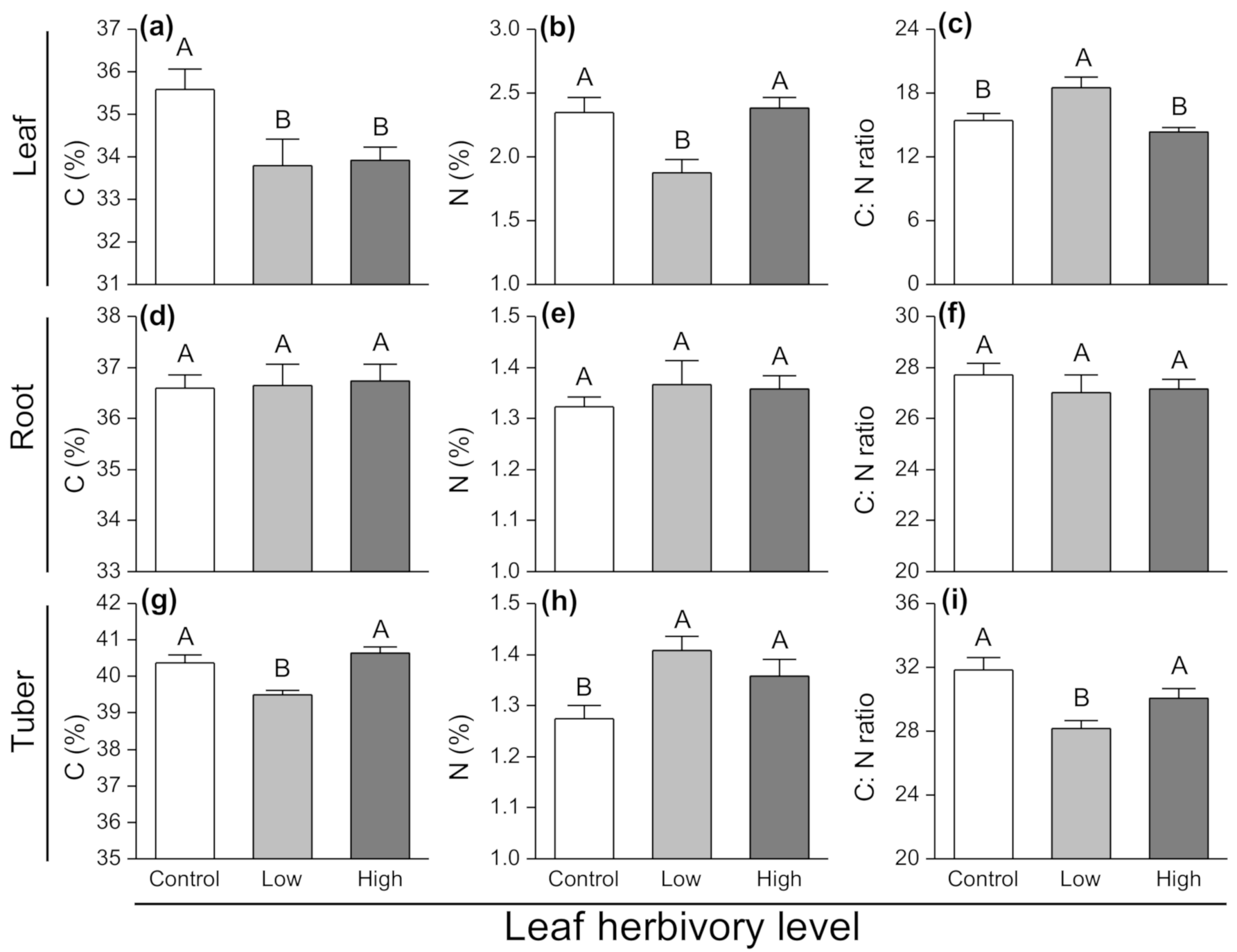
3.3. Effects of Leaf Herbivory on Plant Nutrients
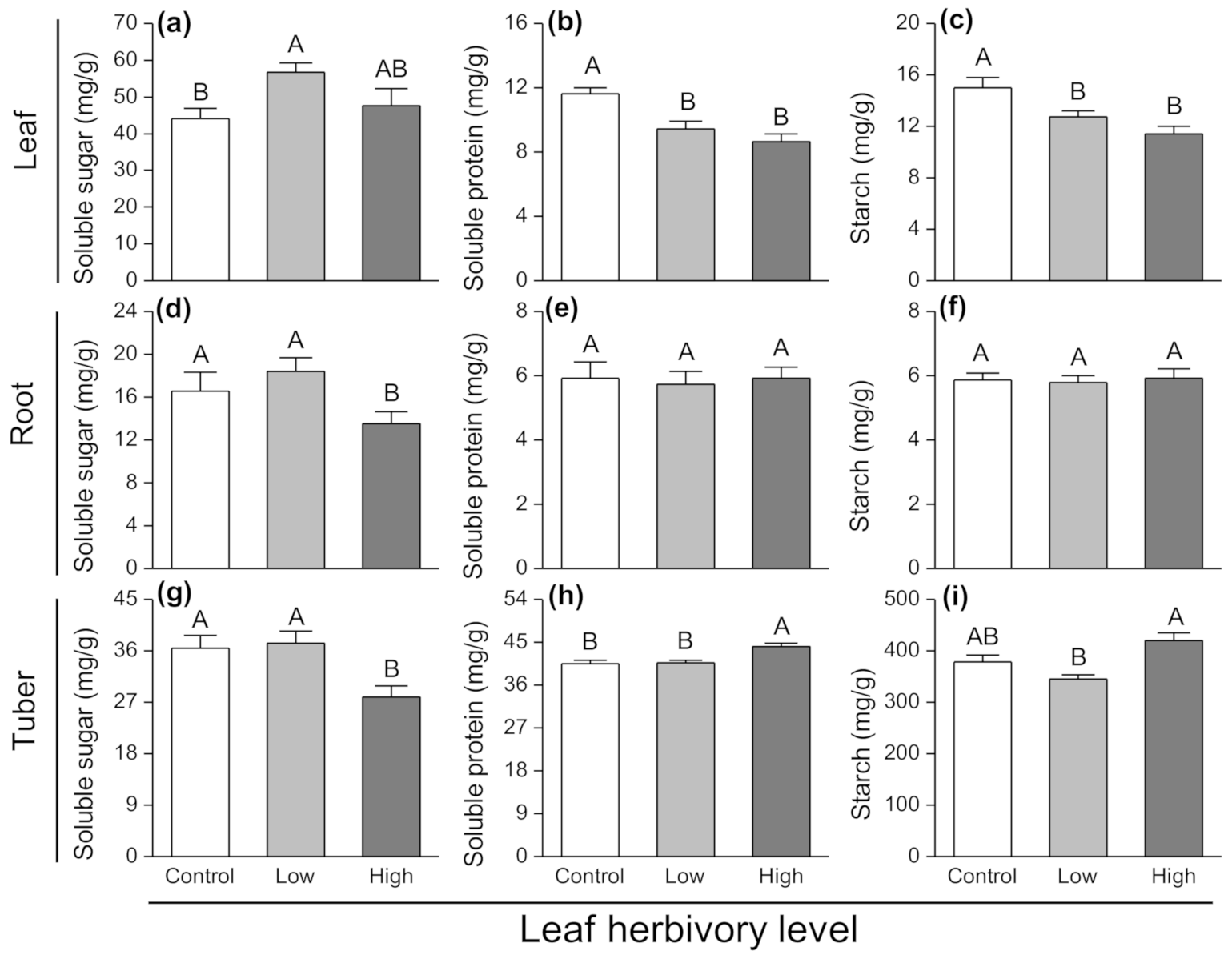
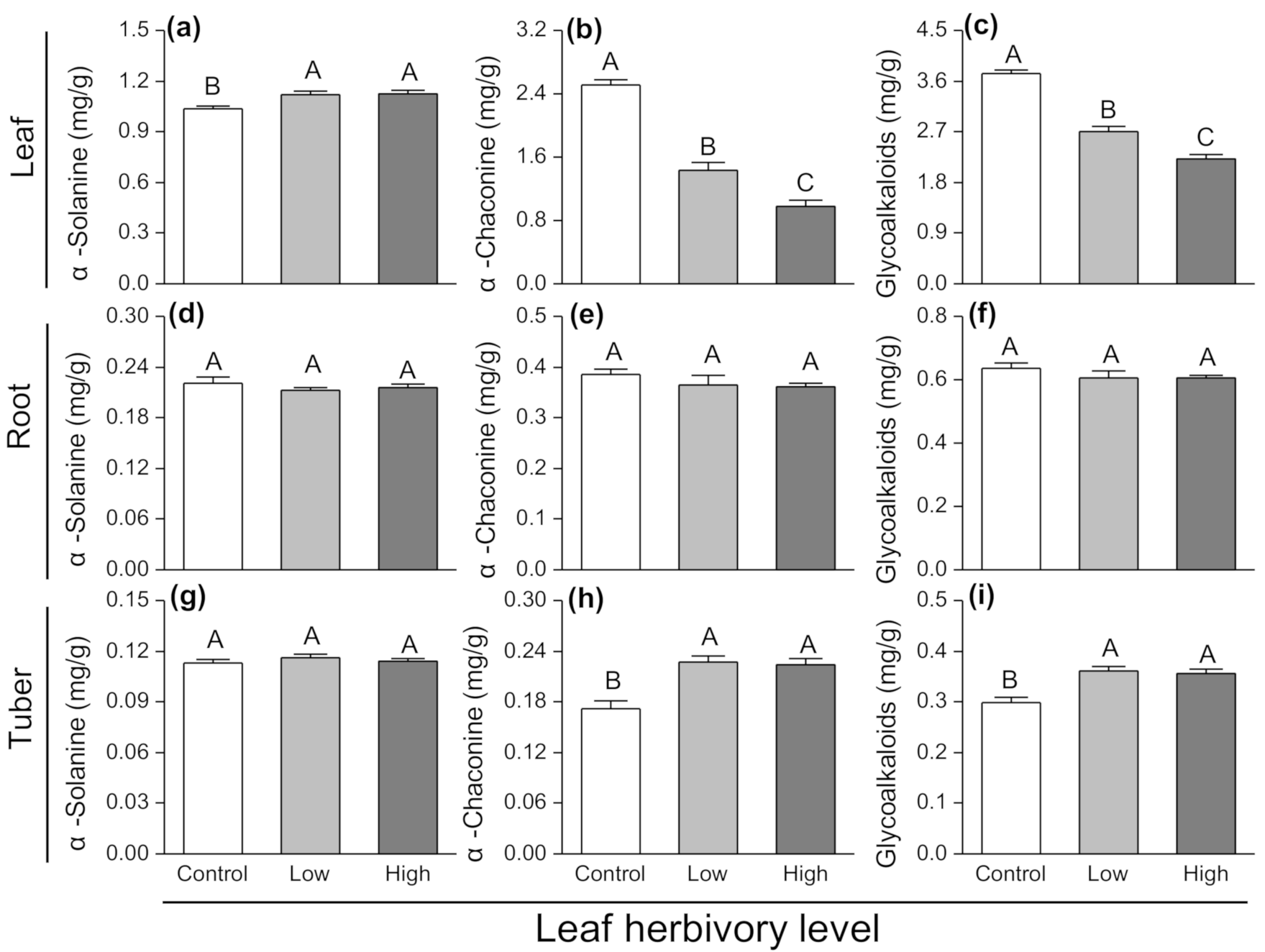
3.4. Effects of Leaf Herbivory on Plant Secondary Chemicals
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van der Putten, W.H.; Vet, L.E.M.; Harvey, J.A.; Wackers, F.L. Linking above- and belowground multitrophic interactions of plants, herbivores, pathogens, and their antagonists. Trends Ecol. Evol. 2001, 16, 547–554. [Google Scholar] [CrossRef]
- Huang, W.; Siemann, E.; Xiao, L.; Yang, X.; Ding, J. Species-specific defence responses facilitate conspecifics and inhibit heterospecifics in above-belowground herbivore interactions. Nat. Commun. 2014, 5, 4851. [Google Scholar] [CrossRef] [PubMed]
- Arce, C.C.M.; Machado, R.A.R.; Ribas, N.S.; Cristaldo, P.F.; Ataíde, L.M.S.; Pallini, Â.; Carmo, F.M.; Freitas, L.G.; Lima, E. Nematode root herbivory in tomato increases leaf defenses and reduces leaf miner oviposition and performance. J. Chem. Ecol. 2017, 43, 120–128. [Google Scholar] [CrossRef]
- Machado, R.A.R.; Arce, C.C.M.; McClure, M.A.; Baldwin, I.T.; Erb, M. Aboveground herbivory induced jasmonates disproportionately reduce plant reproductive potential by facilitating root nematode infestation. Plant Cell Environ. 2018, 41, 797–808. [Google Scholar] [CrossRef] [PubMed]
- Abdala-Roberts, L.; Pérez Niño, B.; Moreira, X.; Parra-Tabla, V.; Grandi, L.; Glauser, G.; Benrey, B.; Turlings, T.C.J. Effects of early-season insect herbivory on subsequent pathogen infection and ant abundance on wild cotton (Gossypium hirsutum). J. Ecol. 2019, 107, 1518–1529. [Google Scholar] [CrossRef]
- Masters, G.J.; Brown, V.K.; Gange, A.C. Plant mediated interactions between aboveground and below-ground insect herbivores. Oikos 1993, 66, 148. [Google Scholar] [CrossRef]
- Bezemer, T.M.; Wagenaar, R.; van Dam, N.M.; Wackers, F.L. Interactions between above- and belowground insect herbivores as mediated by the plant defense system. Oikos 2003, 101, 555–562. [Google Scholar] [CrossRef]
- Huang, W.; Siemann, E.; Yang, X.; Wheeler, G.S.; Ding, J. Facilitation and inhibition: Changes in plant nitrogen and secondary metabolites mediate interactions between above-ground and below-ground herbivores. Proc. R. Soc. B-Biol. Sci. 2013, 280, 20131318. [Google Scholar] [CrossRef] [PubMed]
- Van Geem, M.; Gols, R.; Raaijmakers, C.E.; Harvey, J.A. Effects of population-related variation in plant primary and secondary metabolites on aboveground and belowground multitrophic interactions. Chemoecology 2016, 26, 219–233. [Google Scholar] [CrossRef]
- Bezemer, T.M.; van Dam, N.M. Linking aboveground and belowground interactions via induced plant defenses. Trends Ecol. Evol. 2005, 20, 617–624. [Google Scholar] [CrossRef] [Green Version]
- Johnson, S.N.; Clark, K.E.; Hartley, S.E.; Jones, T.H.; McKenzie, S.W.; Koricheva, J. Aboveground-belowground herbivore interactions: A meta-analysis. Ecology 2012, 93, 2208–2215. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Ortiz, E.V.; Garrido, E.; Poveda, K.; Jander, G. Potato tuber herbivory increases resistance to aboveground lepidopteran herbivores. Oecologia 2016, 182, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, I.; Halitschke, R.; Kessler, A.; Rehill, B.J.; Sardanelli, S.; Denno, R.F. Physiological integration of roots and shoots in plant defense strategies links above- and belowground herbivory. Ecol. Lett. 2008, 11, 841–851. [Google Scholar] [CrossRef]
- Soler, R.; Bezemer, T.M.; Cortesero, A.M.; Van der Putten, W.H.; Vet, L.E.; Harvey, J.A. Impact of foliar herbivory on the development of a root-feeding insect and its parasitoid. Oecologia 2007, 152, 257–264. [Google Scholar] [CrossRef] [Green Version]
- Robert, C.A.; Erb, M.; Duployer, M.; Zwahlen, C.; Doyen, G.R.; Turlings, T.C. Herbivore-induced plant volatiles mediate host selection by a root herbivore. New Phytol. 2012, 194, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Mundim, F.M.; Alborn, H.T.; Vieira-Neto, E.H.M.; Bruna, E.M. A whole-plant perspective reveals unexpected impacts of above-and belowground herbivores on plant growth and defense. Ecology 2018, 98, 70–78. [Google Scholar] [CrossRef]
- Coley, P.D.; Bryant, J.P.; Chapin, F.S., 3rd. Resource availability and plant antiherbivore defense. Science 1985, 230, 895–899. [Google Scholar] [CrossRef] [Green Version]
- Bezemer, T.M.; Wagenaar, R.; Van Dam, N.M.; Van Der Putten, W.H.; Wackers, F.L. Above- and below-ground terpenoid aldehyde induction in cotton, Gossypium herbaceum, following root and leaf injury. J. Chem. Ecol. 2004, 30, 53–67. [Google Scholar] [CrossRef] [Green Version]
- Van Dam, N.M.; Witjes, L.; Svatos, A. Interactions between aboveground and belowground induction of glucosinolates in two wild Brassica species. New Phytol. 2004, 161, 801–810. [Google Scholar] [CrossRef]
- Mithoefer, A.; Boland, W. Plant defense against herbivores: Chemical aspects. Annu. Rev. Plant Biol. 2012, 63, 431–450. [Google Scholar] [CrossRef] [Green Version]
- Jackson, R.B.; Mooney, H.A.; Schulze, E.D. A global budget for fine root biomass, surface area, and nutrient contents. Proc. Natl. Acad. Sci. USA 1997, 94, 7362–7366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westoby, M.; Falster, D.S.; Moles, A.T.; Vesk, P.A.; Wright, I.J. Plant ecological strategies: Some leading dimensions of variation between species. Annu. Rev. Ecol. Syst. 2002, 33, 125–159. [Google Scholar] [CrossRef] [Green Version]
- Evans, J.R. Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia 1989, 78, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Lei, Z.; Rondon, S.I.; Gao, Y. Potential of a strain of Beauveria bassiana (Hypocreales: Cordycipitaceae) for the control of the potato tuberworm, Phthorimaea operculella (Zeller). Int. J. Pest Manag. 2017, 63, 352–354. [Google Scholar] [CrossRef]
- Rondon, S.I. The potato tuberworm: A lierature review of its biology, ecology, and control. Am. J. Potato Res. 2010, 87, 149–166. [Google Scholar] [CrossRef] [Green Version]
- Friedman, M. Potato glycoalkaloids and metabolites: Roles in the plant and in the diet. J. Agric. Food Chem. 2006, 54, 8655–8681. [Google Scholar] [CrossRef]
- Horgan, F.G.; Quiring, D.T.; Lagnaoui, A.; Salas, A.R.; Pelletier, Y. Periderm- and cortex-based resistance to tuber-feeding Phthorimaea operculella in two wild potato species. Entomol. Exp. Appl. 2007, 125, 249–258. [Google Scholar] [CrossRef]
- Rondon, S.I.; Hane, D.C.; Brown, C.R.; Vales, M.I.; Dogramaci, M. Resistance of potato germplasm to the potato tuberworm (Lepidoptera: Gelechiidae). J. Econ. Entomol. 2009, 102, 1649–1653. [Google Scholar] [CrossRef]
- Horgan, F.G.; Quiring, D.T.; Lagnaoui, A.; Salas, A.R.; Pelletier, Y. Variations in resistance against Phthorimaea operculella in wild potato tubers. Entomol. Exp. Appl. 2010, 137, 269–279. [Google Scholar] [CrossRef]
- Horgan, F.G.; Quiring, D.T.; Lagnaoui, A.; Pelletier, Y. Tuber production, dormancy and resistance against Phthorimaea operculella (Zeller) in wild potato species. J. Appl. Entomol. 2013, 137, 739–750. [Google Scholar] [CrossRef]
- Alipour, V.; Mehrkhou, F. Effects of different potato cultivars on life history and demographic parameters of Phthorimaea operculella Zeller (Lepidoptera: Gelechiidae). Int. J. Pest Manag. 2018, 64, 365–371. [Google Scholar] [CrossRef]
- Malakar-Kuenen, R.; Tingey, W.M. Aspects of tuber resistance in hybrid potatoes to potato tuber worm. Entomol. Exp. Appl. 2006, 120, 131–137. [Google Scholar] [CrossRef]
- Kumar, P.; Garrido, E.; Zhao, K.; Zheng, Y.; Alseekh, S.; Vargas-Ortiz, E.; Fernie, A.R.; Fei, Z.; Poveda, K.; Jander, G. Tecia solanivora infestation increases tuber starch accumulation in Pastusa Suprema potatoes. J. Integr. Plant Biol. 2018, 60, 1083–1096. [Google Scholar] [CrossRef] [PubMed]
- Elleuch, M.; Besbes, S.; Roiseux, O.; Blecker, C.; Attia, H. Quality characteristics of sesame seeds and by-products. Food Chem. 2007, 103, 641–650. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Sotelo, A.; Serrano, B. High-performance liquid chromatographic determination of the glycoalkaloids alpha-solanine and alpha-chaconine in 12 commercial varieties of Mexican potato. J. Agric. Food Chem. 2000, 48, 2472–2475. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M.; Roitman, J.N.; Kozukue, N. Glycoalkaloid and calystegine contents of eight potato cultivars. J. Agric. Food Chem. 2003, 51, 2964–2973. [Google Scholar] [CrossRef]
- Friedman, M.; McDonald, G.M. Potato glycoalkaloids: Chemistry, analysis, safety, and plant physiology. Crit. Rev. Plant Sci. 1997, 16, 55–132. [Google Scholar] [CrossRef]
- R Development Core Team. R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Osman, S.F. Glycoalkaloids of the Solanaceae. Recent Adv. Phytochem. 1980, 14, 75–96. [Google Scholar]
- Hlywka, J.J.; Stephenson, G.R.; Sears, M.K.; Yada, R.Y. Effects of insect damage on glycoalkaloid content in potatoes (Solanum tubersoum). J. Agric. Food Chem. 1994, 42, 2545–2550. [Google Scholar] [CrossRef]
- Pariera Dinkins, C.L.; Peterson, R.K.; Gibson, J.E.; Hu, Q.; Weaver, D.K. Glycoalkaloid responses of potato to Colorado potato beetle defoliation. Food Chem. Toxicol. 2008, 46, 2832–2836. [Google Scholar] [CrossRef] [PubMed]
- Phillips, B.J.; Hughes, J.A.; Phillips, J.C.; Walters, D.G.; Anderson, D.; Tahourdin, C.S. A study of the toxic hazard that might be associated with the consumption of green potato tops. Food Chem. Toxicol. 1996, 34, 439–448. [Google Scholar] [CrossRef]
- Berenbaum, M.R. Turnabout is fair play: Secondary roles for primary compounds. J. Chem. Ecol. 1995, 21, 925–940. [Google Scholar] [CrossRef] [PubMed]
- Awmack, C.S.; Leather, S.R. Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol. 2002, 47, 817–844. [Google Scholar] [CrossRef] [PubMed]
- Ishaaya, I.; Moore, I.; Joseph, D. Protease and amylase activity in larvae of the Egyptian cotton worm, Spodoptra littoralis. J. Insect Physiol. 1971, 17, 945–953. [Google Scholar] [CrossRef]
- Wold, E.N.; Marquis, R.J. Induced defense in white oak: Effects on herbivores and consequences for the plant. Ecology 1997, 78, 1356–1369. [Google Scholar] [CrossRef]
- Golizadeh, A.; Esmaeili, N. Comparative life history and fecundity of Phthorimaea operculella (Lepidoptera: Gelechiidae) on leaves and tubers of different potato cultivars. J. Econ. Entomol. 2012, 105, 1809–1815. [Google Scholar] [CrossRef] [PubMed]
- Fenemore, P.G. Oviposition of potato tuber moth, Phthorimaea operculella Zell. (Lepidoptera: Gelechiidae); the influence of adult food, pupal weight, and host-plant tissue on fecundity. N. Z. J. Zool. 1979, 7, 389–395. [Google Scholar] [CrossRef]
- Sonnemann, I.; Buchholz, Y.; Rautenberg, D.; Wurst, S. Land use intensity modulates the impact of root herbivores on plant interactions with above- and below-ground organisms. Agric. Ecosyst. Environ. 2016, 219, 26–31. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Treatment | Sex | Treatment × Sex | |||
|---|---|---|---|---|---|---|
| F2,66 | p | F1,67 | p | F5,63 | p | |
| Laval developmental time (days) | 0.759 | 0.472 | 1.882 | 0.175 | 3.240 | 0.046 |
| Pupal developmental time (days) | 0.800 | 0.454 | 0.082 | 0.776 | 2.546 | 0.086 |
| Pupal weight (mg) | 1.846 | 0.166 | 69.599 | <0.001 | 2.986 | 0.058 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, D.; Wang, Q.; Sun, X.; Gao, Y.; Ding, J. Potato Tuberworm Phthorimaea operculella (Zeller) (Lepidoptera: Gelechioidea) Leaf Infestation Affects Performance of Conspecific Larvae on Harvested Tubers by Inducing Chemical Defenses. Insects 2020, 11, 633. https://doi.org/10.3390/insects11090633
Wang D, Wang Q, Sun X, Gao Y, Ding J. Potato Tuberworm Phthorimaea operculella (Zeller) (Lepidoptera: Gelechioidea) Leaf Infestation Affects Performance of Conspecific Larvae on Harvested Tubers by Inducing Chemical Defenses. Insects. 2020; 11(9):633. https://doi.org/10.3390/insects11090633
Chicago/Turabian StyleWang, Dingli, Qiyun Wang, Xiao Sun, Yulin Gao, and Jianqing Ding. 2020. "Potato Tuberworm Phthorimaea operculella (Zeller) (Lepidoptera: Gelechioidea) Leaf Infestation Affects Performance of Conspecific Larvae on Harvested Tubers by Inducing Chemical Defenses" Insects 11, no. 9: 633. https://doi.org/10.3390/insects11090633
APA StyleWang, D., Wang, Q., Sun, X., Gao, Y., & Ding, J. (2020). Potato Tuberworm Phthorimaea operculella (Zeller) (Lepidoptera: Gelechioidea) Leaf Infestation Affects Performance of Conspecific Larvae on Harvested Tubers by Inducing Chemical Defenses. Insects, 11(9), 633. https://doi.org/10.3390/insects11090633