
Pollination in the Rainforest: Scarce Visitors and Low Effective Pollinators Limit the Fruiting Success of Tropical Orchids




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Species and Sites
2.2. Breeding System
2.3. Flower Visitors
3. Results
3.1. Breeding System
3.2. Flower Visitors
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ollerton, J.; Price, V.; Armbruster, W.S.; Memmott, J.; Watts, S.; Waser, N.M.; Totland, Ø.; Goulson, D.; Alarcón, R.; Stout, J.C.; et al. Overplaying the role of honey bees as pollinators: A comment on Aebi and Neumann (2011). Trends Ecol. Evol. 2012, 27, 141–142. [Google Scholar] [CrossRef]
- Jiang, X.; Deng, H.N.; Qu, J.; Zeng, H.; Cao, G.X. The function of extrafloral nectaries in the pollination and reproduction of Sambucus javanica. Plant Species Biol. 2019, 34, 53–60. [Google Scholar] [CrossRef]
- Maloof, J.E.; Inouye, D.W. Are nectar robbers cheaters or mutualists? Ecology 2000, 81, 2651–2661. [Google Scholar] [CrossRef]
- Genini, J.; Morellato, L.P.C.; Guimarães, P.R.; Olesen, J.M. Cheaters in mutualism networks. Biol. Lett. 2010, 6, 494–497. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zych, M. Pollination biology of Heracleum sphondylium L. (Apiaceae). The advantages of being white and compact. Acta Soc. Bot. Pol. 2002, 71, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Fumero-Cabán, J.J.; Meléndez-Ackerman, E.J. Relative pollination effectiveness of floral visitors of Pitcairnia angustifolia (Bromeliaceae). Am. J. Bot. 2007, 94, 419–424. [Google Scholar] [CrossRef] [PubMed]
- Watts, S.; Ovalle, D.H.; Herrera, M.M.; Ollerton, J. Pollinator effectiveness of native and non-native flower visitors to an apparently generalist Andean shrub, Duranta mandonii (Verbenaceae). Plant Species Biol. 2012, 27, 147–158. [Google Scholar] [CrossRef]
- Wilson, P.; Thomson, J.D. Heterogeneity among floral visitors leads to discordance between removal and deposition of pollen. Ecology 1991, 72, 1503–1507. [Google Scholar] [CrossRef]
- Zych, M.; Goldstein, J.; Roguz, K.; Stpiczyńska, M. The most effective pollinator revisited: Pollen dynamics in a spring-flowering herb. Arthropod. Plant. Interact. 2013, 7, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Singer, R.B.; Sazima, M. The pollination of Stenorrhynchos lanceolatus (Aublet) L. C. Rich. (Orchidaceae: Spiranthinae) by hummingbirds in southeastern Brazil. Plant Syst. Evol. 2000, 223, 221–227. [Google Scholar] [CrossRef]
- Sugiura, N. Specialized pollination by carpenter bees in Calanthe striata (Orchidaceae), with a review of carpenter bee pollination in orchids. Bot. J. Linn. Soc. 2013, 171, 730–743. [Google Scholar] [CrossRef] [Green Version]
- Johnson, S.D. Evidence for Batesian mimicry in a butterfly-pollinated orchid. Biol. J. Linn. Soc. 1994, 53, 91–104. [Google Scholar] [CrossRef]
- Suetsugu, K.; Tanaka, K. Diurnal butterfly pollination in the orchid Habenaria radiata. Entomol. Sci. 2014, 17, 443–445. [Google Scholar] [CrossRef]
- Pansarin, E.R.; Pansarin, L.M. Crane flies and microlepidoptera also function as pollinators in Epidendrum (Orchidaceae: Laeliinae): The reproductive biology of E. avicula. Plant Species Biol. 2017, 32, 200–209. [Google Scholar] [CrossRef]
- Houlihan, P.R.; Stone, M.; Clem, S.E.; Owen, M.; Emmel, T.C. Pollination ecology of the ghost orchid (Dendrophylax lindenii): A first description with new hypotheses for Darwin’s orchids. Sci. Rep. 2019, 9, 12850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schiestl, F.P.; Glaser, F. Specific ant-pollination in an alpine orchid and the role of floral scent in attracting pollinating ants. Alp. Bot. 2012, 122, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Dodson, C. The Importance of pollination in the evolution of the orchids of tropical America. Am. Orchid Soc. Bull. 1962, 31, 731–735. [Google Scholar]
- Van der Pijl, L.; Dodson, C.H. Orchid Flowers: Their Pollination and Evolution; University of Miami Press: Miami, FL, USA, 1966. [Google Scholar]
- Dafni, A. Mimicry and deception in pollination. Annu. Rev. Ecol. Syst. 1984, 15, 259–278. [Google Scholar] [CrossRef]
- Jersáková, J.; Johnson, S.D.; Kindlmann, P. Mechanisms and evolution of deceptive pollination in orchids. Biol. Rev. Camb. Philos. Soc. 2006, 81, 219–235. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Li, D.; Ren, Z.; Xiang, X. A generalized deceptive pollination system of Doritis pulcherrima (Aeridinae: Orchidaceae) with non-reconfigured pollinaria. BMC Plant Biol. 2012, 12, 67. [Google Scholar] [CrossRef] [Green Version]
- Neiland, M.R.M.; Wilcock, C.C. Fruit set, nectar reward, and rarity in the Orchidaceae. Am. J. Bot. 1998, 85, 1657–1671. [Google Scholar] [CrossRef]
- Tremblay, R.L.; Ackerman, J.D.; Zimmerman, J.K.; Calvo, R.N. Variation in sexual reproduction in orchids and its evolutionary consequences: A spasmodic journey to diversification. Biol. J. Linn. Soc. 2005, 84, 1–54. [Google Scholar] [CrossRef]
- Marques, I.; Draper, D.; Riofrío, L.; Naranjo, C. Multiple hybridization events, polyploidy and low postmating isolation entangle the evolution of neotropical species of Epidendrum (Orchidaceae). BMC Evol. Biol. 2014, 14, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Liu, Q.; Han, J.Y.; Gao, J.Y. Different pollinator assemblages ensure reproductive success of Cleisostoma linearilobatum (Orchidaceae) in fragmented holy hill forest and traditional tea garden. Sci. Rep. 2016, 6, 21435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caballero-Villalobos, L.; Silva-Arias, G.A.; Buzatto, C.R.; Nervo, M.H.; Singer, R.B. Generalized food-deceptive pollination in four Cattleya (Orchidaceae: Laeliinae) species from Southern Brazil. Flora Morphol. Distrib. Funct. Ecol. Plants 2017, 234, 195–206. [Google Scholar] [CrossRef]
- Dupont, Y.L.; Padrón, B.; Olesen, J.M.; Petanidou, T. Spatio-temporal variation in the structure of pollination networks. Oikos 2009, 118, 1261–1269. [Google Scholar] [CrossRef]
- Hegland, S.J.; Dunne, J.; Nielsen, A.; Memmott, J. How to monitor ecological communities cost-efficiently: The example of plant-pollinator networks. Biol. Conserv. 2010, 143, 2092–2101. [Google Scholar] [CrossRef]
- Kaiser-Bunbury, C.N.; Muff, S.; Memmott, J.; Müller, C.B.; Caflisch, A. The robustness of pollination networks to the loss of species and interactions: A quantitative approach incorporating pollinator behaviour. Ecol. Lett. 2010, 13, 442–452. [Google Scholar] [CrossRef] [Green Version]
- Burkle, L.A.; Alarcón, R. The future of plant-pollinator diversity: Understanding interaction networks acrosss time, space, and global change. Am. J. Bot. 2011, 98, 528–538. [Google Scholar] [CrossRef]
- Olesen, J.M.; Bascompte, J.; Dupont, Y.L.; Jordano, P. The modularity of pollination networks. Proc. Natl. Acad. Sci. USA 2007, 104, 19891–19896. [Google Scholar] [CrossRef] [Green Version]
- Alarcón, R. Congruence between visitation and pollen-transport networks in a California plant-pollinator community. Oikos 2010, 119, 35–44. [Google Scholar] [CrossRef]
- Vicens, N.; Bosch, J. Weather-dependent pollinator activity in an apple orchard, with special reference to Osmia cornuta and Apis mellifera (Hymenoptera: Megachilidae and Apidae). Environ. Entomol. 2000, 29, 413–420. [Google Scholar] [CrossRef]
- Tuell, J.K.; Isaacs, R. Weather during bloom affects pollination and yield of highbush blueberry. J. Econ. Entomol. 2010, 103, 557–562. [Google Scholar] [CrossRef] [PubMed]
- Vega, Y.; Marques, I. Both biotic and abiotic factors influence floral longevity in three species of Epidendrum (orchidaceae). Plant Species Biol. 2014, 30, 184–192. [Google Scholar] [CrossRef]
- Bawa, K.S.; Kang, H.; Grayum, M.H. Relationships among time, frequency, and duration of flowering in tropical rain forest trees. Am. J. Bot. 2003, 90, 877–887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bawa, K.S. Plant-pollinator interactions in tropical rain forests. Annu. Rev. Ecol. Syst. 1990, 21, 399–422. [Google Scholar] [CrossRef]
- Ruilova, X.R.; Marques, I. Better common than rare? Effects of low reproductive success, scarce pollinator visits and interspecific gene flow in threatened and common species of Tibouchina (Melastomataceae). Plant Species Biol. 2016, 31, 288–295. [Google Scholar] [CrossRef]
- Vega, Y.; Marques, I.; Castro, S.; Loureiro, J. Outcomes of extensive hybridization and introgression in Epidendrum (orchidaceae): Can we rely on species boundaries? PLoS ONE 2013, 8, e80662. [Google Scholar] [CrossRef] [Green Version]
- MAE, Ministerio de Ambiente del Ecuador. Estadísticas de Patrimonio Natural Ecuador continental. In Datos de Bosques, Ecosistemas, Especies, Carbono y Deforestación del Ecuador Continental; Unidad de Procesamiento de Información y Geomática: Quito, Ecuador, 2015. [Google Scholar]
- Macedo, M.N.; DeFries, R.S.; Morton, D.C.; Stickler, C.M.; Galford, G.L.; Shimabukuro, Y.E. Decoupling of deforestation and soy production in the southern Amazon during the late 2000s. Proc. Natl. Acad. Sci. USA 2012, 109, 1341–1346. [Google Scholar] [CrossRef] [Green Version]
- Tapia-Armijos, M.F.; Homeier, J.; Espinosa, C.I.; Leuschner, C.; De La Cruz, M. Deforestation and forest fragmentation in south Ecuador since the 1970s—Losing a hotspot of biodiversity. PLoS ONE 2015, 10, e0133701. [Google Scholar] [CrossRef] [Green Version]
- Vega, Y. Evolucion de las Barreras Reproductivas en el Género Neotropical Epidendrum (Orchidaceae) en Ecuador; Universidad Técnica Particular de Loja: Loja, Ecuador, 2011. [Google Scholar]
- Larson, B.M.H.; Barrett, S.C.H. A comparative analysis of pollen limitation in flowering plants. Biol. J. Linn. Soc. 2000, 69, 503–520. [Google Scholar] [CrossRef]
- IBM Corp. Version 23.0. IBM SPSS Statistics for Windows; IBM Corp.: Armonk, NY, USA, 2015. [Google Scholar]
- Dafni, A.; Ivri, Y. Pollintation ecology of, and hybridization between, Orchis coriophora L. and O. collina sol. Ex russ. (orchidaceae) in Israel. New Phytol. 1979, 83, 181–187. [Google Scholar] [CrossRef]
- Dressler, R.L. Biology of the orchid bees (Euglossini). Annu. Rev. Ecol. Syst. 1982, 13, 373–394. [Google Scholar] [CrossRef]
- Waterman, R.J.; Bidartondo, M.I. Deception above, deception below: Linking pollination and mycorrhizal biology of orchids. J. Exp. Bot. 2008, 59, 1085–1096. [Google Scholar] [CrossRef] [PubMed]
- Cozzolino, S.; Widmer, A. Orchid diversity: An evolutionary consequence of deception? Trends Ecol. Evol. 2005, 20, 487–494. [Google Scholar] [CrossRef] [PubMed]
- Pansarin, E.R.; Amaral, M.C.E. Reproductive biology and pollination mechanisms of Epidendrum secundum (Orchidaceae). Floral variation: A consequence of natural hybridization? Plant Biol. 2008, 10, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Sonkoly, J.; Vojtkó, A.E.; Tökölyi, J.; Török, P.; Sramkó, G.; Illyés, Z.; Molnár, V.A. Higher seed number compensates for lower fruit set in deceptive orchids. J. Ecol. 2016, 104, 343–351. [Google Scholar] [CrossRef] [Green Version]
- Johnson, S.D.; Peter, C.I.; Ågren, J. The effects of nectar addition on pollen removal and geitonogamy in the non-rewarding orchid Anacamptis morio. Proc. R. Soc. B Biol. Sci. 2004, 271, 803–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva-Pereira, V.; De Camargo Smidt, E.; Leite Borba, E. Isolation mechanisms between two sympatric Sophronitis (Orchidaceae) species endemic to Northeastern Brazil. Plant Syst. Evol. 2007, 269, 171–182. [Google Scholar] [CrossRef]
- Cuevas, E.; Espino, J.; Marques, I. Reproductive isolation between Salvia elegans and S. fulgens, two hummingbird-pollinated sympatric sages. Plant Biol. 2018, 20, 1075–1082. [Google Scholar] [CrossRef]
- Meyer, B.; Kröger, J.; Steffan-Dewenter, I. Contribution of diurnal and nocturnal pollinators to the reproductive success of the orchid species Gymnadenia conopsea. Entomol. Gen. 2007, 30, 299–300. [Google Scholar] [CrossRef]
- Sletvold, N.; Ågren, J. Pollinator-mediated selection on floral display and spur length in the orchid Gymnadenia conopsea. Int. J. Plant Sci. 2010, 171, 999–1009. [Google Scholar] [CrossRef] [Green Version]
- Duque-Buitrago, C.A.; Alzate-Quintero, N.F.; Otero, J.T. Nocturnal pollination by fungus gnats of the colombian endemic species, Pleurothallis marthae (Orchidaceae: Pleurothallidinae). Lankesteriana 2014, 13, 407–417. [Google Scholar] [CrossRef] [Green Version]
- Ackerman, J.D.; Montalvo, A.M. Short- and long-term limitations to fruit production in a tropical orchid. Ecology 1990, 71, 263–272. [Google Scholar] [CrossRef]
- Cunningham, S.A. Effects of habitat fragmentation on the reproductive ecology of four plant species in mallee woodland. Conserv. Biol. 2000, 14, 758–768. [Google Scholar] [CrossRef]
- Aguilar, R.; Ashworth, L.; Galetto, L.; Aizen, M.A. Plant reproductive susceptibility to habitat fragmentation: Review and synthesis through a meta-analysis. Ecol. Lett. 2006, 9, 968–980. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.J.; Mayfield, M.M. The effect of habitat fragmentation on the bee visitor assemblages of three Australian tropical rainforest tree species. Ecol. Evol. 2018, 8, 8204–8216. [Google Scholar] [CrossRef] [PubMed] [Green Version]




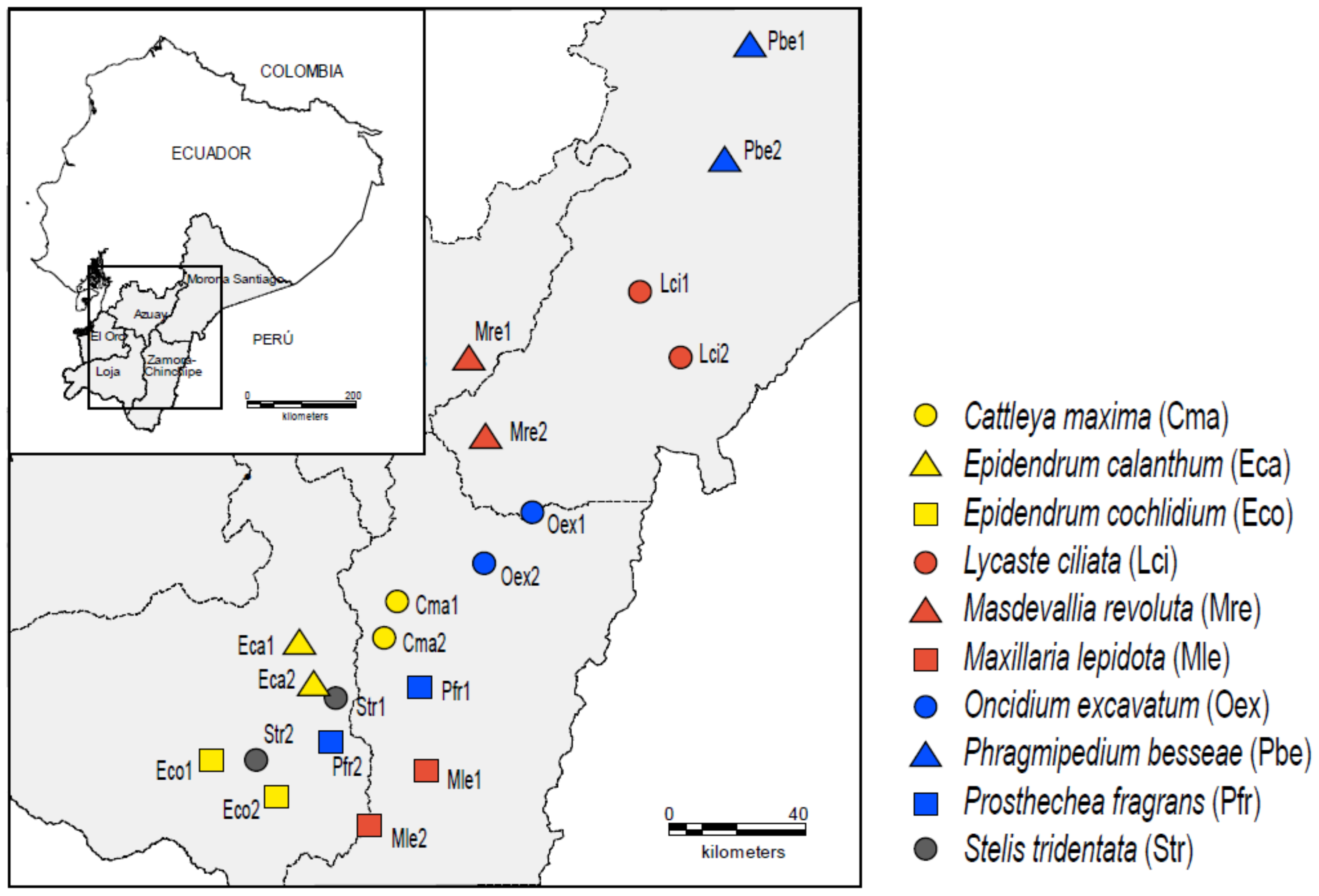{kind=link}
{kind=link}
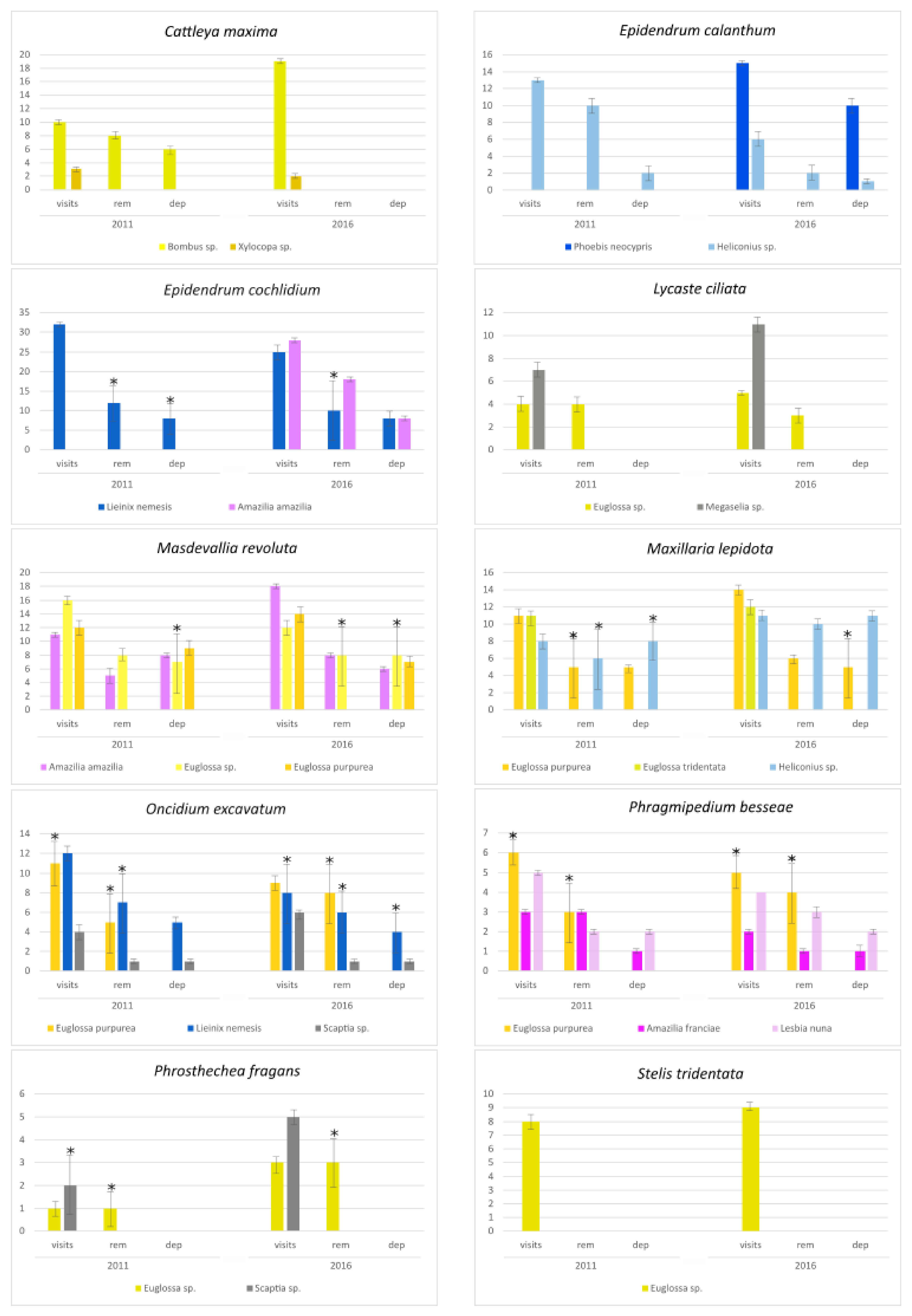{kind=link}
| Orchids | Naturally- Pollinated | Self- Pollinated | Geitonogamous- Pollinated | Cross- Pollinated | PL |
|---|---|---|---|---|---|
| Cattleya maxima | 0.40 ± 0.12 a | 0.42 ± 0.22 a | 0.48 ± 0.18 b | 0.88 ± 0.24 c | 0.55 |
| Epidendrum calanthum | 0.33 ± 0.15 a | 0.37 ± 0.18 b | 0.45 ± 0.18 c | 0.87 ± 0.23 d | 0.62 |
| Epidendrum cochlidium | 0.42 ± 0.11 b | 0.35 ± 0.13 a | 0.59 ± 0.11 c | 0.89 ± 0.20 c | 0.52 |
| Lycaste ciliata | 0.22 ± 0.15 a | 0.20 ± 0.16 a | 0.38 ± 0.12 b | 0.69 ± 0.25 c | 0.68 |
| Masdevallia revoluta | 0.29 ± 0.17 b | 0.20 ± 0.19 a | 0.58 ± 0.11 c | 0.81 ± 0.22 d | 0.64 |
| Maxillaria lepidota | 0.25 ± 0.19 b | 0.17 ± 0.15 a | 0.47 ± 0.18 c | 0.83 ± 0.20 c | 0.69 |
| Oncidium excavatum | 0.34 ± 0.16 b | 0.21 ± 0.18 a | 0.31 ± 0.17 b | 0.79 ± 0.22 c | 0.56 |
| Phragmipedium besseae | 0.20 ± 0.09 a | 0.19 ± 0.10 a | 0.25 ± 0.12 b | 0.68 ± 0.31 c | 0.71 |
| Prosthechea fragrans | 0.18 ± 0.09 a | 0.19 ± 0.11 a | 0.32 ± 0.13 b | 0.61 ± 0.34 c | 0.70 |
| Stelis tridentata | 0.15 ± 0.10 a | 0.14 ± 0.12 a | 0.58 ± 0.15 b | 0.73 ± 0.56 c | 0.79 |
| Orchids | Habit | Flower Colour | Pollinator Group | Flower Visitors | Visit Duration (s) | % of Visits to Other Flowers |
|---|---|---|---|---|---|---|
| Cattleya maxima | Epiphytic | Pink | Bee | Bombus sp. | 8–12 (7–12) | 40% |
| Bee | Xylocopa sp. | 5–9 (5–10) | - | |||
| Epidendrum calanthum | Terrestrial | Pink | Butterfly | Phoebis neocypris | - (11–15) | - |
| Butterfly | Heliconius sp. | 12–19 (13–18) | 20% | |||
| Epidendrum cochlidium | Terrestrial | Red | Butterfly | Lieinix nemesis | 14–18 (11–16) | 20% |
| Hummingbird | Amazilia amazilia | - (5–8) | - | |||
| Lycaste ciliata | Epiphytic | Green-yellow | Bee | Euglossa sp. | 9–12 (9–11) | - |
| Fly | Megaselia sp. | 10–14 (8–13) | - | |||
| Masdevallia revoluta | Epiphytic | Red | Hummingbird | Amazilia amazilia | 5–8 (3–6) | - |
| Bee | Euglossa sp. | 7–10 (8–12) | 50% | |||
| Bee | Euglossa purpurea | 8–11(9–14) | 30% | |||
| Maxillaria lepidota | Epiphytic | Yellow | Bee | Euglossa purpurea | 8–13 (9–15) | 40% |
| Bee | Euglossa tridentata | 9–15 (11–17) | 30% | |||
| Butterfly | Heliconius sp. | 11–15 (14–18) | - | |||
| Oncidium excavatum | Terrestrial | Yellow | Bee | Euglossa purpurea | 9–12 (9–13) | - |
| Butterfly | Lienix nemesis | 14–18 (-) | - | |||
| Fly | Scaptia sp. | 9–13 (8–14) | 2% | |||
| Phragmipedium besseae | Terrestrial | Red | Hummingbird | Amazilia franciae | 5–7 (8–10) | - |
| Hummingbird | Lesbia nuna | 6–9 (7–10) | - | |||
| Bee | Euglossa purpurea | 7–11 (8–13) | - | |||
| Prosthechea fragrans | Epiphytic | Light green | Fly | Scaptia sp. | 5–9 (6–10) | 2% |
| Bee | Euglossa sp. | 11 (12–15) | - | |||
| Stelis tridentata | Epiphytic | Green | Bee | Euglossa sp. | 10–13 (11–14) | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reyes, H.C.; Draper, D.; Marques, I. Pollination in the Rainforest: Scarce Visitors and Low Effective Pollinators Limit the Fruiting Success of Tropical Orchids. Insects 2021, 12, 856. https://doi.org/10.3390/insects12100856
Reyes HC, Draper D, Marques I. Pollination in the Rainforest: Scarce Visitors and Low Effective Pollinators Limit the Fruiting Success of Tropical Orchids. Insects. 2021; 12(10):856. https://doi.org/10.3390/insects12100856
Chicago/Turabian StyleReyes, Hortensia Cabrera, David Draper, and Isabel Marques. 2021. "Pollination in the Rainforest: Scarce Visitors and Low Effective Pollinators Limit the Fruiting Success of Tropical Orchids" Insects 12, no. 10: 856. https://doi.org/10.3390/insects12100856
APA StyleReyes, H. C., Draper, D., & Marques, I. (2021). Pollination in the Rainforest: Scarce Visitors and Low Effective Pollinators Limit the Fruiting Success of Tropical Orchids. Insects, 12(10), 856. https://doi.org/10.3390/insects12100856