Is Orius sauteri Poppius a Promising Biological Control Agent for Walnut Aphids? An Assessment from the Laboratory to Field

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Collection
2.2. Laboratory Experiments
2.2.1. The Functional Response of Predatory O. sauteri Bugs to Increasing Density of P. juglandis and C. juglandicola Aphids
2.2.2. Predator Preference for Prey Species
2.2.3. Biological Control Efficacy of Predatory O. sauteri against a Single Population of P. juglandis or C. juglandicola
2.3. Field Experiments
2.3.1. Biological Control Efficacy of Predatory O. sauteri against a Single Population of P. juglandis or C. juglandicola
2.3.2. Biological Control Efficacy of Predatory O. sauteri against a Mixed Population of P. juglandis and C. juglandicola
2.4. Data Analysis
2.4.1. Predator Preference for Prey Species
2.4.2. Biological Control Efficacy of Predatory O. sauteri against P. juglandis and C. juglandicola
3. Results
3.1. The Functional Response of Predatory O. sauteri Bugs to Increasing Density of P. juglandis and C. juglandicola
3.2. Predator Preference for Prey Species
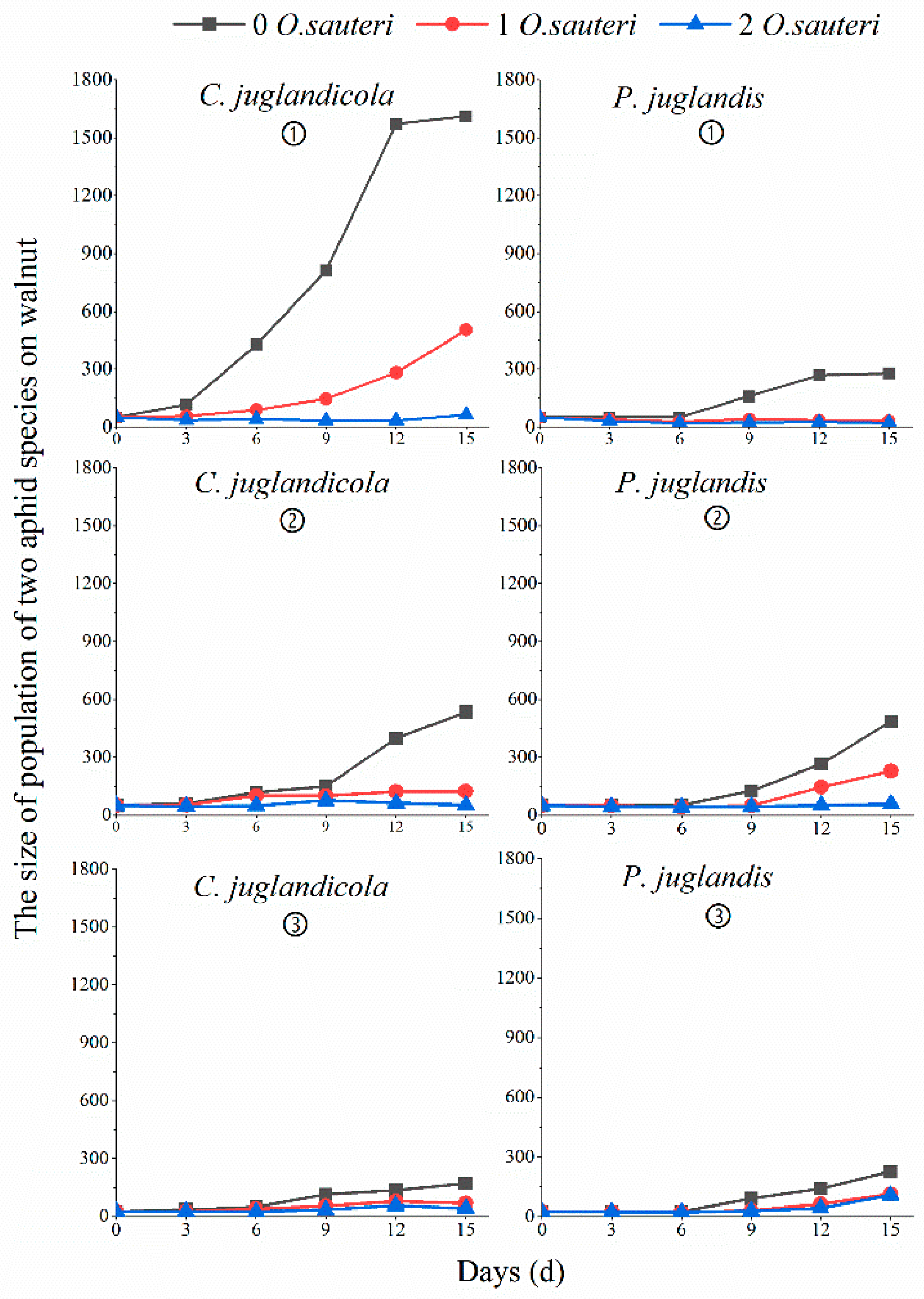
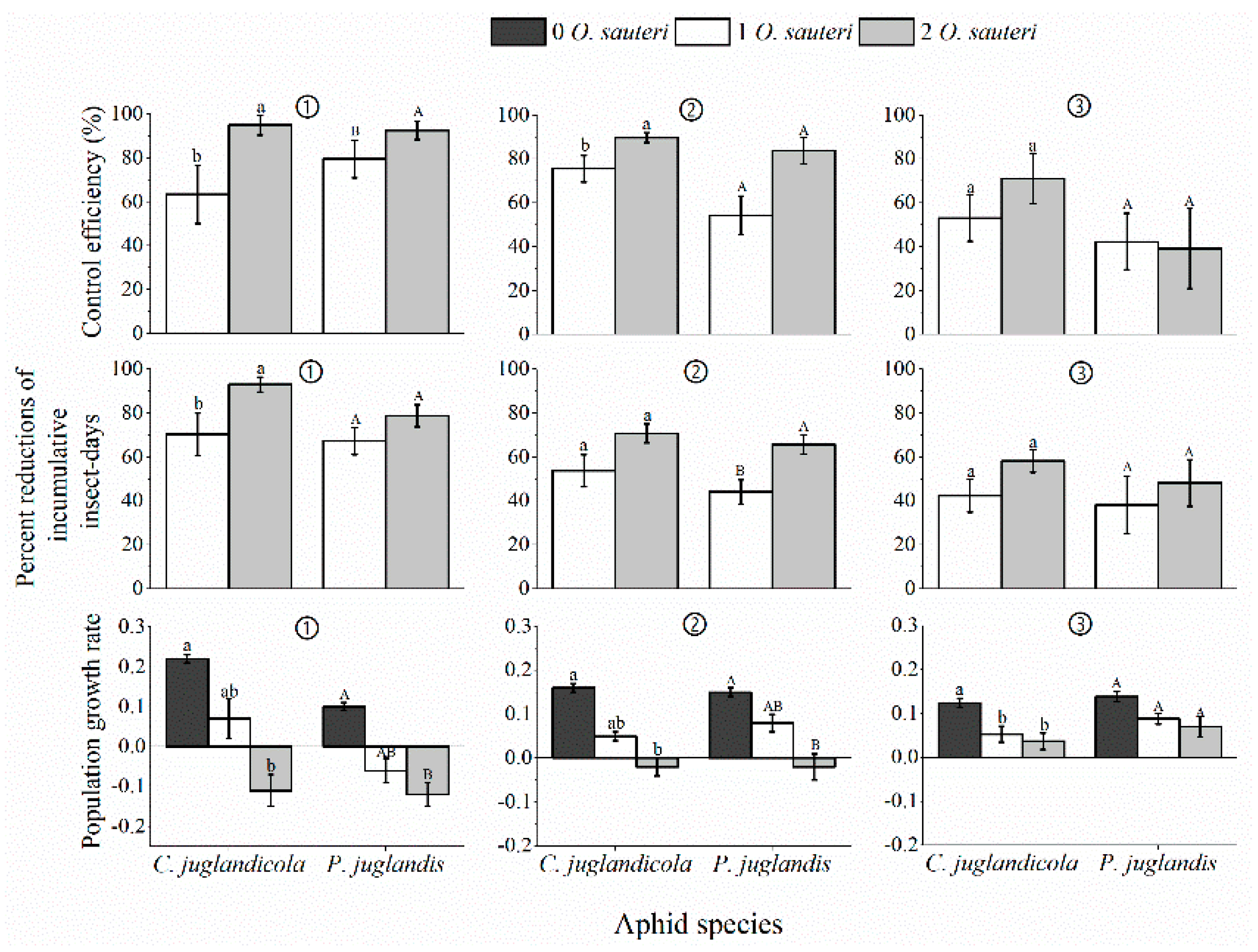
3.3. Biological Control Efficacy of Predatory O. sauteri against a Single Population of P. juglandis or C. juglandicola in the Laboratory and Field
3.4. Biological Control Efficacy of Predatory O. sauteri against a Mixed Population of P. juglandis and C. juglandicola Aphids in the Open Field
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Orre-Gordon, G.U.S.; Wratten, S.D.; Tompkins, J.M.; Jonsson, M.; Jacometti, M.A. Provision of floral resources for biological control: Restoring an important ecosystem service. Funct. Ecosyst. Communities 2007, 1, 86–94. [Google Scholar]
- O’Neil, R.J.; Giles, K.L.; Obrycki, J.J.; Mahr, D.L.; Legaspi, J.C.; Katovich, K. Evaluation of the Quality of Four Commercially Available Natural Enemies. Biol. Control 1998, 11, 1–8. [Google Scholar] [CrossRef]
- van Lenteren, J.C. Ecosystem services to biological control of pests: Why are they ignored? Proc. Neth. Entomol. Soc. Meet. 2006, 17, 103–111. [Google Scholar]
- Costanza, R.; d’Arge, R.; de Groot, R.; Farber, S.; Grassot, M.; Hannon, B.; Limburg, K.; Naeem, S.; O’Neill, R.V.; Paruelo, J.; et al. The value of the world’s ecosystem services and natural capital. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- van Lenteren, J.C. IOBC Internet Book of Biological Control. IOBC-Global 2012, 6, 182. [Google Scholar]
- KrzyŻAnowski, R. Population dynamics and spatial distribution of Panaphis juglandis (Goeze, 1778) (Hemiptera: Aphididae) on common walnut (Juglans regia L.). Folia Pomer. Univ. Technol. Stetin. Agric. Aliment. Pisc. Zootech. 2018, 341, 29–40. [Google Scholar] [CrossRef]
- Aqaverdi, N.I.; Inqilab, N.G. Some bioecological pecularities of panaphis juglandis (Goeze, 1778) and chromaphis juglandicola (Kaltenbach, 1843) (Hemiptera, Aphididae) the pests of Persian walnut (Juglans regia L.) in Azerbaijan. J. Entomol. Zool. Stud. 2018, 6, 800–803. [Google Scholar]
- Steinmann, K.P.; Zhang, M.; Grant, J.A.; Pickel, C.; Goodhue, R.E. Pheromone-based pest management can be cost-effective for walnut growers. Calif. Agric. 2008, 62, 105–110. [Google Scholar] [CrossRef] [Green Version]
- Akkopru, E.P.; Han, R.A.; Okut, H.; Chi, H. Demographic assessment of plant cultivar resistance to insect pests: A case study of the dusky-veined walnut aphid (Hemiptera: Callaphididae) on five walnut cultivars. J. Econ. Entomol. 2015, 108, 378–387. [Google Scholar] [CrossRef]
- Wani, S.; Ahmad, S. Biology and population dynamics of new colour morph of Chromaphis juglandicola Kalt. J. Entomol. Zool. Stud. 2015, 3, 28–31. [Google Scholar]
- Jaskiewicz, B.; Kmiec, K. The occurrence of Panaphis juglandis (Goetze) and Chromaphis juglandicola (Kalt.) on walnut under the urban conditions of Lublin. Acta Sci. Pol. Hortorum Cultus 2007, 6, 15–26. [Google Scholar]
- van den Bosch, R.; Frazer, B.D.; Davis, C.S.; Messenger, P.S.; Hom, R. Trioxys pallidus an effective walnut aphid parasite from Iran. Calif. Agric. 1970, 24, 8–10. [Google Scholar]
- Singh, R.; Singh, G. Chapter 3: Aphids and their biocontrol. In Ecofriendly Pest Management for Food and Security; Academic Press: Cambridge, MA, USA, 2016; pp. 63–108. [Google Scholar]
- Letourneau, D.K.; Altierl, M.A. Abundance patterns of a predator, Orius tristicolor (Hemiptera: Anthocoridae), and its prey, Frankliniella occidentalis (Tysanoptera Thripidae) habitat attraction in polycultures versus monocultures. Environ. Entomol. 1983, 12, 1464–1469. [Google Scholar] [CrossRef]
- Isenhour, D.J.; Yeargan, K.V. Predation by orius insidiosus on the soybean thrips, sericothrips variabilis: Effect of prey stage and density. Environ. Entomol. 1981, 10, 496–500. [Google Scholar] [CrossRef]
- Isenhour, D.J.; Yeargan, K.V. Interactive behavior of Orius insidiosus [Hem.: Anthocoridae] and Sericothrips variabilis [Thys.: Thripidae]: Predator searching strategies and prey escape tactics. Entomophaga 1981, 26, 213–220. [Google Scholar] [CrossRef]
- Wang, Y.P.; Wu, H.; Bu, W.J.; Xu, H.C. Geographic distribution of the genus Orius Wolff (Heteroptera: Anthocoridae). J. Zhejiang For. Coll. 2003, 20, 389–393. [Google Scholar]
- Nagai, K.; Yano, E. Predation by Orius sauteri (Poppius) (Heteroptera: Anthocoridae) on Thrips palmi Karny (Thysanoptera: Thripidae): Functional response and selective predation. Appl. Entomol. Zool. 2000, 35, 565–574. [Google Scholar] [CrossRef] [Green Version]
- Paik, C.H.; Lee, G.H.; Hwang, C.Y.; Kim, S.J. Predatory Response of the Pirate Bug, Orius sauteri Poppius (Heteroptera: Anthocoridae) on Frankliniella occidentalis, Aphid gossypii and Tetranychus urticae. Korean J. Appl. Entomol. 2010, 49, 401–407. [Google Scholar] [CrossRef] [Green Version]
- Ogino, T.; Yamaguchi, T.; Uehara, T.; Kainoh, Y.; Shimoda, M. Analysis of the activity rhythm of the predatory bug Orius sauteri (Poppius) (Heteroptera: Anthocoridae) for optimizing its selective light attraction. Appl. Entomol. Zool. 2019, 55, 115–120. [Google Scholar] [CrossRef]
- Holling, C.S. Some characteristics of simple types of predation and parasitism. Can. Entomol. 1959, 7, 385–398. [Google Scholar] [CrossRef]
- Jeschke, J.M.; Tollrian, K.R. Predator functional responses: Discriminaing between handling and digesting pery. Ecol. Monogr. 2002, 72, 95–112. [Google Scholar] [CrossRef]
- Chakravarti, I.M.; Laha, G.A.; Roy, J. Handbook of Methods of Applied Statistics; John Wiley and Sons: Hoboken, NJ, USA, 1967; Volume 1, pp. 392–394. [Google Scholar]
- Mei, Z.J.; Lu, Y.j.; Li, J.Y.; Wang, Z.Y.; Zheng, M.; Wang, Y. Predation functional response and preference of several the third instar s, torage pests by Xylocoris flavipes. J. Chin. Cereals Oils Assoc. 2019, 34, 104–109. [Google Scholar]
- Jabra-Rizk, M.A.; Shirtliff, M.; James, C.; Meiller, T. Effect of farnesol on Candida dubliniensis biofilm formation and fluconazole resistance. FEMS Yeast Res. 2006, 6, 1063–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruppel, R.F. Cumulative insect-days as an index of crop protectionl. J. Econ. Entomol 1983, 76, 375–377. [Google Scholar] [CrossRef]
- Chen, X.D.; Stark, J.D. Individual- and population-level toxicity of the insecticide, spirotetramat and the agricultural adjuvant, Destiny to the Cladoceran, Ceriodaphnia dubia. Ecotoxicology 2010, 19, 1124–1129. [Google Scholar] [CrossRef] [PubMed]
- Han, L.L.; Dong, T.Y.; Zhao, K.J.; Zhu, M.; Sun, W.P.; Xu, Z.X.; Shi, L. Predation of Aphis glycines by Orius sauteri nymphs. Chin. J. Biol. Control 2015, 31, 322–326. [Google Scholar]
- Wu, Y.Q.; Zhao, M.Q.; Yang, S.F.; Duan, Y.; Jiang, Y.L. Predations of Orius sauteri (Hemiptera: Anthocoridae) on four insect pests. Chin. J. Biol. Control 2010, 26, 13–17. [Google Scholar]
- Yerim, L.; Sora, K.; Seunghwan, L. A first record of three aphid pests (Aphididae: Calaphidinae) on walnut in Korea. J. Asia-Pac. Biodivers. 2018, 11, 531–537. [Google Scholar]
- Abbasi, Z.; Sultana, R.; Wagan, M.S. Impact of abiotic factors on population fluctuation of aphid (Hemiptera: Aphididae) on the different wheat varieties from Larkana district. J. Entomol. Zool. Stud. 2019, 7, 608–611. [Google Scholar]
- Karczmarz, K. Numerical strength dynamics of Chromaphis juglandicola (Kalt. 1843) on common walnut (Juglans regia L.) in Lublin town plantings. Acta Sci. Pol. Hortorum Cultus 2010, 9, 121–132. [Google Scholar]
- Romero, G.Q.; Gonçalves-Souza, T.; Kratina, P.; Marion, N.A.C.; Petry, W.K.; Sobral-Souza, T.; Roslin, T. Global predation pressure redistribution under future climate change. Nat. Clim. Chang. 2018, 8, 1087–1091. [Google Scholar] [CrossRef]
- Lin, Q.C.; Chen, H.; Yin, Y.Y.; Zhang, S.C.; Yu, Y.; Zhuang, Q.Y.; Zheng, L.; Zhai, Y.F. Effects of temperature on the development and predation of Aphidoletes aphidimyza (Rondani) larvae. Chin. Appl. Entomol. 2019, 56, 79–84. [Google Scholar]
- Bretagnolle, V.; Gallis, H. Chapter 15: Predator–prey interactions and climate change. In Effects of Climate Change on Birds; Oxford University Press: Oxford, UK, 2019; pp. 119–220. [Google Scholar]
- Barton, B.T. Reduced wind strengthens top-down control of an insect herbivore. Ecology 2014, 95, 2375–2381. [Google Scholar] [CrossRef] [Green Version]
- Feng, N.; Fan, F.; Tao, B.; Yang, X.F.; Wei, G.S. Effect of spectral sensitivity response on the phototaxis of Orius sauteri (Poppius). Acta Ecol. Sin. 2015, 35, 4811–4815. [Google Scholar]
- Lecato, G.L. Predation by Xylocoris flavipes [Hem.: Anthocoridae]: Influence of stage, species and density of prey and of starvation and density of predator. Entomophage 1976, 21, 217–221. [Google Scholar] [CrossRef]
- Fantinou, A.A.; Perdikis, D.C.; Maselou, D.A.; Lambropoulos, P.D. Prey killing without consumption: Does Macrolophus pygmaeus show adaptive foraging behaviour? Biol. Control 2008, 47, 187–193. [Google Scholar] [CrossRef]
- Maselou, D.A.; Perdikis, D.C.; Sabelis, M.W.; Fantinou, A.A. Use of plant resources by an omnivorous predator and the consequences for effective predation. Biol. Control 2014, 79, 92–100. [Google Scholar] [CrossRef]
- Wani, S.A.; Ahmad, S.T. Competition and niche-partitioning in two species of walnut aphids. Int. J. Sci. Res. Rev. 2014, 3, 120–125. [Google Scholar]



{kind=link}
{kind=link}
| Prey | Prey Density (Aphids/Leaf Disc) | |||||
|---|---|---|---|---|---|---|
| 5 | 10 | 15 | 20 | 25 | 35 | |
| Chromaphis juglandicola | 4.2 ± 0.7 bc | 4.8 ± 1.3 b | 6.2 ± 1.1 ab | 9.0 ± 1.1 ab | 11.5 ± 0.7 ab | 11.7 ± 2.2 a |
| Panaphis juglandis | 3.8 ± 0.3 c | 4.0 ± 0.3 c | 6.5 ± 1.3 abc | 8.5 ± 1.3 abc | 8.8 ± 0.5 ab | 9.2 ± 1.6 abc |
| P. juglandis | C. juglandicola | Cain Index | ||
|---|---|---|---|---|
| Density | Predation (Np1) | Density | Predation (Np2) | |
| 15 | 6.87 ± 0.47 | 15 | 4.25 ± 0.51 | 1.82 ± 0.22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, T.; Zhang, P.; Ma, C.; Yasir Ali, M.; Gao, G.; Lu, Z.; Zalucki, M.P. Is Orius sauteri Poppius a Promising Biological Control Agent for Walnut Aphids? An Assessment from the Laboratory to Field. Insects 2021, 12, 25. https://doi.org/10.3390/insects12010025
Wang T, Zhang P, Ma C, Yasir Ali M, Gao G, Lu Z, Zalucki MP. Is Orius sauteri Poppius a Promising Biological Control Agent for Walnut Aphids? An Assessment from the Laboratory to Field. Insects. 2021; 12(1):25. https://doi.org/10.3390/insects12010025
Chicago/Turabian StyleWang, Ting, Ping Zhang, Chenyang Ma, Muhammad Yasir Ali, Guizhen Gao, Zhaozhi Lu, and Myron P. Zalucki. 2021. "Is Orius sauteri Poppius a Promising Biological Control Agent for Walnut Aphids? An Assessment from the Laboratory to Field" Insects 12, no. 1: 25. https://doi.org/10.3390/insects12010025
APA StyleWang, T., Zhang, P., Ma, C., Yasir Ali, M., Gao, G., Lu, Z., & Zalucki, M. P. (2021). Is Orius sauteri Poppius a Promising Biological Control Agent for Walnut Aphids? An Assessment from the Laboratory to Field. Insects, 12(1), 25. https://doi.org/10.3390/insects12010025