Are Yellow Sticky Cards and Light Traps Effective on Tea Green Leafhoppers and Their Predators in Chinese Tea Plantations?

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
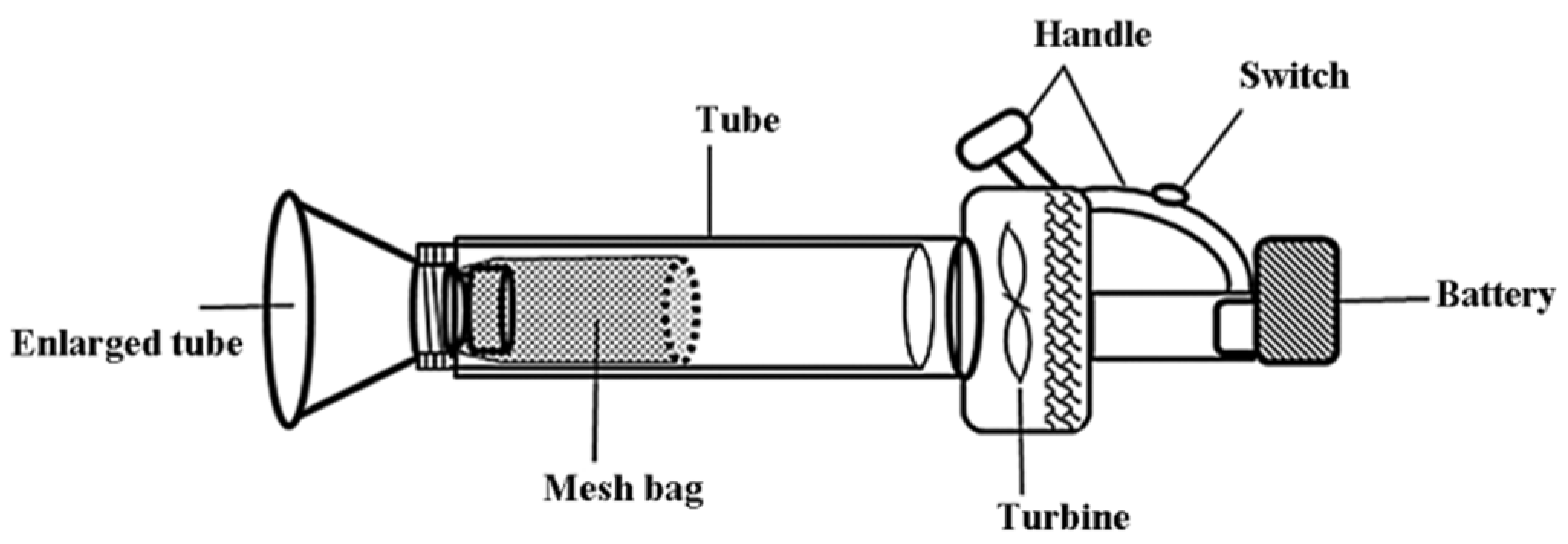
2.2. Open-Field Experiments
2.3. Exclosure Experiment
2.4. Statistical Analyses
2.4.1. Open-Field Experiments
2.4.2. Exclosure Experiment
3. Results
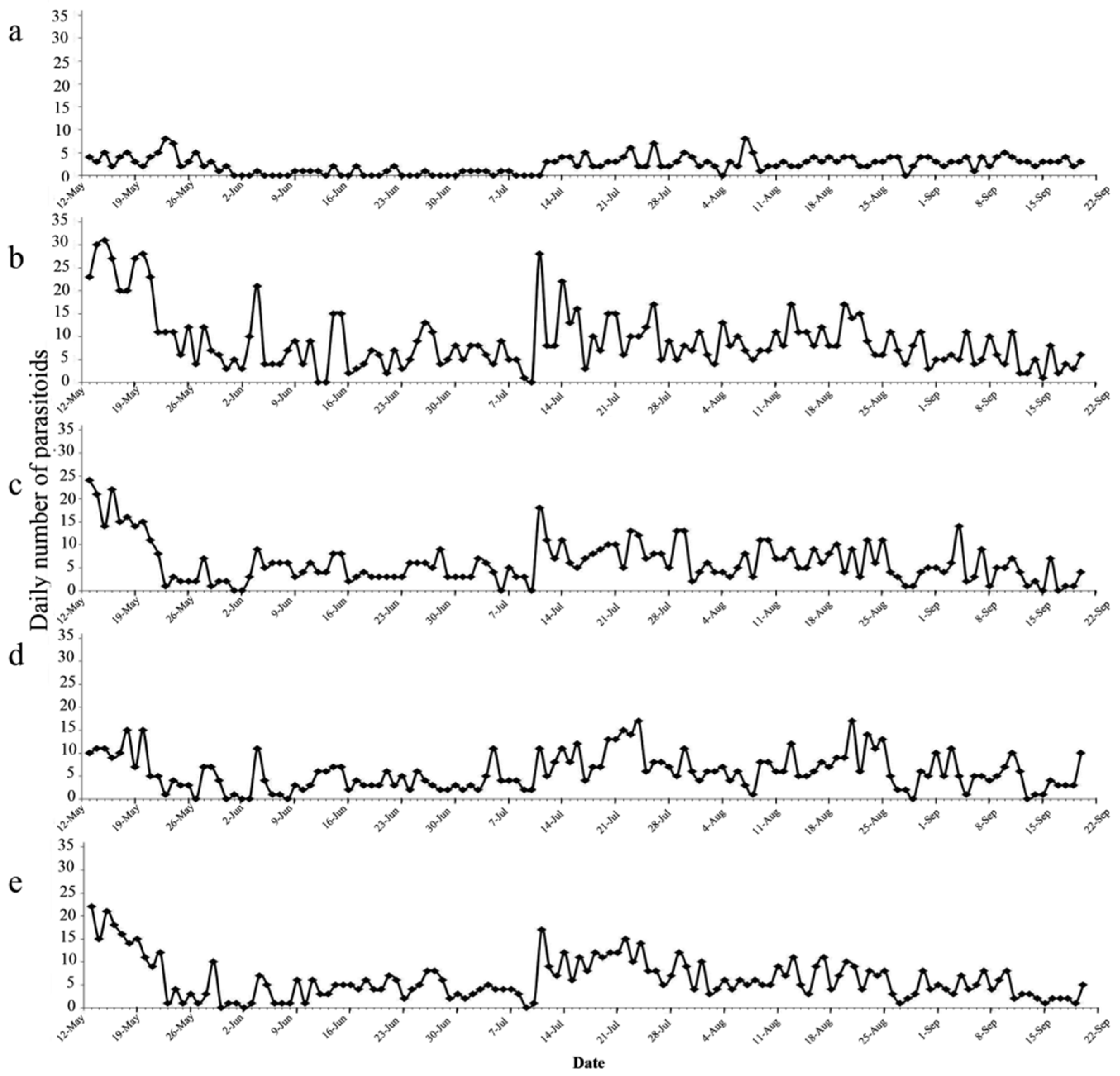
3.1. Sticky Traps Attracted Predators and More E. Onukii Males than Females
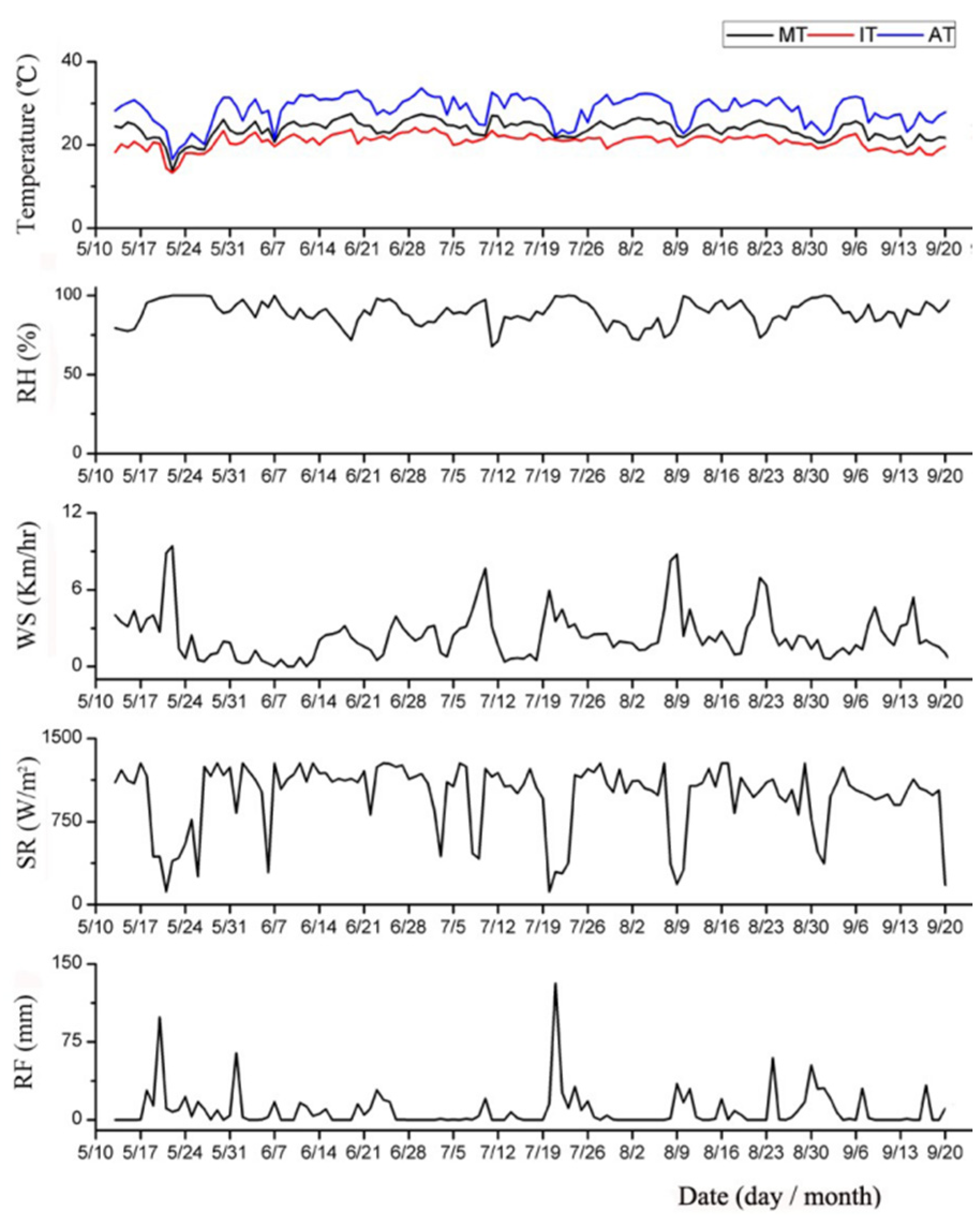
3.2. Weather Factors Influenced the Effectiveness of The Traps
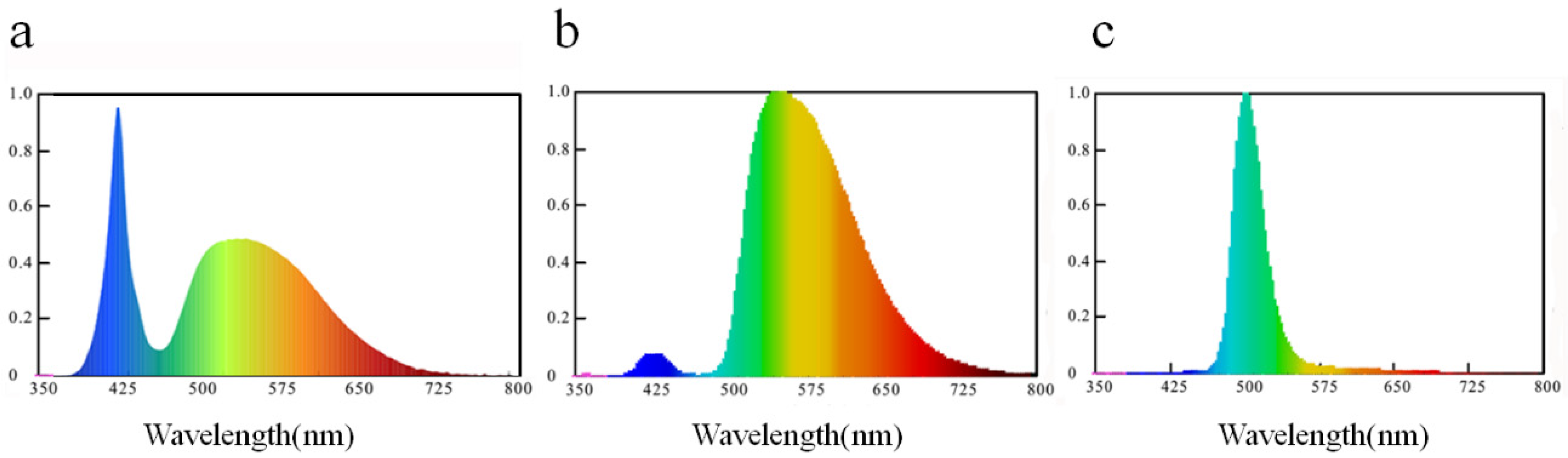
3.3. Light without Trap Worked Better than Sticky Cards to Control Leafhopper
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statements
Acknowledgments
Conflicts of Interest
References
- Wu, M.X.; Han, R.J.; Wang, S.Y.; Jiang, P.; Zhang, Q.L.; Xia, L.S.; Han, B.Y. Promoting effect of intercropping on insect diversity in Chinese plum-tea or medical dogwood-tea or chestnut-tea intercrop plantations. Chin. Bull. Entomol. 2010, 47, 1165–1169. [Google Scholar]
- Marquard, E.; Schmid, B.; Roscher, C.; De Luca, E.; Nadrowski, K.; Weisser, W.W.; Weigelt, A. Changes in the abundance of grassland species in monocultures versus mixtures and their relation to biodiversity effects. PLoS ONE 2013, 8, e75599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mortimer, P.E.; Gui, H.; Xu, J.; Zhang, C.; Barrios, E.; Hyde, K.D. Alder trees enhance crop productivity and soil microbial biomass in tea plantations. Appl. Soil Ecol. 2015, 96, 25–32. [Google Scholar] [CrossRef]
- Chen, Z.M.; Lin, Z. Tea and human health: Biomedical functions of tea active components and current issues. J. Zhejiang Univ. Sci. B 2015, 16, 87–102. [Google Scholar] [CrossRef] [Green Version]
- Hayward, D.G.; Wong, J.W.; Park, H.Y. Determinations for pesticides on Black, Green, Oolong and White teas by gas chromatography triple-quadrupole mass spectrometry. J. Agric. Food Chem. 2015, 63, 8116–8124. [Google Scholar] [CrossRef]
- Qin, D.; Zhang, L.; Xiao, Q.; Dietrich, C.; Matsumura, M. Clarification of the identity of the tea green leafhopper based on morphological comparison between Chinese and Japanese specimens. PLoS ONE 2015, 10, e0139202. [Google Scholar] [CrossRef]
- Zhang, W.; Lin, M.; Zhang, H. Relationship between temperature and development of Emapoasca vitis Göthe (Lepidoptera: Cicadelidae). J. Anhui Agric. Univ. 1997, 24, 332–335. [Google Scholar]
- Tang, Z.Y.; Tang, X.J. Effects and forecast of temperature on occurrence of Empoasca pirisuga Matumura. J. Anhui Agric. Sci. 2010, 38, 3523–3524. [Google Scholar]
- Jin, S.; Chen, Z.M.; Backus, E.A.; Sun, X.L.; Xiao, B. Characterization of EPG waveforms for the tea green leafhopper; Empoasca vitis Göthe (Hemiptera: Cicadellidae); on tea plants and their correlation with stylet activities. J. Insect Physiol. 2012, 58, 1235–1244. [Google Scholar] [CrossRef]
- Nakai, M. Biological control of tortricidae in tea fields in Japan using insect viruses and parasitoids. Virol. Sin. 2009, 24, 323–332. [Google Scholar] [CrossRef]
- Chen, L.L.; You, M.S.; Chen, S.B. Effects of cover crops on spider communities in tea plantations. Biol. Control 2011, 59, 326–335. [Google Scholar] [CrossRef]
- Uesugi, R.; Sato, Y. Counting adults on a yellow sticky trap is useful for monitoring the tea spiny whitefly, Aleurocanthus camelliae (Hemiptera, Aleyrodidae), at the early invasion stage. Jpn. J. Appl. Entomol. Zool. 2013, 57, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Sinu, P.A.; Mandal, P.; Banerjee, D.; Mallick, S.; Talukdar, T.; Pathak, S.K. Moth pests collected in light traps of tea plantations in North East India: Species composition, seasonality and effect of habitat type. Curr. Sci. 2013, 104, 646–651. [Google Scholar]
- Bian, L.; Sun, X.L.; Luo, Z.X.; Zhang, Z.Q.; Chen, Z.M. Design and selection of trap color for capture of the tea leafhopper, Empoasca vitis, by orthogonal optimization. Entomol. Exp. Appl. 2014, 151, 247–258. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, C.; Zhang, J.C.; Zhao, S.Y. Discussion on applicability of the technology of using light to trap in the field of pests and diseases control in tea plantation of China. Key Eng. Mater. 2014, 575, 487–493. [Google Scholar] [CrossRef]
- Yee, W.L. Ambient light intensity and direction determine relative attractiveness of yellow traps to Rhagoletis indifferens (Diptera, Tephritidae). Can. Entomol. 2015, 147, 776–786. [Google Scholar] [CrossRef]
- Szenasi, A.; Marko, V. Flea beetles (Coleoptera, Chrysomelidae; Alticinae) in Bt-(MON810) and near isogenic maize stands, Species composition and activity densities in Hungarian fields. Crop Prot. 2015, 77, 38–44. [Google Scholar] [CrossRef]
- Böll, S.; Herrmann, J. A long-term study on the population dynamics of the grape leafhopper (Empoasca vitis) and antagonistic mymarid species. J. Pest Sci. 2004, 77, 33–42. [Google Scholar] [CrossRef]
- Johansen, N.S.; Vanninen, I.; Pinto, D.M.; Nissinen, A.I.; Shipp, L. In the light of new greenhouse technologies, 2. Direct effects of artificial lighting on arthropods and integrated pest management in greenhouse crops. Ann. Appl. Biol. 2011, 159, 1–27. [Google Scholar] [CrossRef]
- Verovnik, R.; Fiser, Z.; Zaksek, V. How to reduce the impact of artificial lighting on moths, A case study on cultural heritage sites in Slovenia. J. Nat. Conserv. 2015, 28, 105–111. [Google Scholar] [CrossRef]
- Junnila, A.; Kline, D.L.; Müller, G.C. Comparative efficacy of small commercial traps for the capture of adult Phlebotomus papatasi. J. Vector Ecol. 2011, 36, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Stukenberg, N.; Gebauer, K.; Poehling, H.M. Light emitting diode (LED)-based trapping of the greenhouse whitefly (Trialeurodes vaporariorum). J. Appl. Entomol. 2015, 139, 268–279. [Google Scholar] [CrossRef]
- Pawson, S.M.; Watt, M.S.; Brockerhoff, E.G. Using differential responses to light spectra as a monitoring and control tool for Arhopalus ferus (Coleoptera, Cerambycidae) and other exotic wood-boring pests. J. Econ. Entomol. 2009, 102, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Lessio, F.; Tedeschi, R.; Pajoro, M.; Alma, A. Seasonal progression of sex ratio and phytoplasma infection in Scaphoideus titanus Ball (Hemiptera, Cicadellidae). Bull. Entomol. Res. 2009, 99, 377–383. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.Q.; Zeng, Z.H.; Huang, H.S.; Zhou, Y.M.; Vasseur, L.; You, M.S. Identification of Empoasca onukii (Hemiptera, Cicadellidae) and monitoring of its populations in the tea plantations of South China. J. Econ. Entomol. 2015, 108, 1025–1033. [Google Scholar] [CrossRef]
- Altermatt, F.; Baumeyer, A.; Ebert, D. Experimental evidence for male biased flight-to-light behavior in two moth species. Entomol. Exp. Appl. 2009, 130, 259–265. [Google Scholar] [CrossRef]
- Goretti, E.; Coletti, A.; Di Veroli, A.; Di Giulio, A.M.; Gaino, E. Artificial light device for attracting pestiferous chironomids (Diptera), A case study at Lake Trasimeno (Central Italy). Ital. J. Zool. 2011, 78, 336–342. [Google Scholar] [CrossRef]
- Chen, Z.; Xu, R.; Kuang, R.P.; Sun, R. Phototactic behaviour of the parasitoid Encarsia formosa (Hymenoptera, Aphelinidae). Biocontrol Sci. Technol. 2016, 26, 250–262. [Google Scholar] [CrossRef]
- Boeckmann, E.; Hommes, M.; Meyhöefer, R. Yellow traps reloaded, what is the benefit for decision making in practice? J. Pest Sci. 2015, 88, 439–449. [Google Scholar] [CrossRef]
- Rrichards, O.W. Hymenoptera, Introduction and Key to Families; China Science Press: Beijing, China, 1985. (In Chinese) [Google Scholar]
- Feng, Z. Spiders of China in Colour; Hunan Science and Technology Press: Changsha, China, 1990; ISBN 7535704654. (In Chinese) [Google Scholar]
- Liu, S.; Li, Z.; Sui, Y.; Schaefer, D.A.; Alele, P.O.; Chen, J.; Yang, X. Spider foraging strategies dominate pest suppression in organic tea plantations. Biocontrol 2015, 60, 839–847. [Google Scholar] [CrossRef]
- Chen, Y.F. Review on the investigation and protection measurement of spiders in Chinese tea gardens. Acta Arachnol. Sin. 2004, 13, 125–128. [Google Scholar]
- Heiling, A.M. Why do nocturnal orb-web spiders (Araneidae) search for light? Behav. Ecol. Sociobiol. 1999, 46, 43–49. [Google Scholar] [CrossRef]
- Lizotte, R.S.; Rovner, J.S. Nocturnal capture of fireflies by lycosid spiders, visual versus vibratory stimuli. Anim. Behav. 1988, 36, 1809–1815. [Google Scholar] [CrossRef]
- Virant–Doberlet, M.; King, R.A.; Polajnar, J.; Symondson, W.O.C. Molecular diagnostics reveal spiders that exploit prey vibrational signals used in sexual communication. Mol. Ecol. 2011, 20, 2204–2216. [Google Scholar] [CrossRef] [PubMed]
- Han, B.Y.; Lin, J.L.; Zhou, X.G.; Zhang, J.M. Investigation on morphology of eggs of tea green leafhoppers and morphology and parasitic percentages of mymarids parasitizing the leafhopper eggs. J. Anhui Agric. Univ. 2009, 36, 13–17. [Google Scholar]
- Abdus, S.K. Biology of Anagrus optabilis (perkins) (Hymenoptera, Mymaridae), an egg parasitoid of Delphacid planthoppers. Esakia 1984, 22, 129–144. [Google Scholar]
- Charnov, E.L.; Skinner, S.W. Complementary approaches to the understanding of parasitoid oviposition decisions. Environ. Entomol. 1985, 14, 383–391. [Google Scholar] [CrossRef]
- Takagi, M. The reproductive strategy of the gregarious parasitoid, Pteromalus puparum (Hymenoptera, Pteromalidae). Oecologia 1987, 71, 321–324. [Google Scholar] [CrossRef]
- Berndt, L.A.; Wratten, S.D.; Hassan, P.G. Effects of buckwheat flowers on leafroller (Lepidoptera, Tortricidae) parasitoids in a New Zealand vineyard. Agric. For. Entomol. 2002, 4, 39–45. [Google Scholar] [CrossRef]
- Shi, L.Q.; Vasseur, L.; Huang, H.S.; Zeng, Z.H.; Hu, G.P.; Liu, X.; You, M.S. Adult tea green leafhoppers, Empoasca onukii (Matsuda), change behaviors under varying light conditions. PLoS ONE 2017, 12, e0168439. [Google Scholar] [CrossRef]
- Pellegrino, A.C.; Nardi, C.; Bezner-Kerr, W.; Guglielmo, C.G.; Bento, J.M.S.; Mcneil, J.N. Weather forecasting by insects, modified sexual behaviour in response to atmospheric pressure changes. PLoS ONE 2013, 8, e75004. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.V.R.; Rashmi, T.; Verghese, A. Foraging activity of Indian honey bee, Apis cerana in relation to ambient climate variables under tropical conditions. J. Environ. Biol. 2015, 36, 577–581. [Google Scholar]
- Sanderson, R.A.; Goffe, L.A.; Leifert, C. Time-series models to quantify short-term effects of meteorological conditions on bumblebee forager activity in agricultural landscapes. Agric. For. Entomol. 2015, 17, 270–276. [Google Scholar] [CrossRef]
- Yase, J.; Yamanaka, M.; Fujii, H.; Kosaka, S. Control of tobacco budworm., Helicoverpa armigera (Hubner), beet armyworm Spodoptera exigua (Hubner), common cutworm., Spodoptera litura (Fabricius), feeding on carnation, roses and chrysanthemum by overnight illumination with yellow fluorescent lamps. Kinki Chugoku Agric. Res. 1997, 93, 10–14. [Google Scholar]
- Yase, J.; Nagaoka, O.; Futai, K.; Izumida, T.; Kosaka, S. Control of cabbage webworm, Hellula undalis Fabricius (Lepidoptera, Pyralidae) using yellow fluorescent lamps. Appl. Entomol. Zool. 2004, 46, 29–37. [Google Scholar]
- Briscoe, A.D.; Chittka, L. The evolution of color vision in insects. Annu. Rev. Entomol. 2001, 46, 471–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bernard, G.D.; Remington, C.L. Color vision in Lycaena butterflies, spectral tuning of receptor arrays in relation to behavioral ecology. Proc. Natl. Acad. Sci. USA 1991, 88, 2783–2787. [Google Scholar] [CrossRef] [Green Version]
- Nishio, J.N. Why are higher plants green? Evolution of the higher plant photosynthetic pigment complement. Plant Cell Environ. 2000, 236, 539–548. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Technique | Spider | Parasitoid | Female Leafhopper | Male Leafhopper | Total Leafhopper | Female Ratio of Leafhopper |
|---|---|---|---|---|---|---|
| Suction Machine | 306.2 ± 13.8 | 65.4 ± 11.2 | 1686.4 ± 69.8 | 604.1 ± 25.3 | 2290.4 ± 74.5 | 0.713 ± 0.014 |
| White Light | 145.4 ± 14.3 a | 160.7 ± 25.2 b | 612.4 ± 78.2 a | 2003.7 ± 231.1 a | 2616.3 ± 182.1 a | 0.234 ± 0.035 a |
| Green Light | 173.6 ± 26.9 a | 154.4 ± 17.1 b | 530.7 ± 68.2 a | 1825.4 ± 220.2 b | 2354.5 ± 253.3 b | 0.228 ± 0.028 a |
| Yellow Light | 151.1 ± 11.7 a | 160.4 ± 30.3 b | 515.6 ± 66.4 a | 1785.3 ± 152.4 b | 2301.3 ± 213.7 b | 0.229 ± 0.013 a |
| Yellow Card | 124.4 ± 18.2 a | 237.1 ± 18.3 a | 465.2 ± 60.2 a | 1806.5 ± 204.3 b | 2272.4 ± 264.3 b | 0.214 ± 0.016 a |
| Treatment | Adult E. Onukii (Y1) | Parasitoid (Y2) | Spider (Y3) |
|---|---|---|---|
| Yellow sticky card | Ln(Y1 + 1) = 3.086–0.001X6, r = 0.179, F = 4.277, p = 0.041 | - | Ln(Y3 + 1) = 0.455 + 0.040X3, r = 0.222, F = 6.703, p = 0.011 |
| White light trap | - | - | Ln(Y3 + 1) = −0.660 + 0.115X1, r = 0.340, F = 16.875, p < 0.001 |
| Green light trap | Ln(Y1 + 1) = 2.598 + 0.009X4, r = 0.221, F = 6.649, p = 0.011 | Ln(Y2 + 1) = 0.123 + 0.048X3 + 0.010X4 + 0.067X7, r = 0.310, F = 4.496, p = 0.005 | Ln(Y3 + 1) = 0.135 + 0.083X1, r = 0.249, F = 8.520, p = 0.004 |
| Yellow light trap | Ln(Y1 + 1) = 2.595 + 0.008X4, r = 0.180, F = 4.298, p = 0.040 | Ln(Y2 + 1) = −0.545 + 0.085X2 + 0.007X4 + 0.097X7, r = 0.364, F = 6.486, p < 0.001 | Ln(Y3 + 1) = 0.013 + 0.084X1, r = 0.255, F = 8.943, p = 0.003 |
| Suction machine (population dynamics) | Ln(Y1 + 1) = 4.295 − 0.061X3, r = 0.242, F = 8.029, p = 0.005 | Ln(Y2 + 1) = 4.151 − 0.149X1, r = 0.408, F = 25.722, p < 0.001 | Ln(Y3 + 1) = 3.486 − 0.044X3 + 0.062X7, r = 0.361, F = 9.568, p < 0.001 |
| Object | White Light | Green Light | Yellow Light | Yellow Sticky Card | Control |
|---|---|---|---|---|---|
| Total E. onukii individuals | 11.68 ± 2.22 b | 12.61 ± 2.02 b | 18.75 ± 2.45 a | 17.40 ± 2.35 a | 17.04 ± 2.44 a |
| E. onukii female | 10.28 ± 1.97 b | 10.72 ± 1.77 b | 15.43 ± 2.09 a | 13.73 ± 1.70 ab | 14.3 ± 2.19 a |
| E. onukii male | 1.41 ± 0.31 b | 1.89 ± 0.35 b | 3.32 ±0.48 a | 3.68 ± 0.70 a | 2.7 ± 0.31 a |
| Spider | 3.21 ± 0.34 a | 2.95 ± 0.22 bc | 3.05 ± 0.32 ab | 2.85 ± 0.23 c | 3.20 ± 0.23 a |
| Objective | Source of Variation | Df | F-Value | p-Value |
|---|---|---|---|---|
| E. onukii (females + males) | Within subject | |||
| Week | 1, 3.917 | 7.419 | <0.001 | |
| Week × treatment | 4, 15.668 | 1.614 | 0.046 | |
| Between subject | ||||
| Treatment | 4, 20 | 0.361 | 0.834 | |
| Female E. onukii | Within subject | |||
| Week | 1, 4.100 | 8.364 | <0.001 | |
| Week × treatment | 4, 23.419 | 1.794 | 0.035 | |
| Between subject | ||||
| Treatment | 4, 20 | 0.333 | 0.852 | |
| Male E. onukii | Within subject | |||
| Week | 1, 5.855 | 6.052 | <0.001 | |
| Week × treatment | 4, 16.401 | 1.356 | 0.147 | |
| Between subject | ||||
| Treatment | 4, 20 | 0.310 | 0.868 | |
| Spiders | Within subject | |||
| Week | 1, 15 | 3.585 | <0.001 | |
| Week × treatment | 4, 60 | 0.976 | 0.530 | |
| Between subject | ||||
| Treatment | 4, 20 | 0.762 | 0.562 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, L.; He, H.; Yang, G.; Huang, H.; Vasseur, L.; You, M. Are Yellow Sticky Cards and Light Traps Effective on Tea Green Leafhoppers and Their Predators in Chinese Tea Plantations? Insects 2021, 12, 14. https://doi.org/10.3390/insects12010014
Shi L, He H, Yang G, Huang H, Vasseur L, You M. Are Yellow Sticky Cards and Light Traps Effective on Tea Green Leafhoppers and Their Predators in Chinese Tea Plantations? Insects. 2021; 12(1):14. https://doi.org/10.3390/insects12010014
Chicago/Turabian StyleShi, Longqing, Haifang He, Guang Yang, Huoshui Huang, Liette Vasseur, and Minsheng You. 2021. "Are Yellow Sticky Cards and Light Traps Effective on Tea Green Leafhoppers and Their Predators in Chinese Tea Plantations?" Insects 12, no. 1: 14. https://doi.org/10.3390/insects12010014
APA StyleShi, L., He, H., Yang, G., Huang, H., Vasseur, L., & You, M. (2021). Are Yellow Sticky Cards and Light Traps Effective on Tea Green Leafhoppers and Their Predators in Chinese Tea Plantations? Insects, 12(1), 14. https://doi.org/10.3390/insects12010014