
Synaptic Interactions in Scorpion Peg Sensilla Appear to Maintain Chemosensory Neurons within Dynamic Firing Range


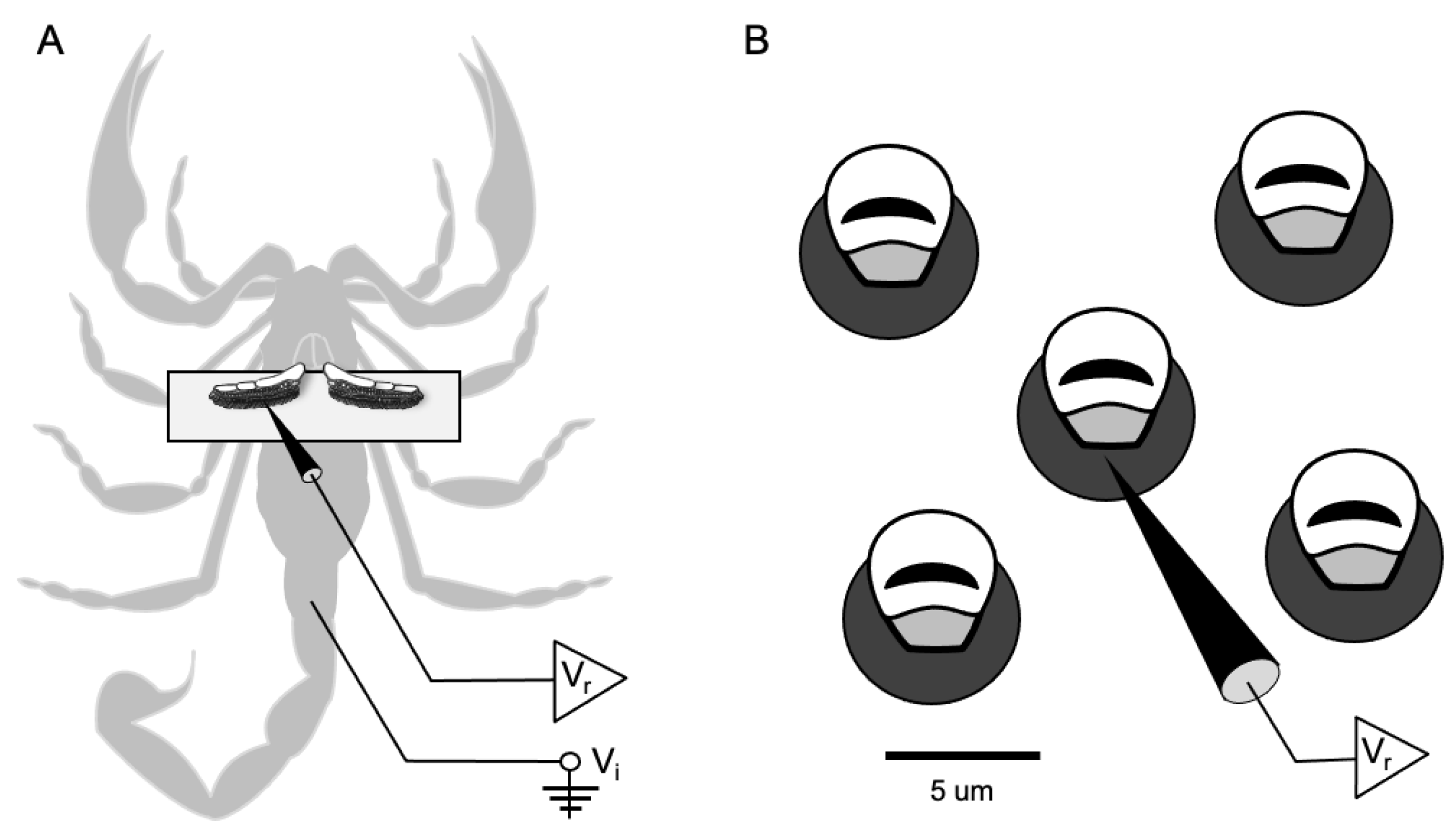{kind=link}
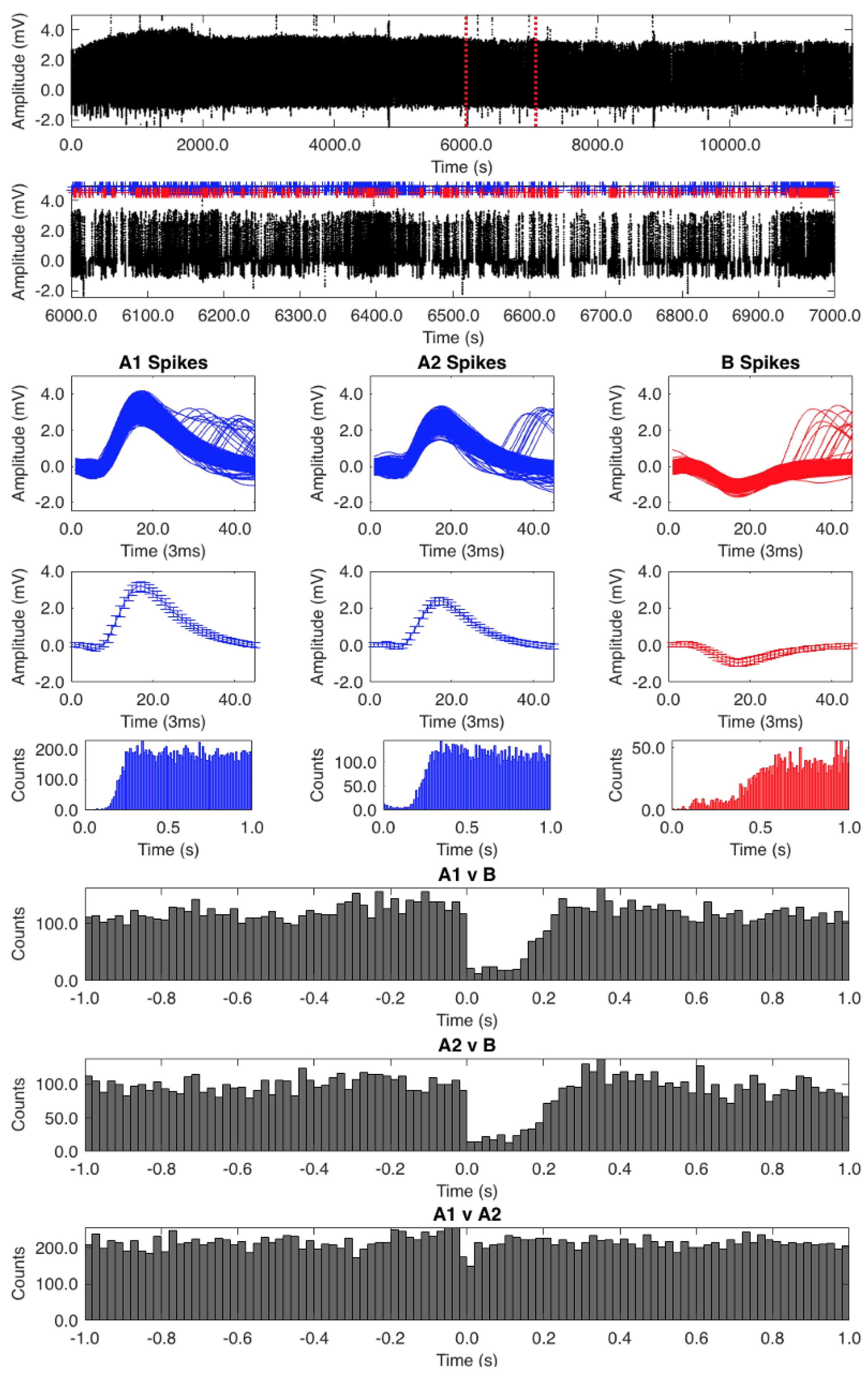{kind=link}
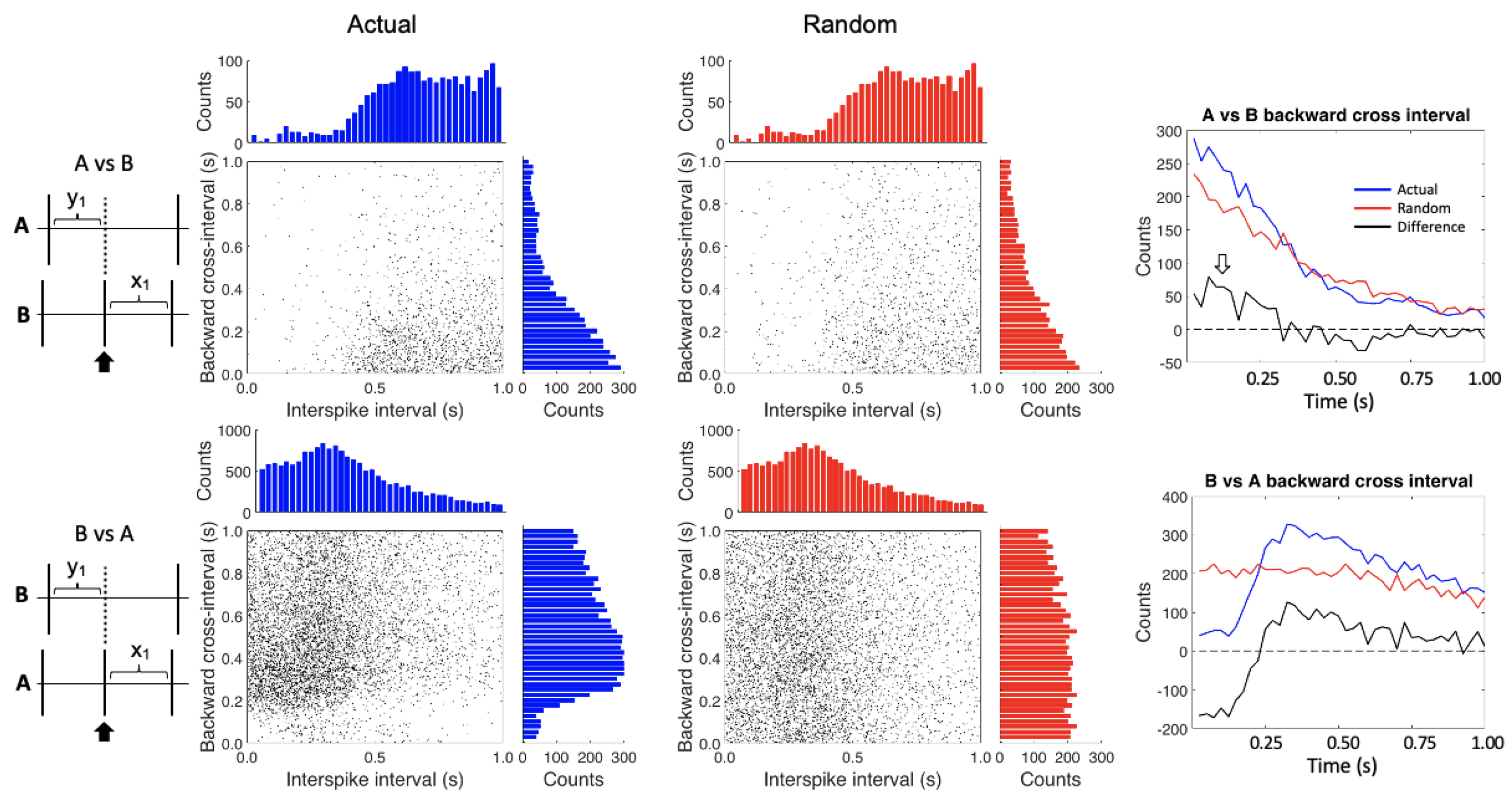{kind=link}
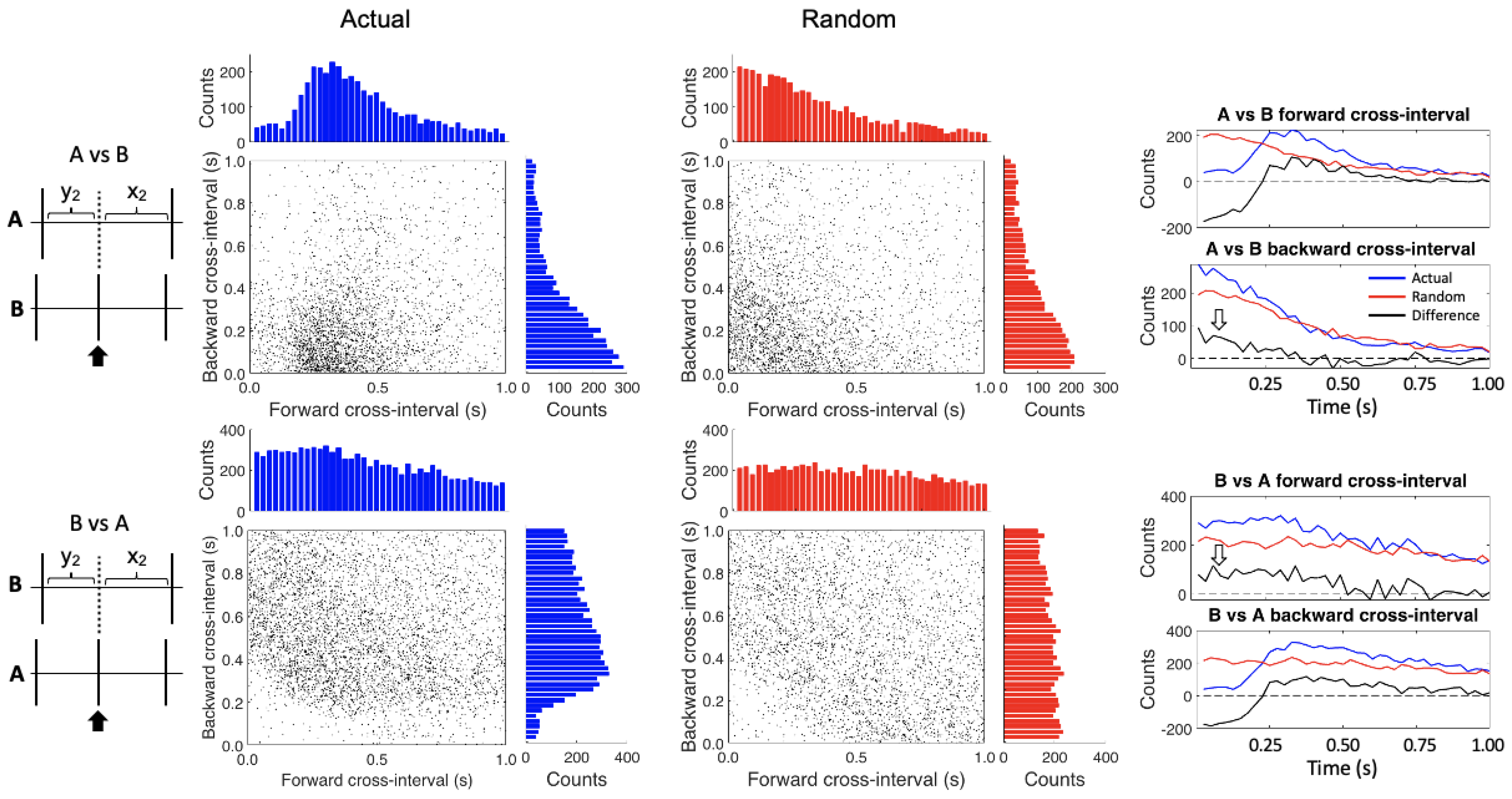{kind=link}
{kind=link}
{kind=link}
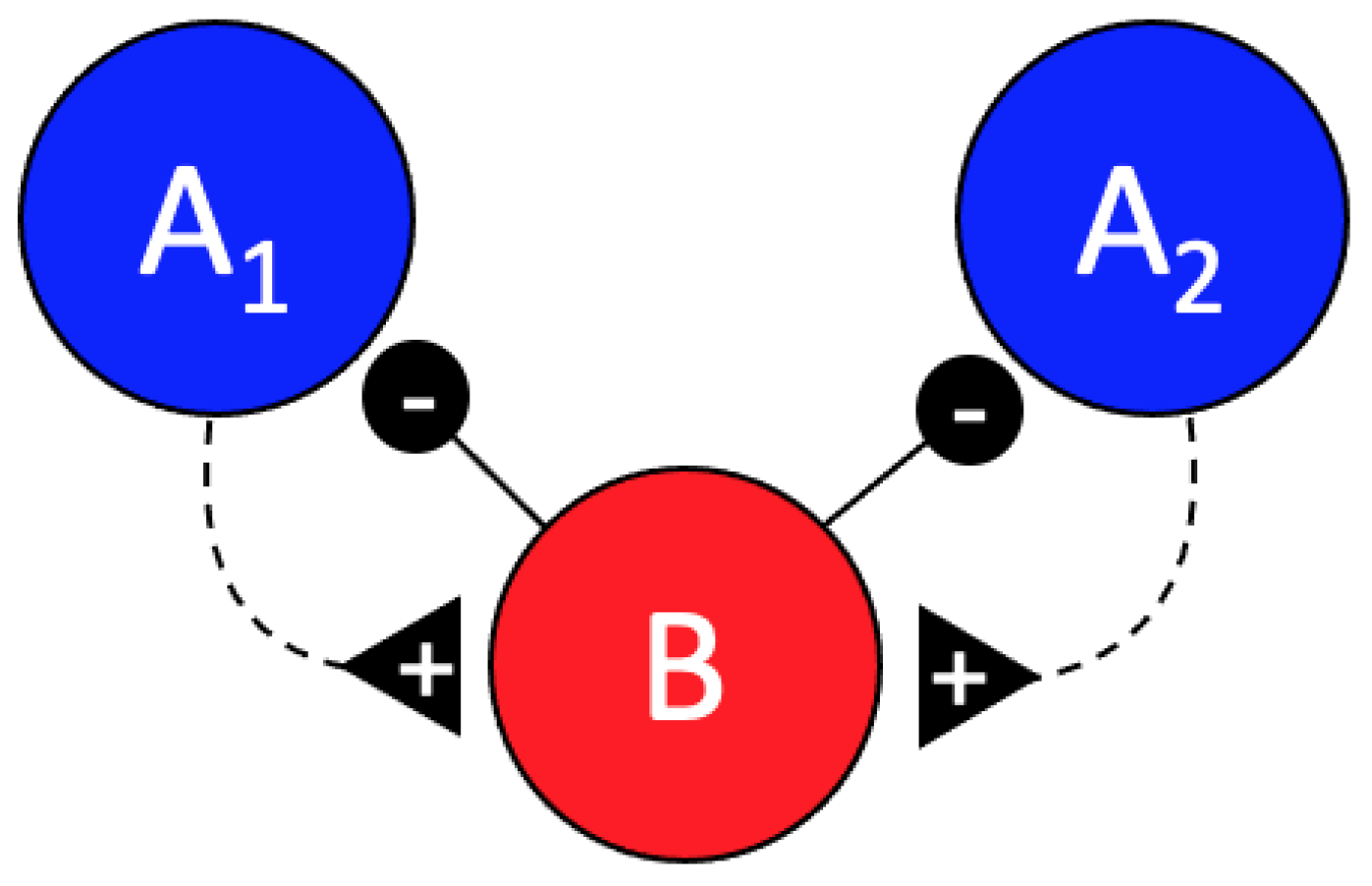
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Scorpions
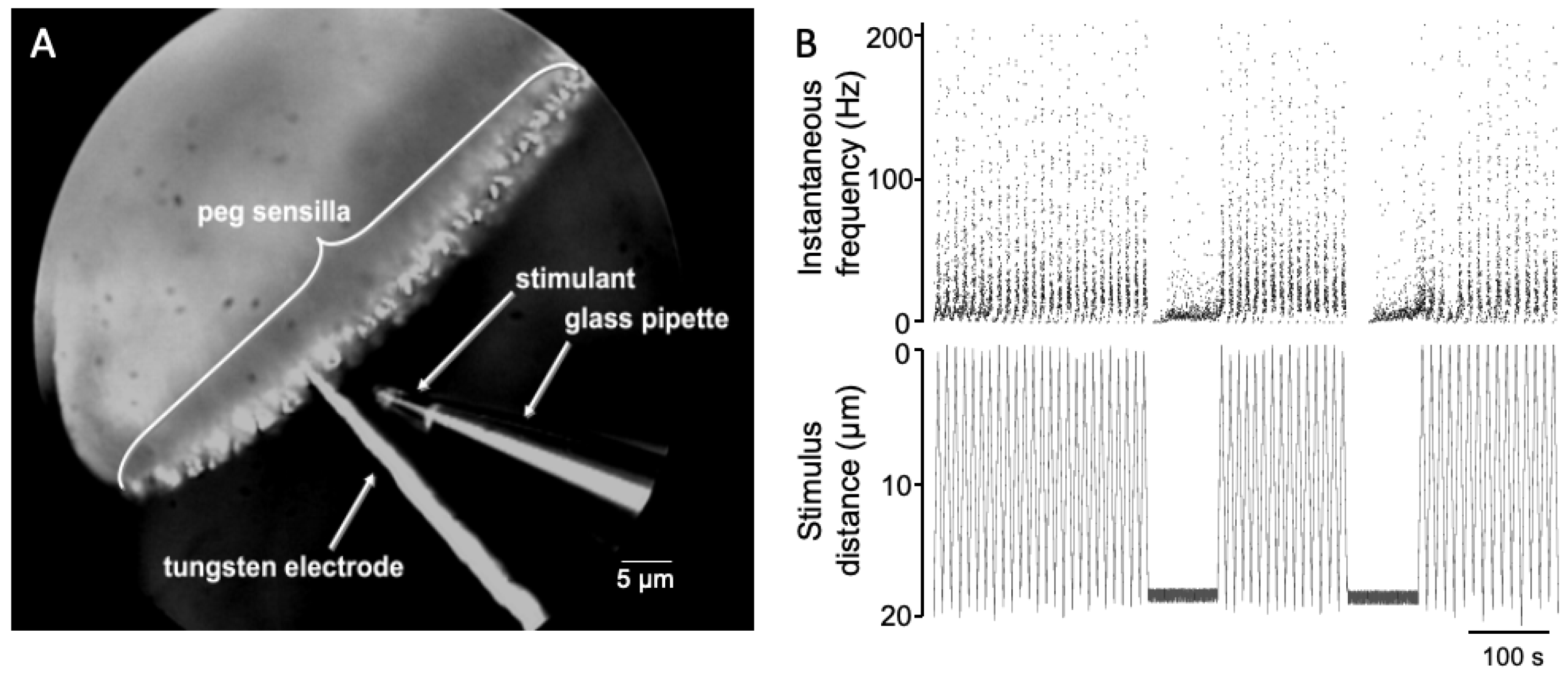
2.2. Preparation for Recording
2.3. Processing of Action Potentials
2.4. Auto- and Cross-Correlation Analysis
2.5. Conditional Interspike Interval and Conditional Cross-Interval Plots
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Gaffin, D.D.; Brownell, P.H. Response properties of chemosensory peg sensilla on the pectines of scorpions. J. Comp. Physiol. A 1997, 181, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Gaffin, D.D.; Brownell, P.H. Electrophysiological evidence of synaptic interactions within chemosensory sensilla of scorpion pectines. J. Comp. Physiol. A 1997, 181, 301–307. [Google Scholar] [CrossRef]
- Foelix, R.F.; Müller-Vorholt, G. The fine structure of scorpion sensory organs. II. Pecten sensilla. Bull. Br. Arachnol. Soc. 1983, 6, 68–74. [Google Scholar]
- Ivanov, V.; Balashov, Y. The structural and functional organization of the pectine in a scorpion Buthus eupeus Koch (Scorpiones, Buthidae) studied by electron microscopy. In The Fauna and Ecology of Arachnida; Trudy Zoological Institute: Leningrad, Russia, 1979; Volume 85, pp. 73–87. [Google Scholar]
- Knowlton, E.D.; Gaffin, D.D. Functionally redundant peg sensilla on the scorpion pecten. J. Comp. Physiol. A 2011, 197, 895. [Google Scholar] [CrossRef] [PubMed]
- Krapf, D. Contact chemoreception of prey in hunting scorpions (Arachnida: Scorpiones). Zool. Anz. 1986, 217, 119–129. [Google Scholar]
- Gaffin, D.D.; Brownell, P.H. Evidence of chemical signaling in the sand scorpion, Paruroctonus Mesaensis (Scorpionida: Vaejovidae). Ethology 1992, 91, 59–69. [Google Scholar] [CrossRef]
- Taylor, M.S.; Cosper, C.R.; Gaffin, D.D. Behavioral evidence of pheromonal signaling in desert grassland scorpions Paruroctonus Utahensis. J. Arachnol. 2012, 40, 240–244. [Google Scholar] [CrossRef]
- Gaffin, D.D.; Brayfield, B.P. Exploring the chemo-textural familiarity hypothesis for scorpion navigation. J. Arachnol. 2017, 45, 265–270. [Google Scholar] [CrossRef]
- Musaelian, A.; Gaffin, D.D. High-throughput simulations indicate feasibility of navigation by familiarity with a local sensor such as scorpion pectines. bioRxiv 2020. [Google Scholar] [CrossRef]
- Gaffin, D.D. Analysis of sensory processing in scorpion peg sensilla. J. Arachnol. 2010, 38, 1–8. [Google Scholar] [CrossRef]
- Gaffin, D.D. Electrophysiological analysis of synaptic interactions within peg sensilla of scorpion pectines. Microsc. Res. Tech. 2002, 58, 325–334. [Google Scholar] [CrossRef]
- Chapman, R.F.; Ascoli-Christensen, A.; White, P.R. Sensory coding for feeding deterrence in the grasshopper Schistocerca. Am. J. Exp. Biol. 1991, 158, 241–259. [Google Scholar] [CrossRef]
- Su, C.Y.; Menuz, K.; Reisert, J.; Carlson, J.R. Non-synaptic inhibition between grouped neurons in an olfactory circuit. Nature 2012, 492, 66–71. [Google Scholar] [CrossRef] [Green Version]
- Foelix, R.F.; Troyer, D. Giant neurons and associated synapses in the peripheral nervous system of whip spiders. J. Neurocytol. 1980, 9, 517–535. [Google Scholar] [CrossRef] [PubMed]
- Foelix, R.F. Occurrence of synapses in peripheral sensory nerves of arachnids. Nature 1975, 254, 146–148. [Google Scholar] [CrossRef] [PubMed]
- Foelix, R.; Troyer, D.; Igelmund, P. Peripheral synapses and giant neurons in whip spiders. Microsc. Res. Tech. 2002, 58, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Hartline, H.K.; Ratliff, F. Spatial summation of inhibitory influences in the eye of Limulus, Mutual Interact. Recept. Units. J. Gen. Physiol. 1958, 41, 1049–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuffler, S.W. Discharge patterns and functional organization of mammalian retina. J. Neurophysiol. 1953, 16, 37–68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Urban, N.N. Lateral inhibition in the olfactory bulb and in olfaction. Physiol. Behav. 2002, 77, 607–612. [Google Scholar] [CrossRef]
- Melville, J.M.; Tallarovic, S.K.; Brownell, P.H. Evidence of mate trailing in the giant hairy desert scorpion, Hadrurus Arizonensis (Scorpionida, Iuridae). J. Insect Behav. 2003, 16, 97–115. [Google Scholar] [CrossRef]
- Skutelsky, O. Flexibility in foraging tactics of Buthus Occ. Scorpions A Response Above-Ground Act. Termit. J. Arachnol. 1995, 23, 46–47. [Google Scholar]
- Tam, D.C.; Ebner, T.J.; Knox, C.K. Conditional cross-interval correlation analyses with applications to simultaneously recorded cerebellar Purkinje neurons. J. Neurosci. Methods 1988, 23, 23–33. [Google Scholar] [CrossRef]
- Knowlton, E.D.; Gaffin, D.D. Electrophysiology of scorpion peg sensilla. JoVE (J. Vis. Exp.) 2011, 50, e2642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knowlton, E.D.; Gaffin, D.D. A new approach to examining scorpion peg sensilla: The mineral oil flood technique. J. Arachnol. 2009, 37, 379–382. [Google Scholar] [CrossRef]
- Perkel, D.H.; Gerstein, G.L.; Moore, G.P. Neuronal spike trains and stochastic point processes: I. The single spike train. Biophys. J. 1967, 7, 391–418. [Google Scholar] [CrossRef] [Green Version]
- Perkel, D.H.; Gerstein, G.L.; Smith, M.S.; Tatton, W.G. Nerve-impulse patterns: A quantitative display technique for three neurons. Brain Res. 1975, 100, 271–296. [Google Scholar] [CrossRef]
- Perkel, D.H.; Gerstein, G.L.; Moore, G.P. Neuronal spike trains and stochastic point processes: II. Simultaneous spike trains. Biophys. J. 1967, 7, 419–440. [Google Scholar] [CrossRef] [Green Version]
- Brownell, P.H. Glomerular cytoarchitectures in chemosensory cystems of arachnids. Ann. N. Y. Acad. Sci. 1998, 855, 502–507. [Google Scholar] [CrossRef]
- Brownell, P. Sensory ecology and orientational behaviors. In Scorpion Biology and Research; Oxford University Press: Oxford, UK, 2001; pp. 159–183. [Google Scholar]
- Drozd, D.; Wolf, H.; Stemme, T. Structure of the pecten neuropil pathway and its innervation by bimodal peg afferents in two scorpion species. PLoS ONE 2020, 15, e0243753. [Google Scholar] [CrossRef]
- Hughes, K.L.; Gaffin, D.D. Investigating sensory processing in the pectines of the striped bark scorpion, Centruroides vittatus. Invertebr. Neurosci. 2019, 19, 9. [Google Scholar] [CrossRef] [PubMed]
- Knowlton, E.D.; Gaffin, D.D. A new tip-recording method to test scorpion pecten chemoresponses to water-soluble stimulants. J. Neurosci. Methods 2010, 193, 264–270. [Google Scholar] [CrossRef]
- Whitmire, C.; Stanley, G. Rapid sensory adaptation redux: A circuit perspective. Neuron 2016, 92, 298–315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miriyala, A.; Kessler, S.; Rind, F.C.; Wright, G.A. Burst firing in fee gustatory neurons prevents adaptation. Curr. Biol. 2018, 28, 1585–1594.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaffin, D.D.; Walvoord, M.E. Scorpion peg sensilla: Are they the same or are they different? Euscorpius 2004, 17, 7–15. [Google Scholar]
- Cohen, R.A. Lateral inhibition. In Encyclopedia of Clinical Neuropsychology; Kreutzer, J.S., DeLuca, J., Caplan, B., Eds.; Springer: New York, NY, USA, 2011; pp. 1436–1437. [Google Scholar] [CrossRef]
- Ratliff, F.; Knight, B.W.; Milkman, N. Superposition of excitatory and inhibitory influences in the retina of Limulus: Eff. Delayed Inhib. Proc. Natl. Acad. Sci. USA 1970, 67, 1558–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pruszynski, J.A.; Johansson, R.S. Edge-orientation processing in first-order tactile neurons. Nat. Neurosci. 2014, 17, 1404–1409. [Google Scholar] [CrossRef] [PubMed]
- Melville, J.M. The Pectines of Scorpions: Analysis of Structure and Function. Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, 2000. [Google Scholar]
- Prévost, E.D.; Stemme, T. Non-visual homing and the current status of navigation in scorpions. Anim. Cogn. 2020, 23, 1215–1234. [Google Scholar] [CrossRef]
- Gaffin, D.D.; Curry, C.M. Arachnid navigation—A review of classic and emerging models. J. Arachnol. 2020, 48, 1–25. [Google Scholar] [CrossRef]
- Hebets, E.A.; Chapman, R.F. Electrophysiological studies of olfaction in the whip spider Phrynus parvulus (Arachnida, Amblypygi). J. Insect Physiol. 2000, 46, 1441–1448. [Google Scholar] [CrossRef] [Green Version]
- Hebets, E.A.; Aceves-Aparicio, A.; Aguilar-Argüello, S.; Bingman, V.P.; Escalante, I.; Gering, E.J.; Nelsen, D.R.; Rivera, J.; Sánchez-Ruiz, J.; Segura-Hernández, L.; et al. Multimodal sensory reliance in the nocturnal homing of the amblypygid Phrynus pseudoparvulus (Class Arachnida, Order Amblypygi)? Behav. Process. 2014, 108, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Hebets, E.A.; Gering, E.J.; Bingman, V.P.; Wiegmann, D.D. Nocturnal homing in the tropical amblypygid Phrynus pseudoparvulus (Class Arachnida, Order Amblypygi). Anim. Cogn. 2014, 17, 1013–1018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebets, E.A. Relating the unique sensory system of amblypygids to the ecology and behavior of Phrynus parvulus Costa Rica (Arachnida, Amblypygi). Can. J. Zool. Ott. 2002, 80, 286–295. [Google Scholar] [CrossRef]
- Bingman, V.P.; Graving, J.M.; Hebets, E.A.; Wiegmann, D.D. Importance of the antenniform legs, but not vision, for homing by the neotropical whip spider Paraphrynus laevifrons. J. Exp. Biol. 2017, 220, 885–890. [Google Scholar] [CrossRef] [Green Version]
- Foelix, R.; Hebets, E. Sensory biology of whip spiders (Arachnida, Amblypygi). Andrias 2001, 15, 129–140. [Google Scholar]
- Fabian-Fine, R.; Meinertzhagen, I.A.; Seyfarth, E.A. Organization of efferent peripheral synapses at mechanosensory neurons in spiders. J. Comp. Neurol. 2000, 420, 195–210. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gaffin, D.D.; Shakir, S.F. Synaptic Interactions in Scorpion Peg Sensilla Appear to Maintain Chemosensory Neurons within Dynamic Firing Range. Insects 2021, 12, 904. https://doi.org/10.3390/insects12100904
Gaffin DD, Shakir SF. Synaptic Interactions in Scorpion Peg Sensilla Appear to Maintain Chemosensory Neurons within Dynamic Firing Range. Insects. 2021; 12(10):904. https://doi.org/10.3390/insects12100904
Chicago/Turabian StyleGaffin, Douglas D., and Safra F. Shakir. 2021. "Synaptic Interactions in Scorpion Peg Sensilla Appear to Maintain Chemosensory Neurons within Dynamic Firing Range" Insects 12, no. 10: 904. https://doi.org/10.3390/insects12100904
APA StyleGaffin, D. D., & Shakir, S. F. (2021). Synaptic Interactions in Scorpion Peg Sensilla Appear to Maintain Chemosensory Neurons within Dynamic Firing Range. Insects, 12(10), 904. https://doi.org/10.3390/insects12100904