Slug Monitoring and Impacts on the Ground Beetle Community in the Frame of Sustainable Pest Control in Conventional and Conservation Agroecosystems

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Trial No. 1: Efficacy of Traps and Refuges in Monitoring Slugs and Ground Beetles in Conventional and Conservation Agroecosystems
2.2. Trial No. 2: Efficacy of Traps and Refuges in Monitoring Slugs and Ground Beetles in a No-Till Field
2.3. Data Collection and Species Identification
2.4. DNA Barcoding
2.5. Statistical Analysis
3. Results
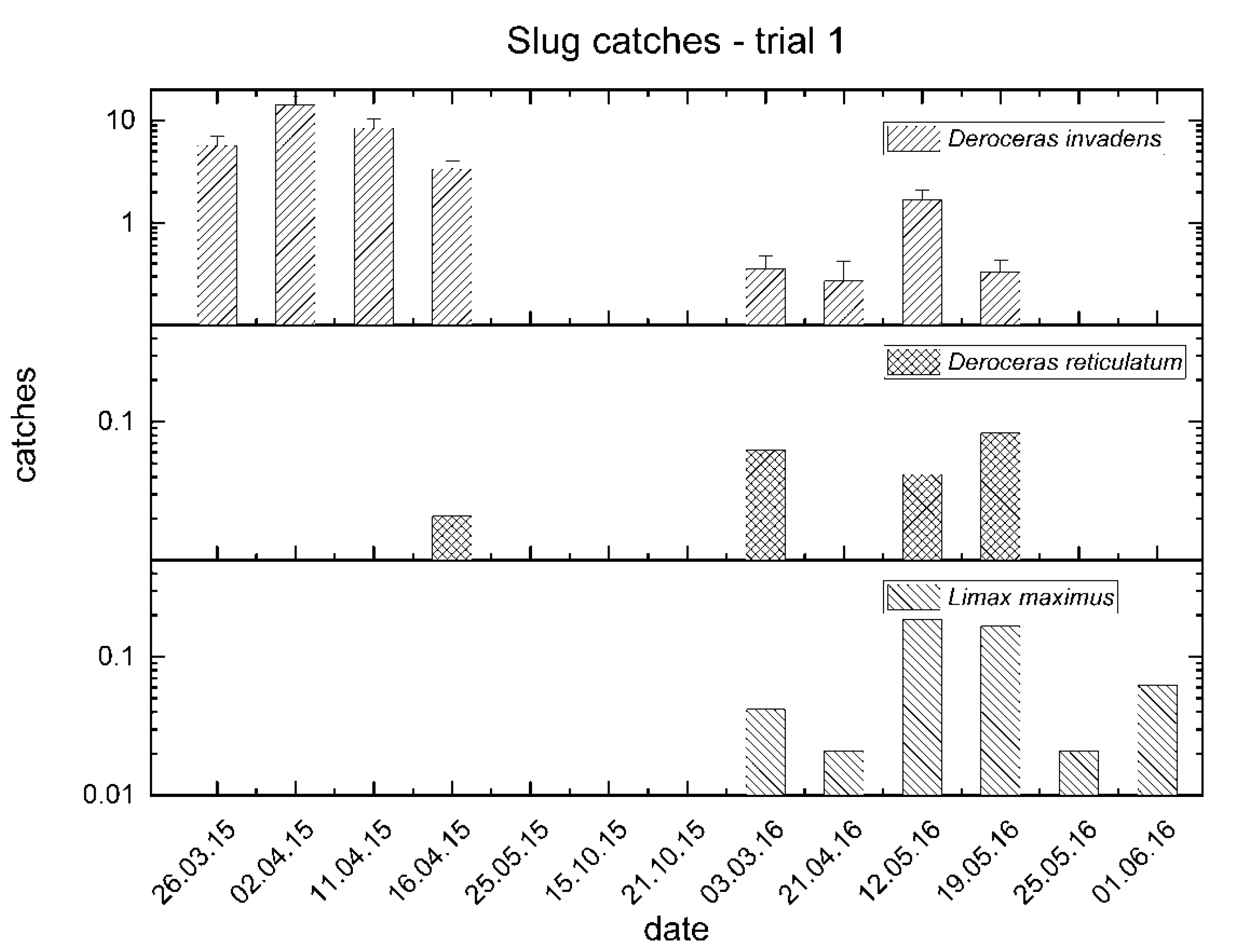
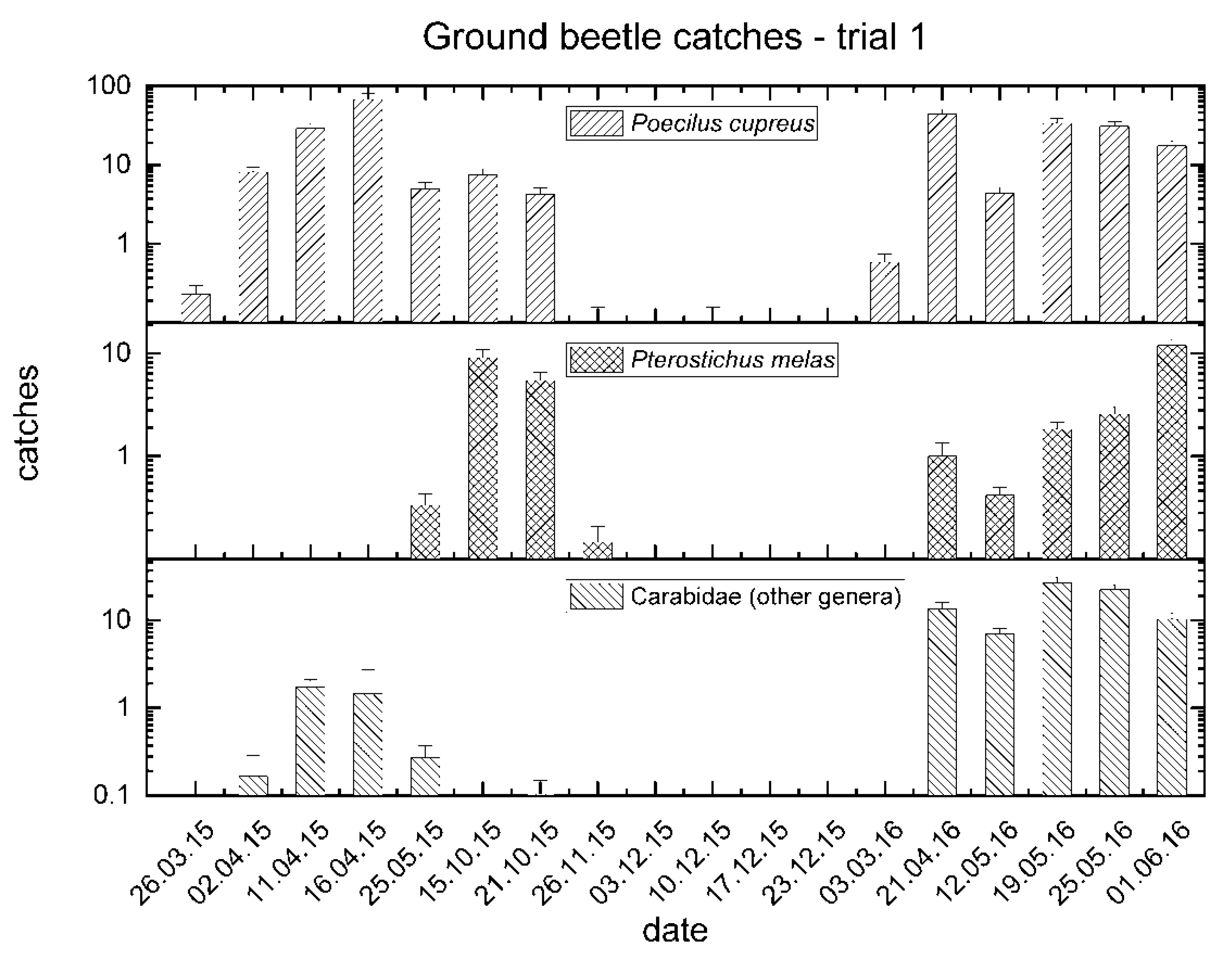
3.1. Trial No. 1
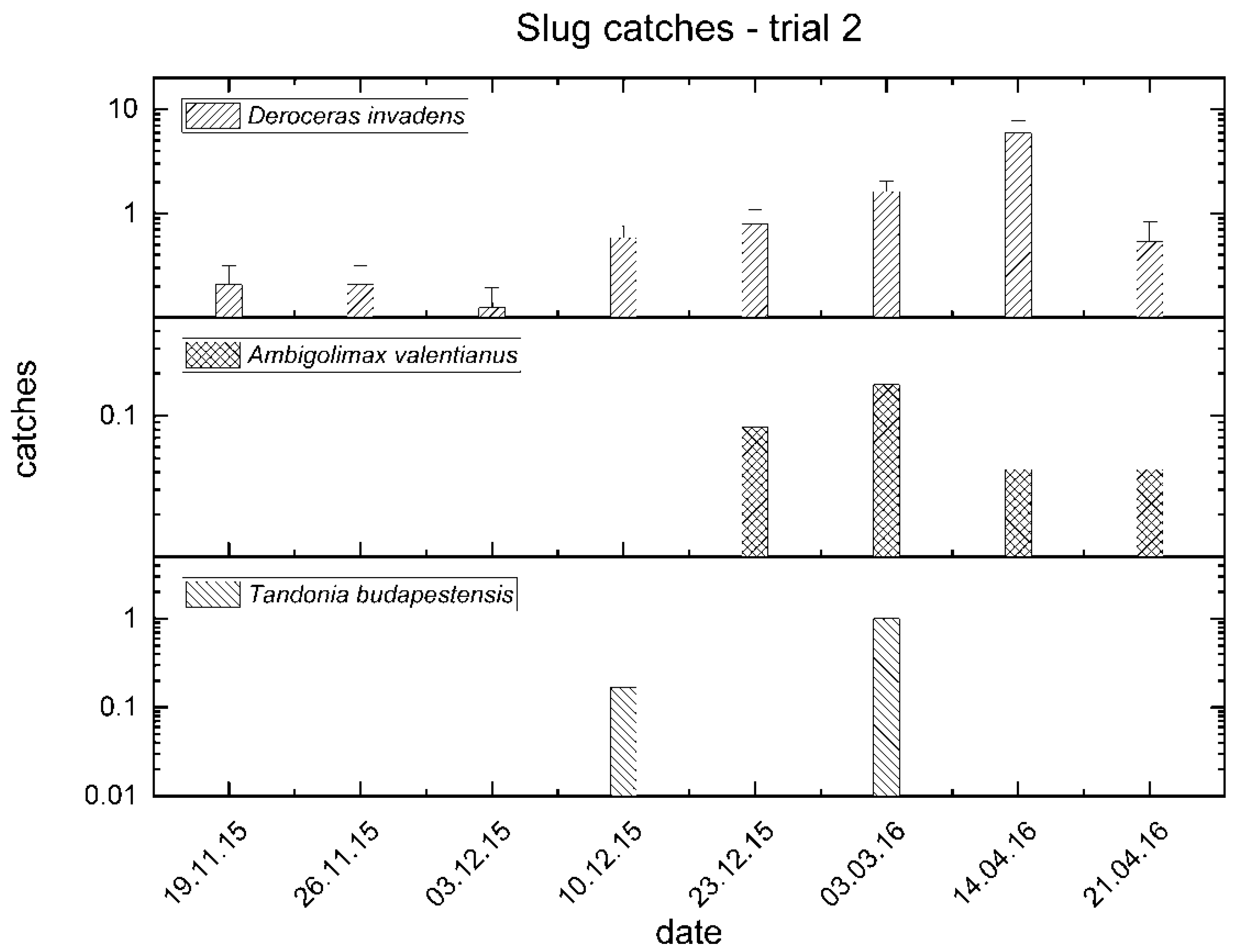
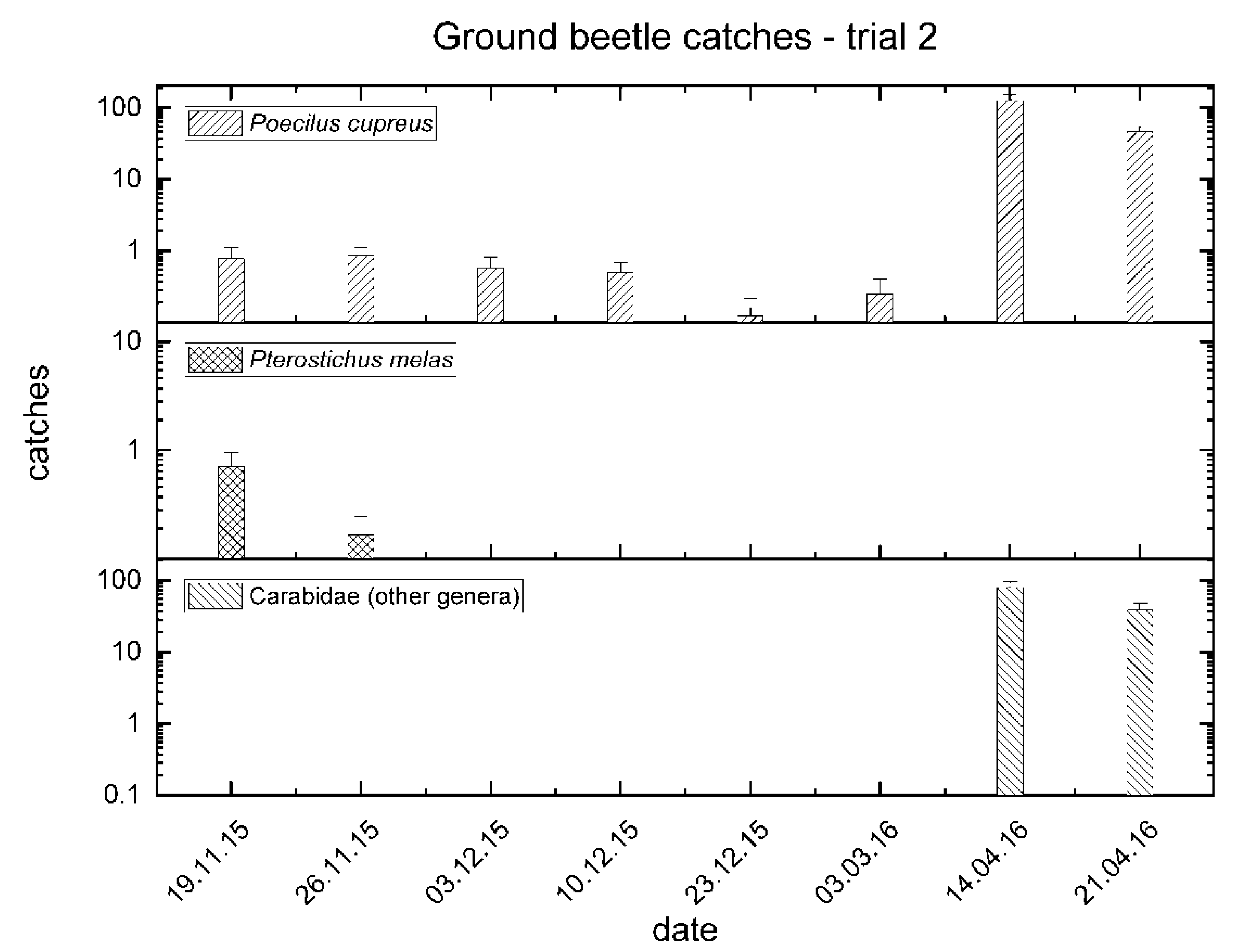
3.2. Trial No. 2
3.3. Slug DNA Barcoding
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lal, R. A system approach to conservation agriculture. J. Soil Water Conserv. 2015, 70, 82A–88A. [Google Scholar] [CrossRef] [Green Version]
- Ranaivoson, L.; Naudin, K.; Ripoche, A.; Affholder, F.; Rabeharisoa, L.; Corbeels, M. Agro-ecological functions of crop residues under conservation agriculture. A review. Agron. Sustain. Dev. 2017, 37, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Tabaglio, V.; Gavazzi, C.; Menta, C. Physico-chemical indicators and microarthropod communities as influenced by no-till, conventional tillage and nitrogen fertilisation after four years of continuous maize. Soil Tillage Res. 2009, 105, 135–142. [Google Scholar] [CrossRef]
- Palm, C.; Blanco-Canqui, H.; DeClerck, F.; Gatere, L.; Grace, P. Conservation agriculture and ecosystem services: An overview. Agric. Ecosyst. Environ. 2014, 187, 87–105. [Google Scholar] [CrossRef] [Green Version]
- Tamburini, G.; De Simone, S.; Sigura, M.; Boscutti, F.; Marini, L. Soil management shapes ecosystem service provision and trade-offs in agricultural landscapes. Proc. R. Soc. B Biol. Sci. 2016, 283, 20161369. [Google Scholar] [CrossRef] [Green Version]
- Stinner, B.R.; House, G.J. Arthropods and other invertebrates in conservation-tillage agriculture. Annu. Rev. Entomol. 1990, 35, 299–318. [Google Scholar] [CrossRef]
- Soane, B.D.; Ball, B.C.; Arvidsson, J.; Basch, G.; Moreno, F.; Roger-Estrade, J. No-till in northern, western and south-western Europe: A review of problems and opportunities for crop production and the environment. Soil Tillage Res. 2012, 118, 66–87. [Google Scholar] [CrossRef] [Green Version]
- Roos, D.; Caminero Saldaña, C.; Arroyo, B.; Mougeot, F.; Luque-Larena, J.; Lambin, X. Unintentional effects of environmentally-friendly farming practices: Arising conflicts between zero-tillage and a crop pest, the common vole (Microtus arvalis). Agric. Ecosyst. Environ. 2019, 272, 105–113. [Google Scholar] [CrossRef]
- Nyamwasa, I.; Zhang, S.; Sun, X.; Yin, J.; Li, X.; Qin, J.; Li, J.; Li, K. Transition challenge to organic agriculture: A course for advancing belowground insect pest management. Appl. Soil Ecol. 2020, 148, 103476. [Google Scholar] [CrossRef]
- Basch, G.; Friedrich, T.; Kassam, A.; Gonzalez-Sanchez, E. Conservation agriculture in Europe. In Conservation Agriculture; Farooq, M., Siddique, K.H.M., Eds.; Springer International Publishing: Cham, Switzerland, 2015; pp. 357–389. [Google Scholar] [CrossRef]
- Leake, A.R. Integrated pest management for conservation agriculture. In Conservation Agriculture: Environment, Farmers Experiences, Innovations, Socio-Economy, Policy; García-Torres, L., Benites, J., Martínez-Vilela, A., Holgado-Cabrera, A., Eds.; Springer: Dordrecht, The Netherlands, 2003; pp. 271–279. [Google Scholar] [CrossRef]
- Hammond, R.B.; Beck, T.; Smith, J.A.; Amos, R.; Barker, J.; Moore, R.; Siegrist, H.; Slates, D.; Ward, B. Slugs in conservation tillage corn and soybeans in the Eastern Corn Belt. J. Entomol. Sci. 1999, 34, 467–478. [Google Scholar] [CrossRef]
- Douglas, M.R.; Tooker, J.F. Slug (Mollusca: Agriolimacidae, Arionidae) ecology and management in no-till field crops, with an emphasis on the mid-Atlantic region. J. Integr. Pest Manag. 2012, 3, C1–C9. [Google Scholar] [CrossRef] [Green Version]
- Kladivko, E.J. Tillage systems and soil ecology. Soil Tillage Res. 2001, 61, 61–76. [Google Scholar] [CrossRef]
- Stavi, I.; Bel, G.; Zaady, E. Soil functions and ecosystem services in conventional, conservation, and integrated agricultural systems. A review. Agron. Sustain. Dev. 2016, 36, 32. [Google Scholar] [CrossRef] [Green Version]
- Nichols, V.; Verhulst, N.; Cox, R.; Govaerts, B. Weed dynamics and conservation agriculture principles: A review. Field Crop. Res. 2015, 183, 56–68. [Google Scholar] [CrossRef] [Green Version]
- Cook, R.T.; Bailey, S.E.R.; McCrohan, C.R. The potential for common weeds to reduce slug damage to winter wheat: Laboratory and fields studies. J. Appl. Ecol. 1997, 34, 79–87. [Google Scholar] [CrossRef]
- Frank, T.; Barone, M. Short-term field study on weeds reducing slug feeding on oilseed rape. Z. Pflanzenkrankh. Pflanzenschutz J. Plant Dis. Prot. 1999, 106, 534–538. [Google Scholar]
- Kozłowski, J.; Kozłowska, M. Weeds as a supplementary or alternative food for Arion lusitanicus Mabille (Gastropoda: Stylommatophora). J. Conchol. 2000, 37, 75–79. [Google Scholar]
- Brooks, A.S.; Wilcox, A.; Cook, R.T.; Crook, M.J. A laboratory-based comparison of a molluscicide and an alternative food source (red clover) as means of reducing slug damage to winter wheat. Pest Manag. Sci. 2005, 61, 715–720. [Google Scholar] [CrossRef]
- Glen, D.M. The effects of cultural measures on cereal pests and their role in integrated pest management. Integr. Pest Manag. Rev. 2000, 5, 25–40. [Google Scholar] [CrossRef]
- Godan, D. Pest Slugs and Snails-Biology and Control; Springer: Heidelberg, Germany, 1983; pp. 1–448. [Google Scholar] [CrossRef]
- Vernava, M.N.; Phillips-Aalten, P.M.; Hughes, L.A.; Rowcliffe, H.; Wiltshire, C.W.; Glen, D.M. Influences of preceding cover crops on slug damage and biological control using Phasmarhabditis hermaphrodita. Ann. Appl. Biol. 2004, 145, 279–284. [Google Scholar] [CrossRef]
- House, G.J.; Alzugaray, M.D.R. Influence of cover cropping and no-tillage practices on community composition of soil arthropods in a North Carolina agroecosystem. Environ. Entomol. 1989, 18, 302–307. [Google Scholar] [CrossRef]
- Willson, H.R.; Eisley, J.B. Effects of tillage and prior crop on the incidence of five key pests on Ohio corn. J. Econ. Entomol. 1992, 85, 853–859. [Google Scholar] [CrossRef]
- Le Gall, M.; Tooker, J.F. Developing ecologically based pest management programs for terrestrial molluscs in field and forage crops. J. Pest Sci. 2017, 90, 825–838. [Google Scholar] [CrossRef]
- Rowen, E.K.; Regan, K.H.; Barbercheck, M.E.; Tooker, J.F. Is tillage beneficial or detrimental for insect and slug management? A meta-analysis. Agric. Ecosyst. Environ. 2020, 294, 105–113. [Google Scholar] [CrossRef]
- Symondson, W.O.C.; Glen, D.M.; Wiltshire, C.W.; Langdon, C.J.; Liddell, J.E. Effects of cultivation techniques and methods of straw disposal on predation by Pterostichus melanarius (Coleoptera: Carabidae) upon slugs (Gastropoda: Pulmonata) in an arable field. J. Appl. Ecol. 1996, 33, 741–753. [Google Scholar] [CrossRef]
- Kennedy, T.F.; Connery, J.; Fortune, T.; Forristal, D.; Grant, J. A comparison of the effects of minimum-till and conventional-till methods, with and without straw incorporation, on slugs, slug damage, earthworms and carabid beetles in autumn-sown cereals. J. Agric. Sci. 2013, 151, 605–629. [Google Scholar] [CrossRef]
- Langmaack, M.; Land, S.; Büchs, W. Effects of different field management systems on the carabid coenosis in oil seed rape with special respect to ecology and nutritional status of predacious Poecilus cupreus L. (Col., Carabidae). J. Appl. Entomol. 2001, 125, 313–320. [Google Scholar] [CrossRef]
- Kosewska, A.; Skalski, T.; Nietupski, M. Effect of conventional and non-inversion tillage systems on the abundance and some life history traits of carabid beetles (Coleoptera: Carabidae) in winter triticale fields. Eur. J. Entomol. 2014, 111, 669–676. [Google Scholar] [CrossRef]
- Barzman, M.; Bàrberi, P.; Birch, A.N.; Boonekamp, P.; Dachbrodt-Saaydeh, S.; Graf, B.; Hommel, B.; Jensen, J.E.; Kiss, J.; Kudsk, P.; et al. Eight principles of integrated pest management. Agron. Sustain. Dev. 2015, 35, 1199–1215. [Google Scholar] [CrossRef]
- Glen, D.M.; Milsom, N.F.; Wiltshire, C.W. Effect of seed depth on slug damage to winter wheat. Ann. Appl. Biol. 1990, 117, 693–701. [Google Scholar] [CrossRef]
- Jordan, V.W.L.; Hutcheon, J.A.; Donaldson, G.V.; Farmer, D.P. Research into and development of integrated farming systems for less-intensive arable crop production: Experimental progress (1989–1994) and commercial implementation. Agric. Ecosyst. Environ. 1997, 64, 141–148. [Google Scholar] [CrossRef]
- Glen, D.M.; Spaull, A.M.; Mowat, D.J.; Green, D.B.; Jackson, A.W. Crop monitoring to assess the risk of slug damage to winter wheat in the United Kingdom. Ann. Appl. Biol. 1993, 122, 161–172. [Google Scholar] [CrossRef]
- Binns, M.R.; Nyrop, J.P. Sampling insect populations for the purpose of IPM decision making. Annu. Rev. Entomol. 1992, 37, 427–453. [Google Scholar] [CrossRef]
- Castle, S.; Naranjo, S.E. Sampling plans, selective insecticides and sustainability: The case for IPM as ‘informed pest management’. Pest Manag. Sci. 2009, 65, 1321–1328. [Google Scholar] [CrossRef] [PubMed]
- Wiktor, A. Agriolimacidae (Gastropoda: Pulmonata)-a systematic monograph. Ann. Zool. 2000, 49, 347–590. [Google Scholar]
- Wiktor, A.; De-Niu, C.; Ming, W. Stylommatophoran slugs of China (Gastropoda: Pulmonata)-Prodromus. Folia Malacol. 2000, 8, 3–35. [Google Scholar] [CrossRef]
- Rowson, B.; Turner, J.A.; Anderson, R.; Symondson, W.O.C. Slugs of Britain and Ireland–Identification, Understanding and Control; Field Studies Council-National Museum of Wales: Telford, UK, 2014; pp. 1–136. [Google Scholar]
- Hůrka, K. Carabidae of the Czech and Slovak Republics; Ing. Vit Kabourek: Zlin, Czech Republic, 1996; pp. 1–565. [Google Scholar]
- Gutiérrez Gregoric, D.E.; Beltramino, A.A.; Vogler, R.E.; Cuezzo, M.G.; Núñez, V.; Gomes, S.R.; Virgillito, M.; Miquel, S.E. First records of four exotic slugs in Argentina. Am. Malacol. Bull. 2013, 31, 245–256. [Google Scholar] [CrossRef] [Green Version]
- Rowson, B.; Anderson, R.; Turner, J.A.; Symondson, W.O.C. The Slugs of Britain and Ireland: Undetected and undescribed species increase a well-studied, economically important fauna by more than 20%. PLoS ONE 2014, 9, e91907. [Google Scholar] [CrossRef] [Green Version]
- Panini, M.; Dradi, D.; Marani, G.; Butturini, A.; Mazzoni, E. Detecting the presence of target-site resistance to neonicotinoids and pyrethroids in Italian populations of Myzus persicae. Pest Manag. Sci. 2014, 70, 931–938. [Google Scholar] [CrossRef]
- Hebert, P.D.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Thomas, R.S.; Glen, D.M.; Symondson, W.O.C. Prey detection through olfaction by the soil-dwelling larvae of the carabid predator Pterostichus melanarius. Soil Biol. Biochem. 2008, 40, 207–216. [Google Scholar] [CrossRef]
- Oberholzer, F.; Escher, N.; Frank, T. The potential of carabid beetles (Coleoptera) to reduce slug damage to oilseed rape in the laboratory. Eur. J. Entomol. 2003, 100, 81–85. [Google Scholar] [CrossRef] [Green Version]
- Oberholzer, F.; Frank, T. Predation by the carabid beetles Pterostichus melanarius and Poecilus cupreus on slugs and slug eggs. Biocontrol Sci. Technol. 2003, 13, 99–110. [Google Scholar] [CrossRef]
- Ribera, I.; Foster, G.N.; Downie, I.S.; McCracken, D.I.; Abernethy, V.J. A comparative study of the morphology and life traits of Scottish ground beetles (Coleoptera, Carabidae). Ann. Zool. Fennici 1999, 36, 21–37. [Google Scholar]
- Zaller, J.; Moser, D.; Drapela, T.; Frank, T. Ground-dwelling predators can affect within-field pest insect emergence in winter oilseed rape fields. Biocontrol 2009, 54, 247–253. [Google Scholar] [CrossRef]
- Holland, J.M. The Agroecology of Carabid Beetles; Intercept Ldt: Andover, UK, 2002; pp. 1–356. [Google Scholar]
- Luff, M.L. Biology of polyphagous ground beetles in agriculture. Agric. Zool. Rev. 1987, 2, 237–278. [Google Scholar]
- Honek, A.; Martinkova, Z.; Jarosik, V. Ground beetles (Carabidae) as seed predators. Eur. J. Entomol. 2003, 100, 531–544. [Google Scholar] [CrossRef] [Green Version]
- Talarico, F.; Giglio, A.; Pizzolotto, R.; Brandmayr, P. A synthesis of feeding habits and reproduction rhythm in Italian seed-feeding ground beetles (Coleoptera: Carabidae). Eur. J. Entomol. 2016, 113, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Pretorius, R.J.; Hein, G.L.; Blankenship, E.E.; Purrington, F.F.; Wilson, R.G.; Bradshaw, J.D. Comparing the effects of two tillage operations on beneficial epigeal arthropod communities and their associated ecosystem services in sugar beets. J. Econ. Entomol. 2018, 111, 2617–2631. [Google Scholar] [CrossRef] [Green Version]
- Tabaglio, V. (Università Cattolica del Sacro Cuore, Piacenza, Italy). Personal communication, 2014. [Google Scholar]
- Hutchinson, J.; Reise, H.; Robinson, D. A biography of an invasive terrestrial slug: The spread, distribution and habitat of Deroceras invadens. NeoBiota 2014, 23, 17–64. [Google Scholar] [CrossRef] [Green Version]
- Araiza-Gómez, V.; Naranjo-García, E.; Zúñiga, G. The exotic slugs of the genus Deroceras (Agriolimacidae) in Mexico: Morphological and molecular characterization, and new data on their distribution. Am. Malacol. Bull. 2017, 35, 126–133. [Google Scholar] [CrossRef]
- Reise, H.; Hutchinson, J.M.C.; Robinson, D.G. Two introduced pest slugs: Tandonia budapestensis new to the Americas, and Deroceras panormitanum new to the eastern USA. Veliger 2006, 48, 110–115. [Google Scholar]
- Wiktor, A. Milacidae (Gastropoda, Pulmonata)-systematic monograph. Ann. Zool. 1987, 41, 153–319. [Google Scholar]
- Vendetti, J.E.; Burnett, E.; Carlton, L.; Curran, A.T.; Lee, C.; Matsumoto, R.; Mc Donnell, R.; Reich, I.; Willadsen, O. The introduced terrestrial slugs Ambigolimax nyctelius (Bourguignat, 1861) and Ambigolimax valentianus (Férussac, 1821) (Gastropoda: Limacidae) in California, with a discussion of taxonomy, systematics, and discovery by citizen science. J. Nat. Hist. 2019, 53, 1607–1632. [Google Scholar] [CrossRef]
- Molinari, F.; Pradelli, G. Carabid population in a crop rotation system in northern Italy. Boll. Zool. Agrar. Bachic. Ser. II 1995, 27, 163–175. [Google Scholar]
- Tuf, I.H.; Dedek, P.; Veselý, M. Does the diurnal activity pattern of carabid beetles depend on season, ground temperature and habitat? Arch. Biol. Sci. 2012, 64, 721–732. [Google Scholar] [CrossRef]
- Fusser, M.S.; Pfister, S.C.; Entling, M.H.; Schirmel, J. Effects of landscape composition on carabids and slugs in herbaceous and woody field margins. Agric. Ecosyst. Environ. 2016, 226, 79–87. [Google Scholar] [CrossRef]
- Honek, A. The effect of temperature on the activity of Carabidae (Coleoptera) in a fallow field. Eur. J. Entomol. 1997, 94, 97–104. [Google Scholar]
- Kulagowski, R.; Riggi, L.; Chailleux, A. Short-term effects of conversion to direct seeding mulch-based cropping systems on macro-fauna and weed dynamics. J. Crop Improv. 2016, 30, 65–83. [Google Scholar] [CrossRef]
- Lemic, D.; Čačija, M.; Gašparić, H.V.; Drmić, Z.; Bažok, R.; Živković, I.P. The ground beetle (Coleoptera: Carabidae) community in an intensively managed agricultural landscape. Appl. Ecol. Environ. Res. 2017, 15, 661–674. [Google Scholar] [CrossRef]
- Mair, J.; Port, G.R. Predation by the carabid beetles Pterostichus madidus and Nebria brevicollis is affected by size and condition of the prey slug Deroceras reticulatum. Agric. For. Entomol. 2001, 3, 99–106. [Google Scholar] [CrossRef]
- Paill, W. Slug feeding in the carabid beetle Pterostichus melanarius: Seasonality and dependence on prey size. J. Molluscan Stud. 2004, 70, 203–205. [Google Scholar] [CrossRef] [Green Version]
- Hatteland, B.A.; Grutle, K.; Mong, C.E.; Skartveit, J.; Symondson, W.O.C.; Solhøy, T. Predation by beetles (Carabidae, Staphylinidae) on eggs and juveniles of the Iberian slug Arion lusitanicus in the laboratory. Bull. Entomol. Res. 2010, 100, 559–567. [Google Scholar] [CrossRef]
- Young, A.G.; Port, G.R.; Craig, A.D.; James, D.A.; Green, T. The use of refuge traps in assessing risk of slug damage: A comparison of trap material and bait. In Slug and Snail Pests in Agriculture; Henderson, L.F., Ed.; British Crop Protection Council: Farnham, UK, 1996; pp. 133–140. [Google Scholar]
- Taylor, J.W. Monograph of the Land and Freshwater Mollusca of the British Isles. 2: Testacellidae, Limacidae, Arionidae; Taylor Bros: Leeds, UK, 1907. [Google Scholar]
- Selim, S.I. The Control of Snail and Slug Pests in Agriculture. Part I. Analysis for and Dissipation of Metaldehyde in Vegetable Crops; Part II. Beer as a Slug Attractant. Ph.D. Thesis, University of California, Davis, CA, USA, 1974. [Google Scholar]
- Piechowicz, B.; Watrakiewicz, R.; Rębisz, E.; Zaręba, L.; Balawejder, M.; Pieniążek, M.; Zwolak, A.; Grodzicki, P.; Sadło, S. Beer as attractant for Arion vulgaris Moquin-Tandon, 1885 (Gastropoda: Pulmonata: Arionidae). Folia Malacol. 2016, 24, 193–200. [Google Scholar] [CrossRef]
- Smith, F.F.; Boswell, A.L. New baits and attractants for slugs. J. Econ. Entomol. 1970, 63, 1919–1922. [Google Scholar] [CrossRef]
- Cravedi, P.; Molinari, F.; Mazzoni, E. Effetti dei cambiamenti ambientali sulla fauna selvatica e sull’entomofauna dei sistemi agricoli e forestali. Ital. J. Agron. 2010, 5, 23–27. [Google Scholar]
- Bohan, D.A.; Bohan, A.C.; Glen, D.M.; Symondson, W.O.C.; Wiltshire, C.W.; Hughes, L. Spatial dynamics of predation by carabid beetles on slugs. J. Anim. Ecol. 2000, 69, 367–379. [Google Scholar] [CrossRef]
- Knapp, M.; Baranovská, E.; Jakubec, P. Effects of bait presence and type of preservative fluid on ground and carrion beetle samples collected by pitfall trapping. Environ. Entomol. 2016, 45, 1022–1028. [Google Scholar] [CrossRef]
- Baini, F.; Del Vecchio, M.; Vizzari, L.; Zapparoli, M. Can the efficiency of pitfall traps in collecting arthropods vary according to the used mixtures as bait? Rend. Lincei 2016, 27, 495–499. [Google Scholar] [CrossRef]
- Schipanski, M.E.; Barbercheck, M.; Douglas, M.R.; Finney, D.M.; Haider, K.; Kaye, J.P.; Kemanian, A.R.; Mortensen, D.A.; Ryan, M.R.; Tooker, J.; et al. A framework for evaluating ecosystem services provided by cover crops in agroecosystems. Agric. Syst. 2014, 125, 12–22. [Google Scholar] [CrossRef]
- Hof, A.R.; Bright, P.W. The impact of grassy field margins on macro-invertebrate abundance in adjacent arable fields. Agric. Ecosyst. Environ. 2010, 139, 280–283. [Google Scholar] [CrossRef]
- Rossi, E.; Antichi, D.; Loni, A.; Canovai, R.; Sbrana, M.; Mazzoncini, M. Ground beetle (Coleoptera: Carabidae) assemblages and slug abundance in agricultural fields under organic and low-input conventional management within a long-term agronomic trial in Central Italy. Environ. Entomol. 2019, 48, 1377–1387. [Google Scholar] [CrossRef] [PubMed]
- Greenslade, P.J.M. Pitfall trapping as a method for studying populations of Carabidae (Coleoptera). J. Anim. Ecol. 1964, 33, 301–310. [Google Scholar] [CrossRef]
- Pearce, J.L.; Venier, L.A.; McKee, J.; Pedlar, J.; McKenney, D. Influence of habitat and microhabitat on carabid (Coleoptera: Carabidae) assemblages in four stand types. Can. Entomol. 2003, 135, 337–357. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Management | D. invadens | P. cupreus | P. melas | Carabidae (Other Genera) |
|---|---|---|---|---|
| CONV-Plough 1 | 4.9 (±1.1) b | 11.7 (±2.2) a | 2.3 (±0.4) a | 4.7 (±0.9) a |
| CONS-Rye 2 | 3.3 (±0.8) ab | 20.4 (±2.7) b | 2.2 (±0.4) a | 5.6 (±1.1) a |
| CONS-Vetch 3 | 1.2 (±0.2) a | 27.0 (±4.1) c | 2.1 (±0.4) a | 9.1 (±1.5) b |
| CONS-Mix 4 | 1.2 (±0.2) a | 18.9 (±2.8) b | 3.5 (±0.7) a | 7.4 (±1.4) ab |
| Trap Type | D. invadens | P. cupreus | P. melas | Carabidae (Other Genera) |
|---|---|---|---|---|
| wooden boards | 0.6 (±0.1) a | 0.1 (±0.0) a | 0.0 (±0.0) a | 0.0 (±0.0) a |
| beer A | 3.4 (±0.6) b | 27.1 (±3.0) b | 3.3 (±0.4) b | 9.1 (±1.2) b |
| beer B | 4.0 (±0.8) b | 31.3 (±3.1) b | 4.3 (±0.6) c | 10.9 (±1.3) b |
| Trap Type | D. invadens | P. cupreus | P. melas | Carabidae (Other Genera) |
|---|---|---|---|---|
| wooden board | 0.6 (±0.1) a | 0.0 (±1.1) a | 0.00 (±0.0) a | 0.0 (±0.0) a |
| beer A | 1.5 (±0.6) a | 29.1 (±8.0) b | 0.16 (±0.1) ab | 20.2 (±5.3) b |
| beer B | 1.6 (±0.6) a | 36.7 (±10.0) b | 0.22 (±0.1) b | 24.1 (±6.8) b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Scaccini, D.; Panini, M.; Chiesa, O.; Nicoli Aldini, R.; Tabaglio, V.; Mazzoni, E. Slug Monitoring and Impacts on the Ground Beetle Community in the Frame of Sustainable Pest Control in Conventional and Conservation Agroecosystems. Insects 2020, 11, 380. https://doi.org/10.3390/insects11060380
Scaccini D, Panini M, Chiesa O, Nicoli Aldini R, Tabaglio V, Mazzoni E. Slug Monitoring and Impacts on the Ground Beetle Community in the Frame of Sustainable Pest Control in Conventional and Conservation Agroecosystems. Insects. 2020; 11(6):380. https://doi.org/10.3390/insects11060380
Chicago/Turabian StyleScaccini, Davide, Michela Panini, Olga Chiesa, Rinaldo Nicoli Aldini, Vincenzo Tabaglio, and Emanuele Mazzoni. 2020. "Slug Monitoring and Impacts on the Ground Beetle Community in the Frame of Sustainable Pest Control in Conventional and Conservation Agroecosystems" Insects 11, no. 6: 380. https://doi.org/10.3390/insects11060380
APA StyleScaccini, D., Panini, M., Chiesa, O., Nicoli Aldini, R., Tabaglio, V., & Mazzoni, E. (2020). Slug Monitoring and Impacts on the Ground Beetle Community in the Frame of Sustainable Pest Control in Conventional and Conservation Agroecosystems. Insects, 11(6), 380. https://doi.org/10.3390/insects11060380