The Relationship between Genus/Species Richness and Morphological Diversity among Subfamilies of Jewel Beetles




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
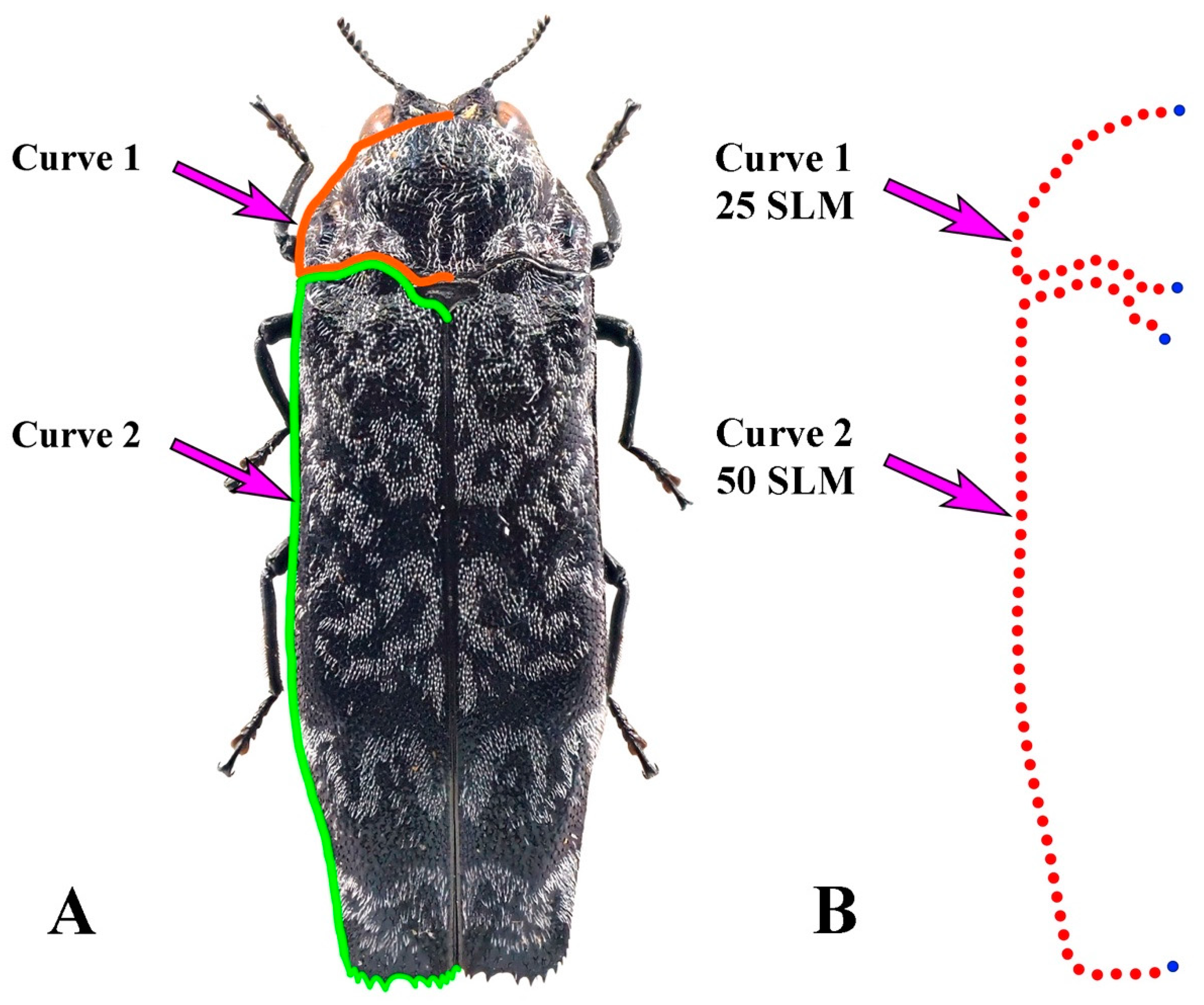
2.1. Taxa Examined
2.2. Genus/Species Richness
2.3. Morphological Diversity
3. Results
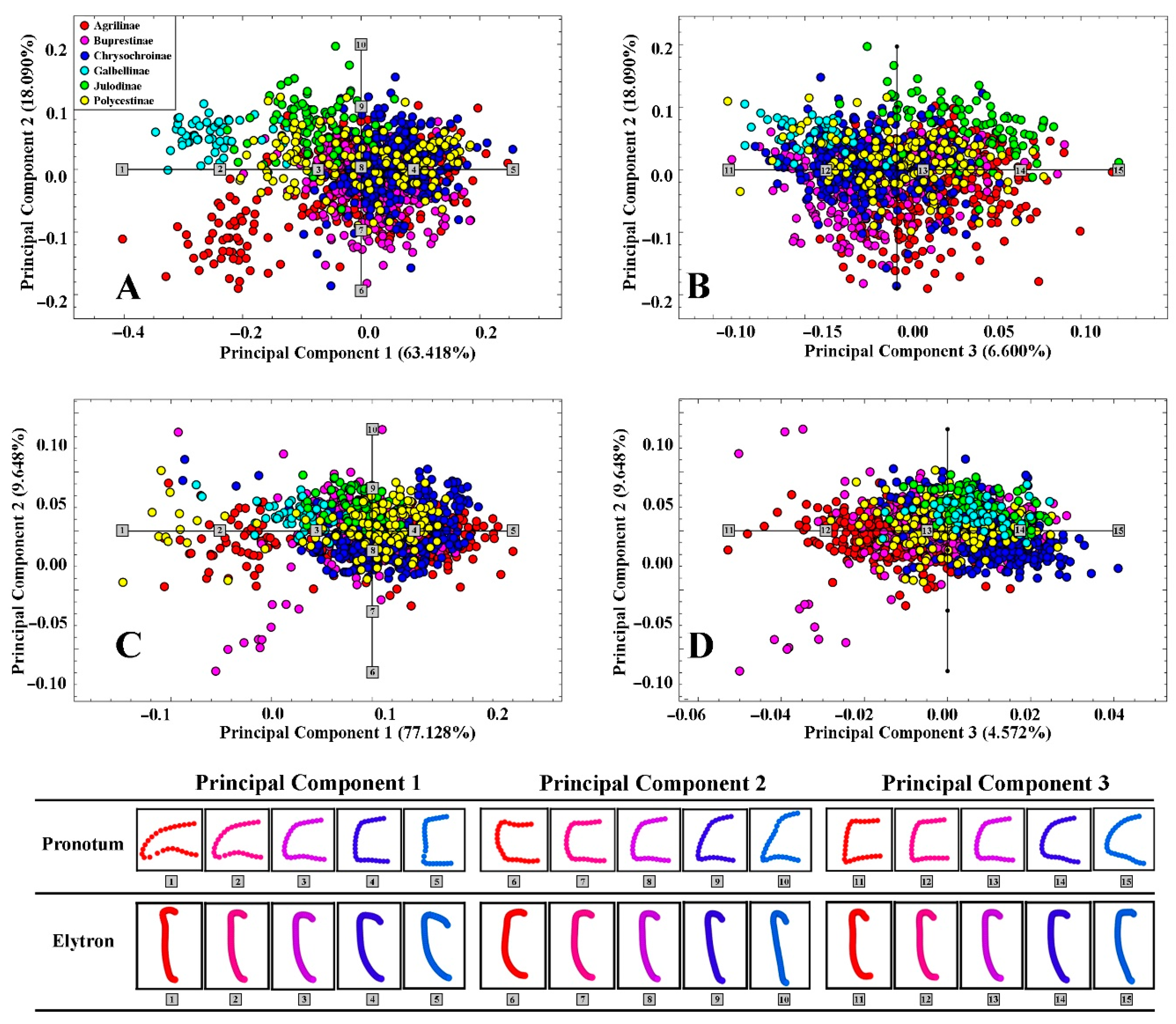
3.1. Morphological Variation of the Pronotum and Elytron
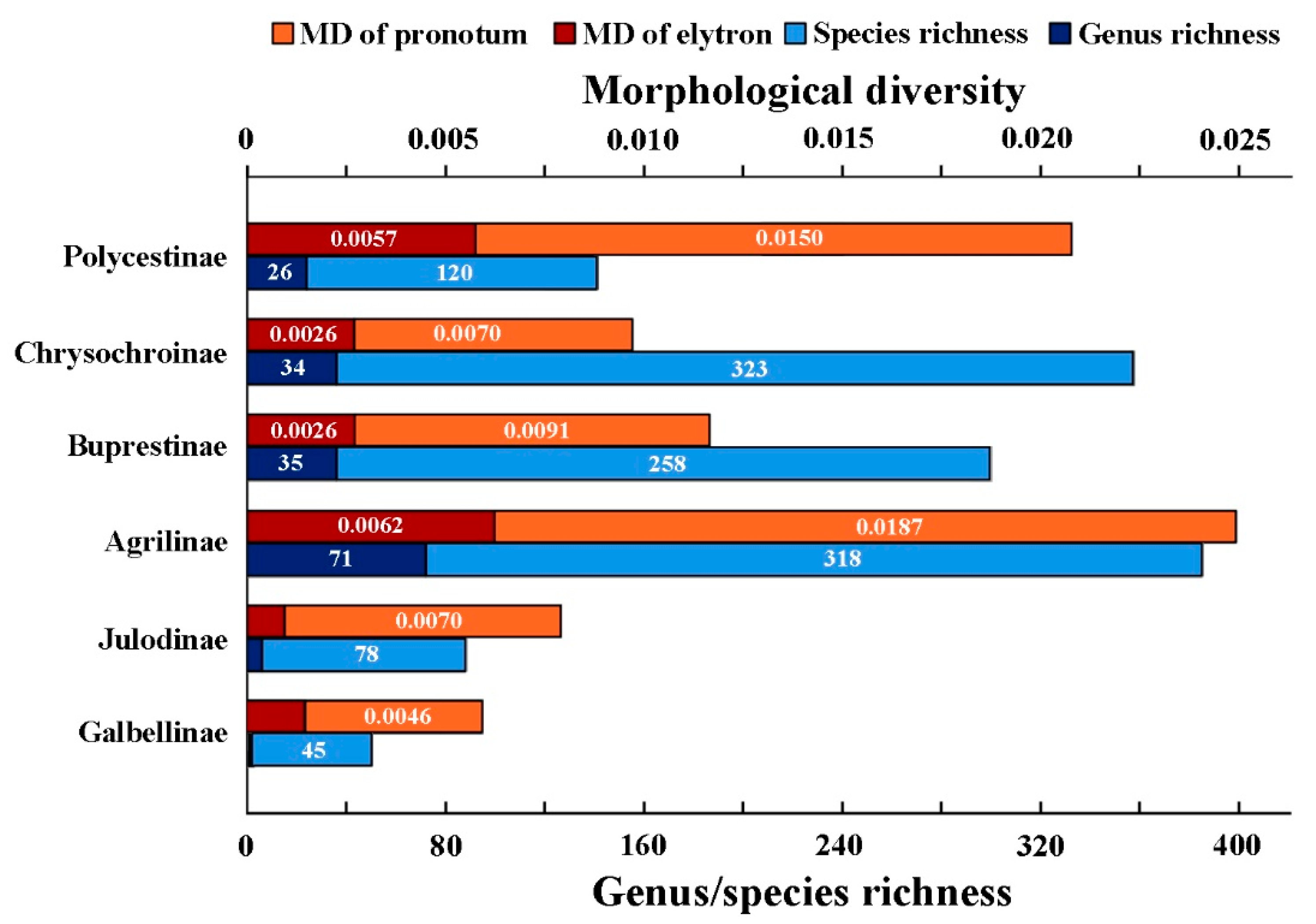
3.2. Morphological Diversity among Groups
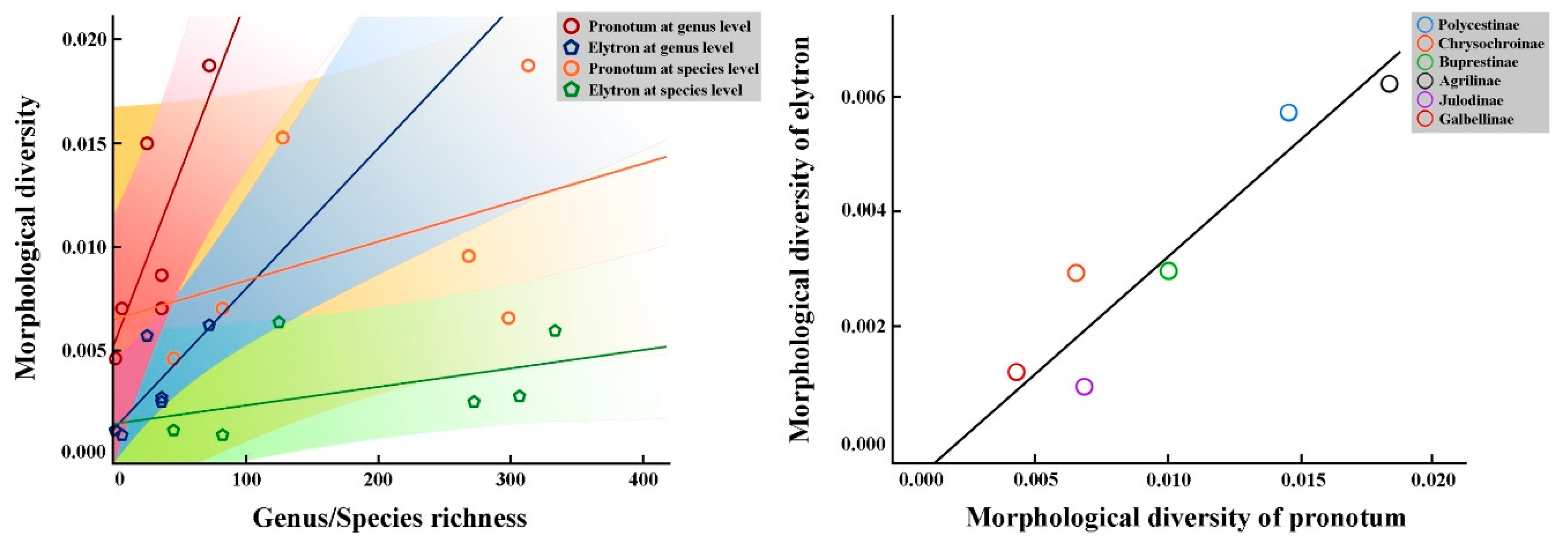
3.3. Correlation Analysis of Test Subfamilies Under Buprestidae
4. Discussion
4.1. Morphological Diversity and Species/Genus Richness of Buprestidae
4.2. Geometric Morphometrics in the Biodiversity Measurement of Higher Taxa
4.3. Biodiversity Measurement Based on Cosmopolitan Dataset
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Williams, P.H.; Humphries, C.J. Comparing Character Diversity Among Biotas; Blackwell Science: Oxford, UK, 1996; pp. 54–76. [Google Scholar]
- Harper, J.L.; Hawksworth, D. Biodiversity: Measurement and estimation. Philos. Trans. R. Soc. Lon B. Biol. Sci. 1994, 345, 5–12. [Google Scholar] [CrossRef]
- Gaston, K.J.; Spicer, J.I. Biodiversity: An Introduction; Wiley: Hoboken, NJ, USA, 2013; p. 208. [Google Scholar]
- Li, Z.; Colinet, G.; Zu, Y.; Wang, J.; An, L.; Li, Q.; Niu, X. Species diversity of Arabis alpina L. communities in two Pb/Zn mining areas with different smelting history in Yunnan Province, China. Chemosphere 2019, 233, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Mohedano-Navarrete, A.; Reyes-Bonilla, H.; López-Pérez, R. Species richness and morphological diversity of the Genus Porites in the Pacific Ocean. In Proceedings of the 11th International Coral Reef Symposium, Ft. Lauderdale, FL, USA, 7–11 July 2008. [Google Scholar]
- Shepherd, U.L.; Kelt, D.A. Mammalian species richness and morphological complexity along an elevational gradient in the arid south-west. J. Biogeogr. 1999, 26, 843–855. [Google Scholar] [CrossRef]
- Lepik, M.; Zobel, K. Is the positive relationship between species richness and shoot morphological plasticity mediated by ramet density or is there a direct link? Oecologia 2015, 178, 867–873. [Google Scholar] [CrossRef] [PubMed]
- Fokin, S.I.; Chivilev, S.M. Paramecium morphometric analysis and taxonomy. Acta Protozool. 2000, 39, 1–14. [Google Scholar]
- Hipsley, C.A.; Miles, D.B.; Müller, J. Morphological disparity opposes latitudinal diversity gradient in lacertid lizards. Biol. Lett. 2014, 10, 20140101. [Google Scholar] [CrossRef]
- Schalk, C.M.; Montaña, C.G.; Springer, L. Morphological diversity and community organization of desert anurans. J. Arid. Environ. 2015, 122, 132–140. [Google Scholar] [CrossRef]
- Kozak, K.H.; Wiens, J.J. What explains patterns of species richness? The relative importance of climatic-niche evolution, morphological evolution, and ecological limits in salamanders. Ecol. Evol. 2016, 6, 5940–5949. [Google Scholar] [CrossRef] [Green Version]
- Pompeo, P.N.; Filho, L.C.I.D.O.; Dos Santos, M.A.B.; Mafra, Á.L.; Baretta, D. Morphological diversity of Coleoptera (Arthropoda: Insecta) in agriculture and forest systems. Revista Brasileira de Ciência do Solo 2017, 41, 41. [Google Scholar] [CrossRef] [Green Version]
- Alhajeri, B.H.; Steppan, S.J. Disparity and Evolutionary Rate Do Not Explain Diversity Patterns in Muroid Rodents (Rodentia: Muroidea). Evol. Biol. 2018, 45, 324–344. [Google Scholar] [CrossRef]
- Souza, I.L.; Tomazella, V.B.; Santos, A.J.N.; Moraes, T.; Silveira, L.C.P. Parasitoids diversity in organic Sweet Pepper (Capsicum annuum) associated with Basil (Ocimum basilicum) and Marigold (Tagetes erecta). Braz. J. Biol. 2019, 79, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Namyatova, A.A.; Contos, P.; Cassis, G. New species, taxonomy, phylogeny, and distribution of the tropical tribe Bothriomirini (Insecta: Heteroptera: Miridae: Cylapinae). Insect Syst. Evol. 2018, 50, 83–146. [Google Scholar] [CrossRef]
- Mindel, B.L.; Neat, F.C.; Trueman, C.N.; Webb, T.J.; Blanchard, J.L. Functional, size and taxonomic diversity of fish along a depth gradient in the deep sea. PeerJ 2016, 4, e2387. [Google Scholar] [CrossRef]
- Gerovasileiou, V.; Voultsiadou, E. Sponge diversity gradients in marine caves of the eastern Mediterranean. J. Mar. Biol. Assoc. United Kingd. 2015, 96, 407–416. [Google Scholar] [CrossRef] [Green Version]
- Neustupa, J.; Černá, K.; Šťastný, J. Diversity and morphological disparity of desmid assemblages in Central European peatlands. Hydrobiology 2009, 630, 243–256. [Google Scholar] [CrossRef] [Green Version]
- Silva, R.R.; Brandão, C.R.F. Ecosystem-Wide Morphological Structure of Leaf-Litter Ant Communities along a Tropical Latitudinal Gradient. PLoS ONE 2014, 9, e93049. [Google Scholar] [CrossRef]
- Safi, K.; Cianciaruso, M.V.; Loyola, R.D.; Brito, D.; Armour-Marshall, K.; Diniz-Filho, J.A.F. Understanding global patterns of mammalian functional and phylogenetic diversity. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2536–2544. [Google Scholar] [CrossRef]
- Kuczynski, L.; Côte, J.; Toussaint, A.; Brosse, S.; Buisson, L.; Grenouillet, G.; Lucie, K.; Jessica, C.; Aurèle, T.; Sébastien, B.; et al. Spatial mismatch in morphological, ecological and phylogenetic diversity, in historical and contemporary European freshwater fish faunas. Ecography 2018, 41, 1665–1674. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Ruan, Y.; Wan, X.; Tong, Y.; Yang, X.; Bai, M. Geometric morphometric analysis of the pronotum and elytron in stag beetles: Insight into its diversity and evolution. ZooKeys 2019, 833, 21–40. [Google Scholar] [CrossRef]
- Tong, Y.; Yang, H.; Ma, D.; Bai, M. Geometric morphometrics evaluation and adaptive evolution study of scarabaeine metendosternites. Acta Entomol. Sin. 2016, 59, 871–879. [Google Scholar]
- Farré, M.; Tuset, V.M.; Maynou, F.; Recasens, L.; Lombarte, A. Geometric morphology as an alternative for measuring the diversity of fish assemblages. Ecol. Indic. 2013, 29, 159–166. [Google Scholar] [CrossRef]
- Akiyama, K.; Ohmono, S. The Buprestid Beetles of the World; Mushi-Sha: Tokyo, Japan, 2000; p. 341. [Google Scholar]
- Bellamy, C.L. The World of Jewel Beetles (Insecta: Coleoptera: Buprestoidea). Available online: https://cerambycids.com/buprestidae/ (accessed on 5 January 2020).
- Bellamy, C.L. A World Catalogue and Bibliography of the Jewel Beetles (Coleoptera: Buprestoidea). Volume 5: Appendices, Bibliography, Indices; Pensoft: Sofia, Bulgaria, 2009; pp. 2689–3264. [Google Scholar]
- Beutel, R.G.; Leschen, R. Volume 1: Morphology and Systematics (Archostemata, Adephaga, Myxophaga, Polyphaga partim); Walter de Gruyter: Berlin, Germany, 2011; p. 567. [Google Scholar]
- Kundrata, R.; Bocak, L. The phylogeny and limits of Elateridae (Insecta, Coleoptera): Is there a common tendency of click beetles to soft-bodiedness and neoteny? Zoöl. Scr. 2011, 40, 364–378. [Google Scholar] [CrossRef]
- Bocakova, M.; Bocak, L.; Hunt, T.; Teraväinen, M.; Vogler, A.P. Molecular phylogenetics of Elateriformia (Coleoptera): Evolution of bioluminescence and neoteny. Cladistics 2007, 23, 477–496. [Google Scholar] [CrossRef]
- Fedorenko, D.N. Evolution of the Beetle Hind Wing, with Special Reference to Folding (Insecta, Coleoptera); Pensoft: Sofia, Bulgaria, 2009; p. 336. [Google Scholar]
- Sherratt, E.; Gower, D.J.; Klingenberg, C.P.; Wilkinson, M. Evolution of Cranial Shape in Caecilians (Amphibia: Gymnophiona). Evol. Biol. 2014, 41, 528–545. [Google Scholar] [CrossRef]
- Bai, M.; Li, S.; Lu, Y.Y.; Yang, H.D.; Tong, Y.J.; Yang, X.K. Mandible evolution in the Scarabaeinae (Coleoptera: Scarabaeidae) and adaptations to coprophagous habits. Front. Zoöl. 2015, 12, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kukalova-Peck, J.; Lawrence, J.F. Evolution of the hind wing in Coleoptera. Can. Entomol. 1993, 125, 181–258. [Google Scholar] [CrossRef] [Green Version]
- Evans, A.M.; McKenna, D.D.; Bellamy, C.L.; Farrell, B.D. Large-scale molecular phylogeny of metallic wood-boring beetles (Coleoptera: Buprestoidea) provides new insights into relationships and reveals multiple evolutionary origins of the larval leaf-mining habit. Syst. Èntomol. 2015, 40, 385–400. [Google Scholar] [CrossRef]
- Kuban, V.; Majer, K.; Kolibac, J. Classification of the tribe Coraebini Bedel, 1921 (Coleoptera, Buprestidae, Agrilinae). Acta Musei Moraviae Scientiae Biologicae 2000, 85, 185–287. [Google Scholar]
- Jendek, E.; Grebennikov, V. Agrilus (Coleoptera, Buprestidae) of East Asia; Nakladatelství Jan Farkač: Prague, Czech Republic, 2011; p. 362. [Google Scholar]
- Bellamy, C.L. An illustrated summary of the higher classification of the superfamily Buprestoidea (Coleoptera). Folia Heyrovskyana 2003, 10, 197. [Google Scholar]
- Duan, J.J.; Schmude, J.M.; Larson, K.M.; Fuester, R.W.; Gould, J.R.; Ulyshen, M.D. Field parasitism and host specificity of Oobius primorskyensis (Hymenoptera: Encyrtidae), an egg parasitoid of the emerald ash borer (Coleoptera: Buprestidae) in the Russian Far East. Biol. Control. 2019, 130, 44–50. [Google Scholar] [CrossRef]
- Sun, H.; Zhao, W.; Lin, R.; Zhou, Z.; Huai, W.; Yao, Y. The conserved mitochondrial genome of the jewel beetle (Coleoptera: Buprestidae) and its phylogenetic implications for the suborder Polyphaga. Genomics 2020, 112, 3713–3721. [Google Scholar] [CrossRef] [PubMed]
- Arnett, R.H.; Thomas, M.C.; Skelley, P.E.; Frank, J.H. American Beetles, Volume II: Polyphaga: Scarabaeoidea through Curculionoidea; CRC press: Boca Raton, FL, USA, 2002; p. 861. [Google Scholar]
- Green, S.B.; Salkind, N.J. Using SPSS for Windows and Macintosh, Books a la Carte, 8th ed.; Pearson: London, UK, 2016; p. 352. [Google Scholar]
- Chiari, S.; Zauli, A.; Audisio, P.A.; Campanaro, A.; Donzelli, P.F.; Romiti, F.; Svensson, G.P.; Tini, M.; Carpaneto, G.M. Monitoring presence, abundance and survival probability of the stag beetle, Lucanus cervus, using visual and odour-based capture methods: Implications for conservation. J. Insect Conserv. 2014, 18, 99–109. [Google Scholar] [CrossRef]
- Evans, M.E.G. Locomotion in the Coleoptera Adephaga, especially Carabidae. J. Zoöl. 2009, 181, 189–226. [Google Scholar] [CrossRef]
- Chapman, R.F.; Simpson, S.J.; Douglas, A.E. The Insects: Structure and Function, 7th ed.; Cambridge University Press: Cambridge, UK, 1998; p. 929. [Google Scholar]
- Snodgrass, R.E. Principles of Insect Morphology; Cornell University Press: Itahca, NY, USA, 2018; p. 667. [Google Scholar]
- Van de Kamp, T.; Greven, H. On the architecture of beetle elytra. Entomologie Heute 2010, 22, 191–204. [Google Scholar]
- Volkovitsh, M.G. Galbella (Galbella) holzschuhi sp. nov. from Iran (Coleoptera: Buprestidae). Folia Heyrovskyana A 2008, 16, 39–48. [Google Scholar]
- Bellamy, C.; Willams, G. The first Australian Paratrachys (Coleptera: Buprestidae), with comments on the higher classification of the genus. Invertebr. Syst. 1995, 9, 1265–1276. [Google Scholar] [CrossRef]
- Bellamy, C.L.; Holm, E. Revision of the African Species of Galbella (Coleoptera: Buprestidae); Entomology Memoir, Department of Agriculture and Water Supply of Republic of South Africa: Pretoria, South Africa, 1986; p. 39. [Google Scholar]
- Bouchard, P.; Bousquet, Y.; Davies, A.E.; Alonso-Zarazaga, M.A.; Lawrence, J.F.; Lyal, C.H.C.; Newton, A.F.; Reid, C.A.M.; Schmitt, M.; Ślipiński, S.A.; et al. Family-group names in Coleoptera (Insecta). ZooKeys 2011, 88, 1–972. [Google Scholar] [CrossRef] [Green Version]
- Rohlf, F.J. tpsDig, Digitize Landmarks and Outlines, version 2.05. [Software and Manual] Department of Ecology and Evolution, State University of New York at Stony Brook, NY, USA. Available online: http://life.bio.sunysb.edu/morph/ (accessed on 22 December 2020).
- MacLeod, N. Morphometrics: History, development methods and prospects. Zool. Syst. 2017, 42, 4–33. [Google Scholar] [CrossRef]
- Shi, Y.; MacLeod, N. Identification of life-history stages in fusulinid foraminifera. Mar. Micropaleontol. 2016, 122, 87–98. [Google Scholar] [CrossRef]
- MacLeod, N. Form & shape models. Palaeontol. Assoc. Newsl. 2009, 72, 14–27. [Google Scholar]
- Klingenberg, C.P. MorphoJ: An integrated software package for geometric morphometrics. Mol. Ecol. Resour. 2011, 11, 353–357. [Google Scholar] [CrossRef]
- Triantis, K.A.; Rigal, F.; Parent, C.E.; Cameron, R.A.D.; Lenzner, B.; Parmakelis, A.; Yeung, N.W.; Alonso, M.R.; Ibáñez, M.; Martins, A.M.D.F.; et al. Discordance between morphological and taxonomic diversity: Land snails of oceanic archipelagos. J. Biogeogr. 2016, 43, 2050–2061. [Google Scholar] [CrossRef]
- Roy, K.; Foote, M. Morphological approaches to measuring biodiversity. Trends Ecol. Evol. 1997, 12, 277–281. [Google Scholar] [CrossRef]
- Venner, S.; Pélisson, P.-F.; Bel-Venner, M.-C.; Débias, F.; Rajon, E.; Menu, F. Coexistence of insect species competing for a pulsed resource: Toward a unified theory of biodiversity in fluctuatingenvironments. PLoS ONE 2011, 6, e18039. [Google Scholar] [CrossRef] [PubMed]
- Carroll, S.B. The Making of the Fittest: DNA and the Ultimate Forensic Record of Evolution; W.W. Norton & Company: London, UK, 2009; p. 288. [Google Scholar]
- Gory, H.L. Histoire Naturelle et Iconographie des Insectes Coléoptères. Supplement aux Buprestides; P. Duménil: Paris, France, 1841; pp. 73–77. [Google Scholar]
- Bellamy, C.L. Phylogenetic relationships of Xyroscelis (Coleoptera: Buprestidae). Invertebr. Syst. 1997, 11, 569. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, F.S. Ecomorphological relationships in hoverflies (Diptera, Syrphidae). Proc. R. Soc. Lond. Ser. B Boil. Sci. 1985, 224, 91–105. [Google Scholar] [CrossRef]
- Collar, D.C.; Near, T.J.; Wainwright, P.C. Comparative analysis of morphological diversity: Does disparity accumulate at the same rate in two lineages of centrarchid fishes? Evolution 2005, 59, 1783–1794. [Google Scholar] [CrossRef]
- Bellamy, C.L. World Catalogue & Bibliography of the Jewel Beetles (Coleoptera, Buprestoidea). Volume 3, Buprestinae, Pterobothrini through Agrilinae, Rhaeboscelina (Pensoft Series Faunistica; No. 78); Pensoft: Sofia, Bulgaria, 2008; p. 676. [Google Scholar]
- Jendek, E. A comparative study of the abdomen in the family Buprestidae (Coleoptera). Acta Musei Moraviae Scientiae Biologicae 2001, 86, 1–41. [Google Scholar]
- Migliore, L.J.; Biffi, G.; Curletti, G. A revision of Cylindromorphoidina: Comparative morphology, genera reassessment and description of new species (Coleoptera, Buprestidae, Agrilinae). Insect Syst. Evol. 2020, 1, 1–41. [Google Scholar] [CrossRef]
- Herms, D.A.; McCullough, D.G. Emerald Ash Borer invasion of North America: History, biology, ecology, impacts, and management. Annu. Rev. Entomol. 2014, 59, 13–30. [Google Scholar] [CrossRef] [Green Version]
- Jendek, E.; Grebennikov, V.V. Twenty new species of Agrilus (Coleoptera: Buprestidae: Agrilinae) from the Oriental Region. Zootaxa 2018, 4429, 107–131. [Google Scholar] [CrossRef] [PubMed]
- Rao, C.R. The utilization of multiple measurements in problems of biological classification. J. R. Stat. Soc. B. (Methodol.) 1948, 10, 159–203. [Google Scholar] [CrossRef]
- Fisher, R.A. The use of multiple measurements in taxonomic problems. Ann. Eugenics 1936, 7, 179–188. [Google Scholar] [CrossRef]
- Bai, M.; Yang, X.; Li, J.; Wang, W. Geometric Morphometrics, a super scientific computing tool in morphology comparison. Chin. Sci. Bull. 2014, 59, 887–894. [Google Scholar] [CrossRef]
- Pigot, A.L.; Trisos, C.H.; Tobias, J.A. Functional traits reveal the expansion and packing of ecological niche space underlying an elevational diversity gradient in passerine birds. Proc. R. Soc. B Boil. Sci. 2016, 283, 20152013. [Google Scholar] [CrossRef] [Green Version]
- Adams, D.C.; Rohlf, F.J.; Slice, D.E. Geometric morphometrics: Ten years of progress following the ‘revolution’. Ital. J. Zoöl. 2004, 71, 5–16. [Google Scholar] [CrossRef] [Green Version]
- Ricklefs, R.E. Species richness and morphological diversity of passerine birds. Proc. Natl. Acad. Sci. USA 2012, 109, 14482–14487. [Google Scholar] [CrossRef] [Green Version]
- Bookstein, F.L. Morphometric Tools for Landmark Data: Geometry and Biology; Cambridge University Press: Cambridge, UK, 1997; pp. 55–87. [Google Scholar]
- Jolicoeur, P.; Mosimann, J.E. Size and shape variation in the painted turtle. A principal component analysis. Growth 1960, 24, 339–354. [Google Scholar]
- Neige, P. Spatial patterns of disparity and diversity of the recent cuttlefishes (Cephalopoda) across the Old World. J. Biogeogr. 2003, 30, 1125–1137. [Google Scholar] [CrossRef]
- Bertrand, Y.; Pteijel, F.; Rouse, G.W. Taxonomic surrogacy in biodiversity assessments, and the meaning of Linnaean ranks. Syst. Biodivers. 2006, 4, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.S.Y. Species concepts and species reality: Salvaging a Linnaean rank. J. Evolution. Biol. 2003, 16, 179–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walters, S.M. The shaping of angiosperm taxonomy. New Phytol. 1961, 60, 74–84. [Google Scholar] [CrossRef]
- Williams, P.H.; Gaston, K.J. Measuring more of biodiversity—Can higher-taxon richness predict wholesale species richness. Biol. Conserv. 1994, 67, 211–217. [Google Scholar] [CrossRef]
- Dial, K.P.; Marzluff, J.M. Nonrandom diversification with in taxonomic assemblages. Syst. Biol. 1989, 38, 26–37. [Google Scholar] [CrossRef]
- Glazier, D.S. Energetics and taxonomic patterns of species diversity. Syst. Zool. 1987, 36, 62–71. [Google Scholar] [CrossRef]
- Wright, S. Review of The “age and area” concept extended. Ecology 1941, 22, 345–347. [Google Scholar] [CrossRef]
- Stebbins, G.L.; Johnson, R.H.; Haines, W.J. The Major Features of Evolution. George Gaylord Simpson. Columbia Univ. Press, New York, 1953. 434 pp. Illus. $7.50. Science 2006, 119, 699–701. [Google Scholar] [CrossRef]
- Alroy, J. Geographical, environmental and intrinsic biotic controls on Phanerozoic marine diversification. Palaeontol. 2010, 53, 1211–1235. [Google Scholar] [CrossRef]
- Maruvka, Y.E.; Shnerb, N.M.; Kessler, D.A.; Ricklefs, R.E. Model for macroevolutionary dynamics. Proc. Natl. Acad. Sci. USA 2013, 110, E2460–E2469. [Google Scholar] [CrossRef] [Green Version]
- Jablonski, D.; Finarelli, J.A. Congruence of morphologically-defined genera with molecular phylogenies. Proc. Natl. Acad. Sci. USA 2009, 106, 8262–8266. [Google Scholar] [CrossRef] [Green Version]
- Hillebrand, H. On the Generality of the Latitudinal Diversity Gradient. Am. Nat. 2004, 163, 192–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahbek, C. The elevational gradient of species richness: A uniform pattern? Ecography 1995, 18, 200–205. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2012; p. 880. [Google Scholar]
- Shepherd, U. A comparison of species diversity and morphological diversity across the North American latitudinal gradient. J. Biogeogr. 1998, 25, 19–29. [Google Scholar] [CrossRef]
- Legendre, P.; Mi, X.; Ren, H.; Ma, K.; Yu, M.; Sun, I.-F.; He, F. Partitioning beta diversity in a subtropical broad-leaved forest of China. Ecology 2009, 90, 663–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuomisto, H.; Ruokolainen, K.; Yli-Halla, M. Dispersal, environment, and floristic variation of Western Amazonian forests. Science 2003, 299, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Arnan, X.; Cerdá, X.; Retana, J. Partitioning the impact of environment and spatial structure on alpha and beta components of taxonomic, functional, and phylogenetic diversity in European ants. PeerJ 2015, 3, e1241. [Google Scholar] [CrossRef] [Green Version]
- Whittaker, R.J.; Willis, K.J.; Field, R. Scale and species richness: Towards a general, hierarchical theory of species diversity. J. Biogeogr. 2001, 28, 453–470. [Google Scholar] [CrossRef] [Green Version]
- Innes, J.L. Measuring environmental change. In Ecological Scale: Theory and Applications; Peterson, D.L., Parker, V.T., Eds.; Columbia University Press: New York City, NY, USA, 1998; pp. 429–457. [Google Scholar]
- Cornwell, W.K.; Schwilk, D.W.; Ackerly, D.D. A trait-based test for habitat filtering: Convex hull volume. Ecology 2006, 87, 1465–1471. [Google Scholar] [CrossRef]
- Gaston, K.J. Biodiversity–latitudinal gradients. Prog. Phys. Geogr. Earth Environ. 1996, 20, 466–476. [Google Scholar] [CrossRef]
- Fox, J.W. The intermediate disturbance hypothesis should be abandoned. Trends Ecol. Evol. 2013, 28, 86–92. [Google Scholar] [CrossRef]
- Cavender-Bares, J.; Kozak, K.H.; Fine, P.V.A.; Kembel, S.W. The merging of community ecology and phylogenetic biology. Ecol. Lett. 2009, 12, 693–715. [Google Scholar] [CrossRef] [PubMed]
- Slingsby, J.A.; Verboom, G.A. Phylogenetic relatedness limits co-occurrence at fine spatial scales: Evidence from the Schoenoid Sedges (Cyperaceae: Schoeneae) of the Cape Floristic Region, South Africa. Am. Nat. 2006, 168, 14–27. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Groups | Sampling Number of Genus | Sampling Number of Species | Described Number of Genus | Described Number of Species |
|---|---|---|---|---|
| Polycestinae | 26 | 120 | 82 | 1255 |
| Chrysochroinae | 34 | 323 | 112 | 2742 |
| Buprestinae | 35 | 258 | 114 | 3306 |
| Agrilinae | 71 | 318 | 203 | 7196 |
| Julodinae | 6 | 78 | 6 | 141 |
| Galbellinae | 1 | 45 | 1 | 84 |
| Total | 173 | 1106 | 518 | 14,724 |
| Test Groups | MD of Pronotum | MD of Elytron |
|---|---|---|
| Polycestinae | 0.0150 | 0.0057 |
| Chrysochroinae | 0.0070 | 0.0026 |
| Buprestinae | 0.0091 | 0.0026 |
| Agrilinae | 0.0187 | 0.0062 |
| Julodinae | 0.0070 | 0.0009 |
| Galbellinae | 0.0046 | 0.0011 |
| Genus Richness (P) | Species Richness (P) | |
|---|---|---|
| MD of Pronotum | 0.714 | 0.371 |
| MD of Elytron | 0.771 | 0.543 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tong, Y.-J.; Yang, H.-D.; Jenkins Shaw, J.; Yang, X.-K.; Bai, M. The Relationship between Genus/Species Richness and Morphological Diversity among Subfamilies of Jewel Beetles. Insects 2021, 12, 24. https://doi.org/10.3390/insects12010024
Tong Y-J, Yang H-D, Jenkins Shaw J, Yang X-K, Bai M. The Relationship between Genus/Species Richness and Morphological Diversity among Subfamilies of Jewel Beetles. Insects. 2021; 12(1):24. https://doi.org/10.3390/insects12010024
Chicago/Turabian StyleTong, Yi-Jie, Hai-Dong Yang, Josh Jenkins Shaw, Xing-Ke Yang, and Ming Bai. 2021. "The Relationship between Genus/Species Richness and Morphological Diversity among Subfamilies of Jewel Beetles" Insects 12, no. 1: 24. https://doi.org/10.3390/insects12010024
APA StyleTong, Y. -J., Yang, H. -D., Jenkins Shaw, J., Yang, X. -K., & Bai, M. (2021). The Relationship between Genus/Species Richness and Morphological Diversity among Subfamilies of Jewel Beetles. Insects, 12(1), 24. https://doi.org/10.3390/insects12010024