
The Effects of Nosema apis and Nosema ceranae Infection on Survival and Phenoloxidase Gene Expression in Galleria mellonella (Lepidoptera: Galleriidae) Compared to Apis mellifera


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Selection of Insects
2.2. Preparation of Nosema Stock Solutions
2.3. Experimental Infection of G. mellonella and A. mellifera with Nosema spp.
2.4. Determination of the mRNA Level of the Phenoloxidase (PO) Gene
2.5. Statistical Analysis
3. Results
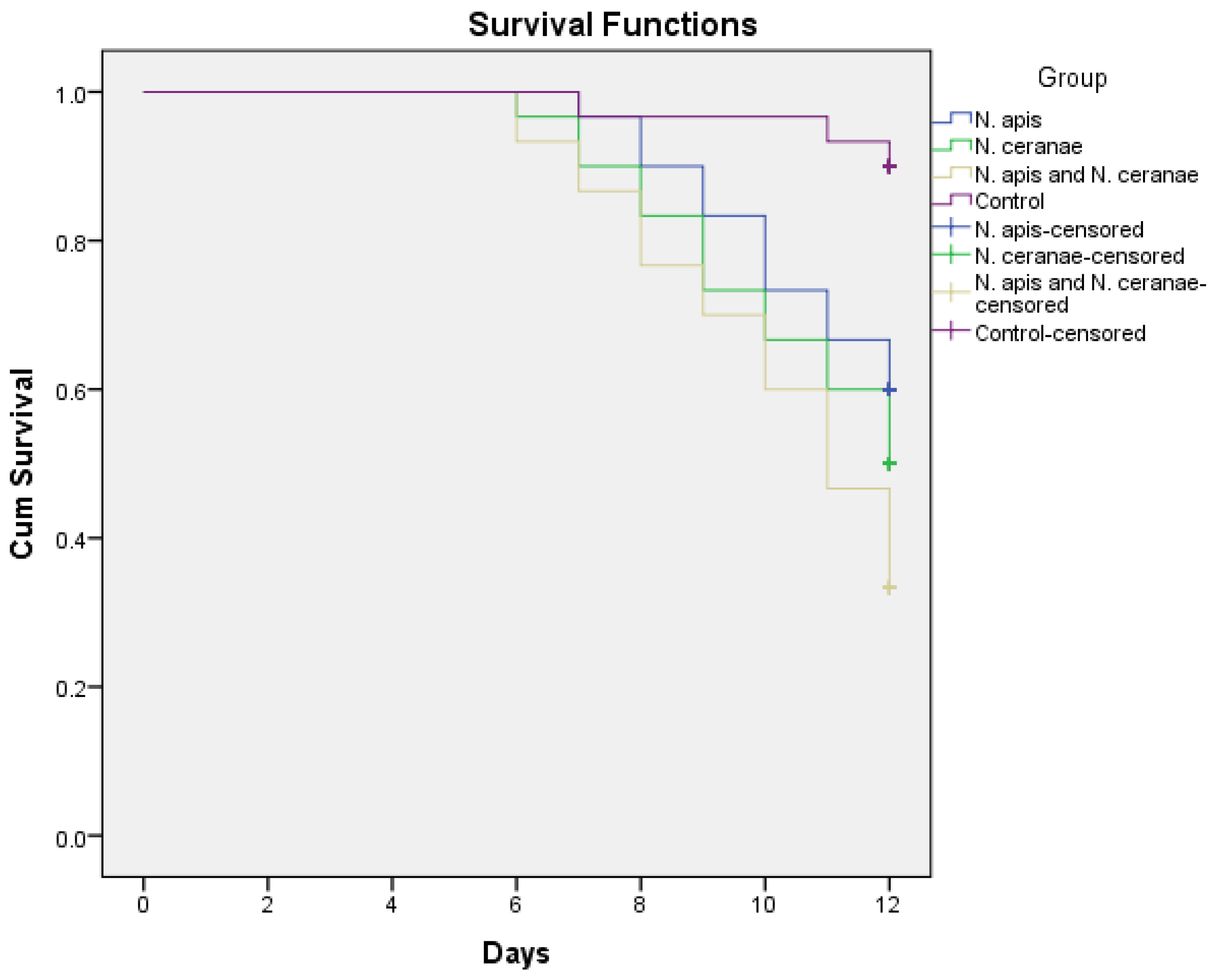
3.1. Effect of Nosema Infections on Moth and Bee Survival
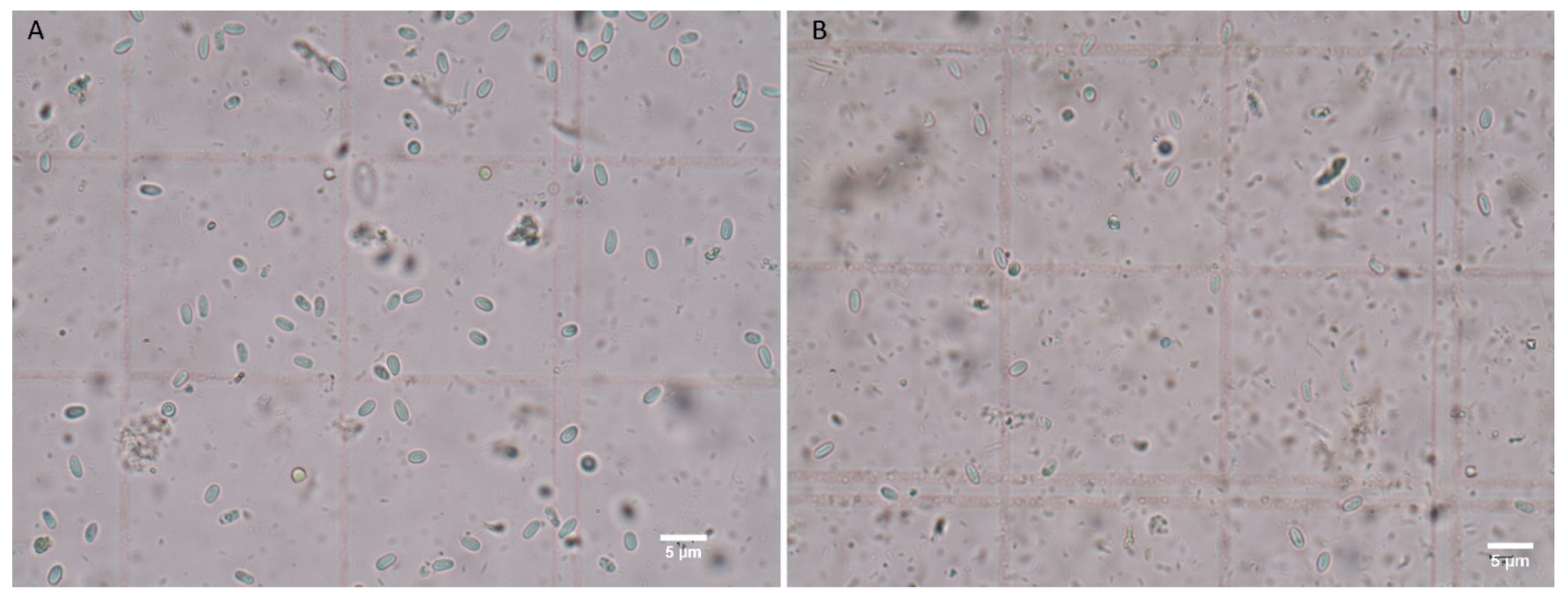
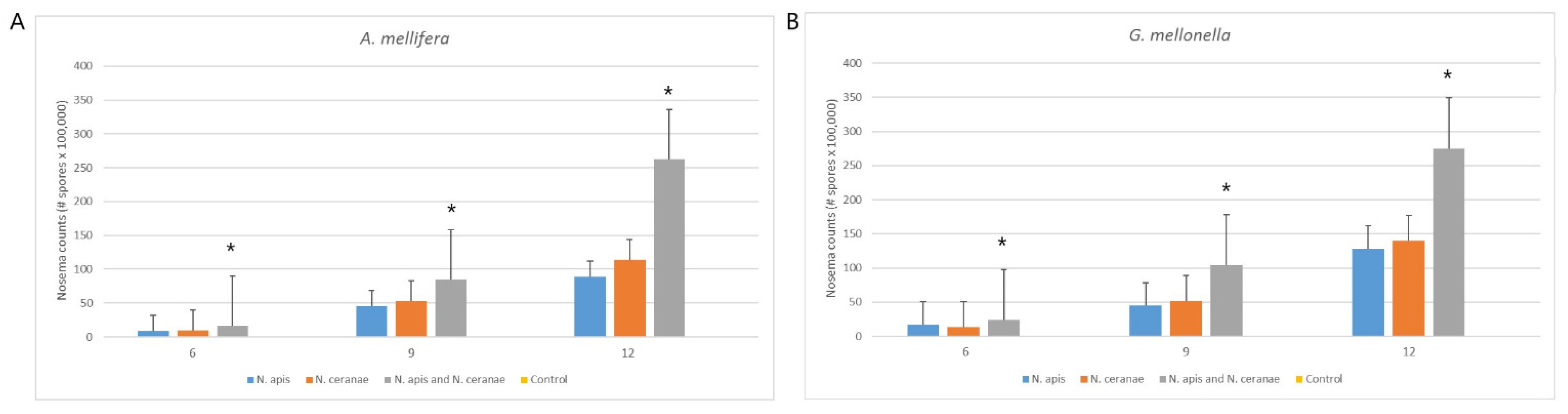
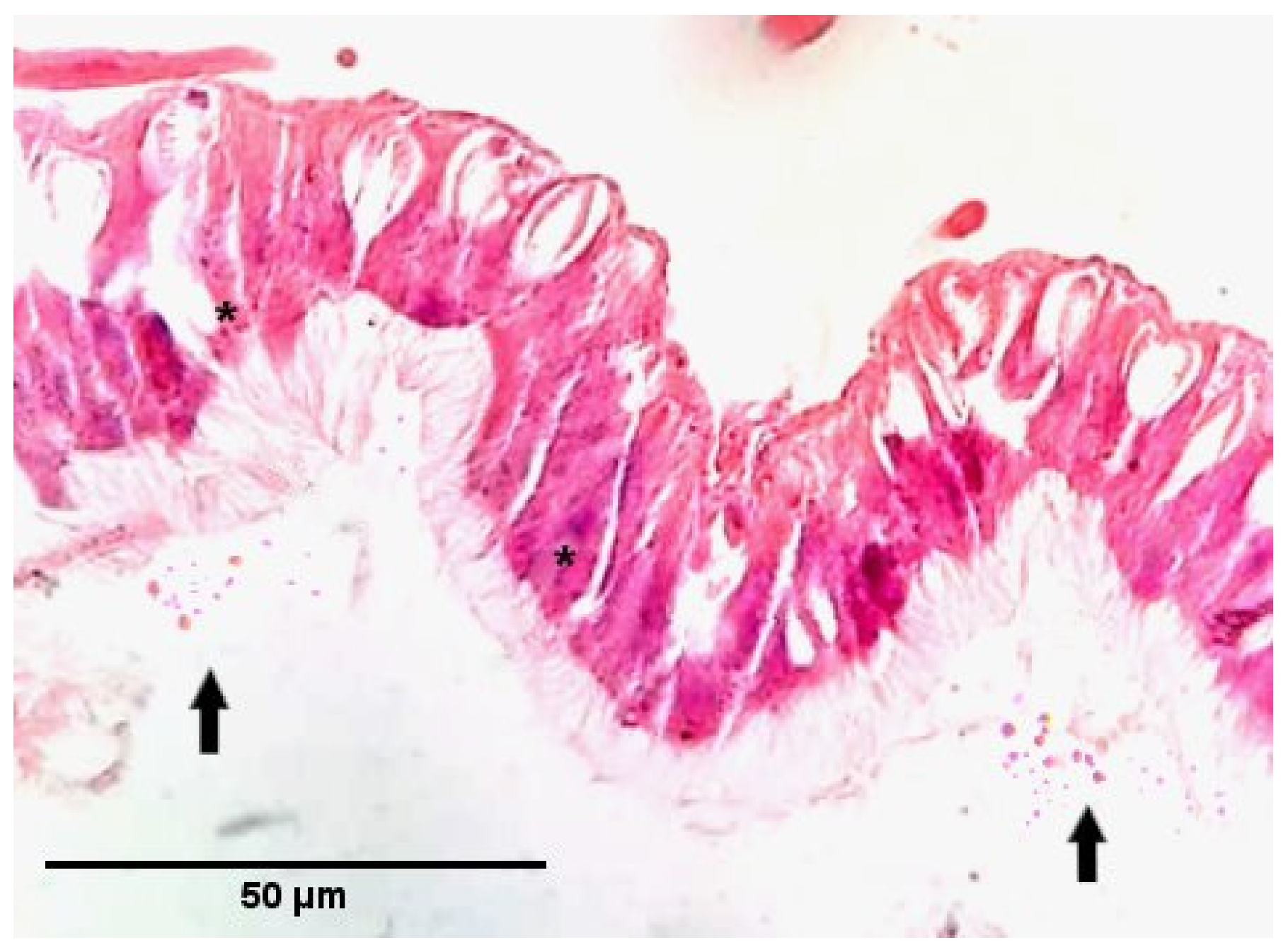
3.2. Infection Capability of Nosema Species to Moth and Bee
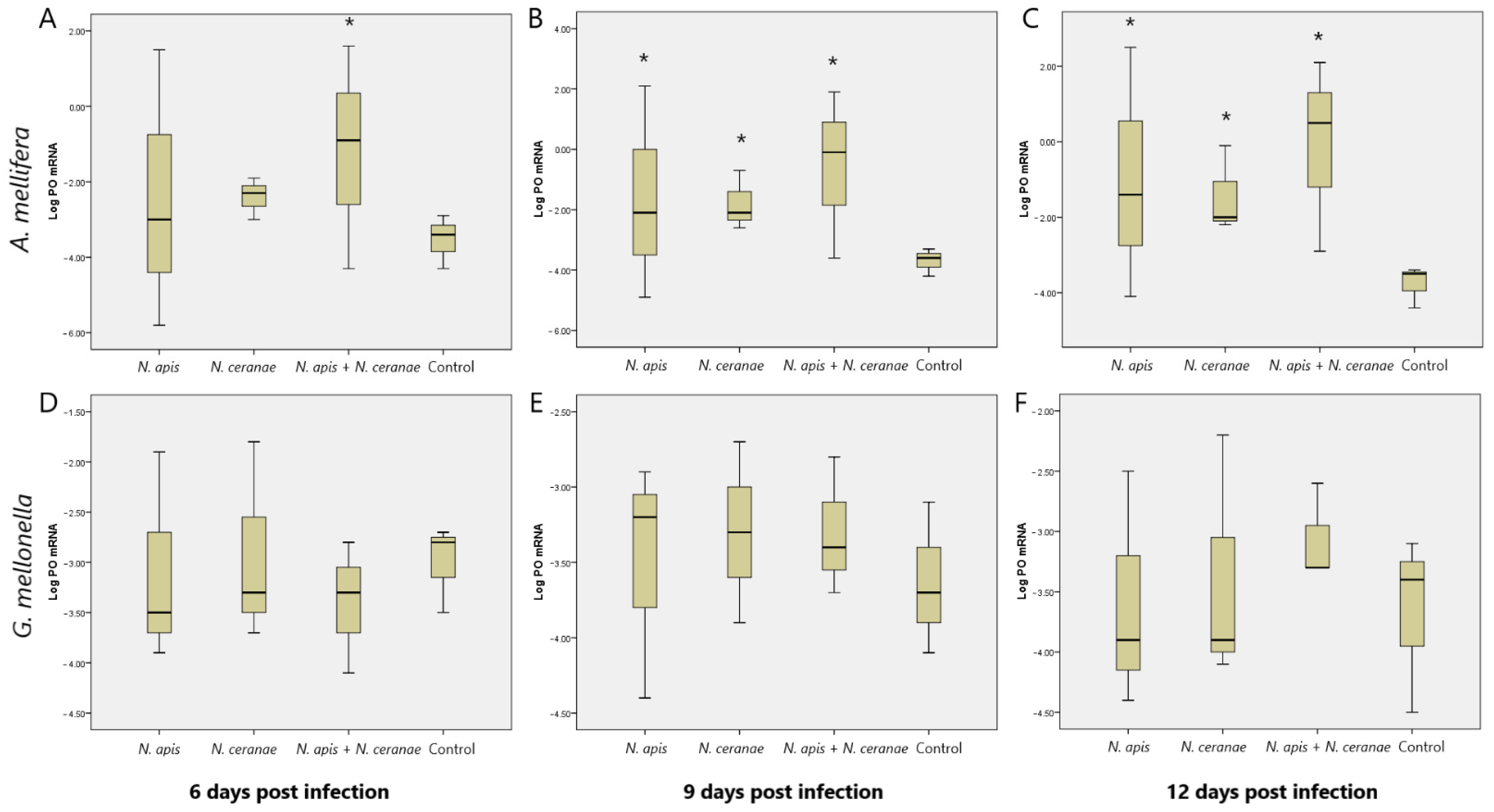
3.3. Effect of Nosema Infections on the Level of Expression of the PO-related Gene in Moth and Bee
4. Discussion
5. Conclusions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ellis, J.D.; Graham, J.R.; Mortensen, A. Standard methods for wax moth research. J. Apic. Res. 2013, 52, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.J.-Y.; Loh, J.M.S.; Proft, T. Galleria mellonella infection models for the study of bacterial diseases and for antimicrobial drug testing. Virulence 2016, 7, 214–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forsgren, E.; Fries, I. Comparative virulence of Nosema ceranae and Nosema apis in individual European honey bees. Vet. Parasitol. 2010, 170, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Lipa, J.J. Microsporidian infections of Galleria mellonella (L.) (Lepidoptera, Galleriidae) with the description of a new species Nosema galleriae n.sp. Acta Protozool. 1977, 16, 141–150. [Google Scholar]
- Kučera, M.; Weiser, J. Lactate dehydrogenase isoenzymes in the larvae of Barathra brassicae and Galleria mellonella during microsporidian infection. J. Invertebr. Pathol. 1975, 25, 109–114. [Google Scholar] [CrossRef]
- Kučera, M.; Weiser, J. Different course of proteolytic inhibitory activity and proteolytic activity in Galleria mellonella larvae infected by Nosema algerae and Vairimorpha heterosporum. J. Invertebr. Pathol. 1985, 45, 41–46. [Google Scholar] [CrossRef]
- Decker, H.; Jaenicke, E. Recent findings on Phenoloxidase activity and antimicrobial activity of hemocyanins. Dev. Comp. Immunol. 2004, 28, 673–687. [Google Scholar] [CrossRef]
- Charnley, K. Physiological aspects of destructive pathogenesis in insects by fungi: A speculative review. In Invertebrate-Microbial Interactions, 2nd ed.; Anderson, J.M., Raymer, A.D.M., Walton, D.W.H., Eds.; Cambridge University Press: London, UK, 1984; pp. 229–270. [Google Scholar]
- González-Santoyo, I.; Córdoba-Aguilar, A. Phenoloxidase: A key component of the insect immune system. Entomol. Exp. Appl. 2012, 142, 1–16. [Google Scholar] [CrossRef]
- Zdybicka-Barabas, A.; Mak, P.; Jakubowicz, T.; Cytryńska, M. Lysozyme and defense peptides as suppressors of phenoloxidase activity in Galleria mellonella. Arch. Insect Biochem. 2014, 87, 1–12. [Google Scholar] [CrossRef]
- Brodschneider, R.; Moosbeckhofer, R.; Crailsheim, K. Surveys as a tool to record winter losses of honey bee colonies: A 2-year case study in Austria and South Tyrol, Tyrol. J. Apic. Res. 2010, 49, 23–30. [Google Scholar] [CrossRef]
- Neumann, P.; Carreck, N.L. Honey bee colony losses. J. Apic. Res. 2010, 49, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Van der Zee, R.; Pisa, L.; Andonov, S.; Brodschneider, R.; Charrière, J.-D.; Chlebo, R.; Coffey, M.F.; Crailsheim, K.; Dahle, D.; Gajda, A.; et al. Managed honey bee colony losses in Canada, China, Europe, Israel and Turkey, for the winters of 2008-9 and 2009-10. J. Apic. Res. 2012, 51, 100–114. [Google Scholar] [CrossRef]
- Paxton, R.J.; Klee, J.; Korpela, S.; Fries, I. Nosema ceranae has infected Apis mellifera in Europe since at least 1998 and may be more virulent than Nosema apis. Apidologie 2007, 38, 558–565. [Google Scholar] [CrossRef]
- Özgör, E.; Celebier, I.; Ulusoy, M.; Keskin, N. First detection of Nosema ceranae and Nosema apis in greater wax moth Galleria mellonella. J. Apic. Sci. 2017, 61, 185–192. [Google Scholar]
- Cantwell, G.E. Standard methods for counting Nosema spores. Am. Bee J. 1970, 110, 222–223. [Google Scholar]
- Bourgeois, A.L.; Rinderer, T.E.; Beaman, L.D.; Danka, R.G. Genetic detection and quantification Nosema apis and N. ceranae in the honey bee. J. Invertebr. Pathol. 2010, 103, 53–58. [Google Scholar] [CrossRef]
- Chertkova, E.A.; Grizanova, E.V.; Dubovskiy, E.V. Bacterial and fungal infections induce bursts of dopamine in the haemolymph of the Colorado potato beetle Leptinotarsa decemlineata and greater wax moth Galleria mellonella. J. Invertebr. Pathol. 2018, 153, 203–206. [Google Scholar] [CrossRef]
- Higes, M.; García-Palencia, P.; Martín-Hernández, R.; Meana, A. Experimental infection of Apis mellifera honeybees with Nosema ceranae (Microsporidia). J. Invertebr. Pathol. 2007, 94, 211–217. [Google Scholar] [CrossRef]
- Emery, H.; Johnston, R.; Rowley, A.F.; Coates, C.J. Indomethacin-induced gut damage in a surrogate insect model, Galleria mellonella. Arch. Toxicol. 2019, 93, 2347–2360. [Google Scholar] [CrossRef] [Green Version]
- Goblirsch, M. Nosema ceranae disease of the honey bee (Apis mellifera). Apidologie 2018, 49, 131–150. [Google Scholar] [CrossRef] [Green Version]
- Antúnez, K.; Martin-Hernandez, R.; Prieto, L.; Meana, A.; Zunino, P.; Higes, M. Immune suppression in the honey bee (Apis mellifera) following infection by Nosema ceranae (Microsporidia). Environ. Microbiol. 2009, 11, 2284–2290. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Scherfer, C.; Korayem, A.M.; Zhao, Z.; Schmidt, O.; Theopold, U. Insect hemolymph clotting: Evidence for interaction between the coagulation system and the prophenoloxidase activating cascade. Insect Biochem. Mol. Biol. 2002, 32, 919–928. [Google Scholar] [CrossRef]
- Yang, X.; Cox-Foster, D.L. Impact of an ectoparasite on the immunity and pathology of an invertebrate: Evidence for host immunosuppression and viral amplification. Proc. Natl. Acad. Sci. USA 2005, 21, 7470–7475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mowlds, P.; Barron, A.; Kavanagh, K. Physical stress primes the immune response of Galleria mellonella larvae to infection by Candida albicans. Microb. Infect. 2008, 10, 628–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, L.M.; Becnel, J.J. Microsporidia: Pathogens of Opportunity, 1st ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2014. [Google Scholar] [CrossRef]
- Klee, J.; Besana, A.M.; Genersch, E.; Gisder, S.; Nanetti, A.; Tam, D.Q.; Chinh, T.X.; Puerta, F.; Ruzi, J.M.; Kryger, P.; et al. Widespread dispersal of the microsporidian Nosema ceranae, an emergent pathogen of the western honey bee, Apis mellifera. J. Invertebr. Pathol. 2007, 96, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tokarev, Y.S.; Grizanova, E.V.; Ignatieva, A.N.; Dubovskiy, I.M. Greater wax moth Galleria mellonella (Lepidoptera: Pyralidae) as a resistant model host for Nosema pyrausta (Microsporidia: Nosematidae). J. Invertebr. Pathol. 2018, 157, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Martín-Hernández, R.; Botías, C.; Barrios, L.; Martínez-Salvador, A.; Meana, A.; Mayack, C.; Higes, M. Comparison of the energetic stress associated with experimental Nosema ceranae and Nosema apis infection of honeybees (Apis mellifera). Parasitol. Res. 2011, 109, 605–612. [Google Scholar] [CrossRef]
- Williams, G.R.; Shutler, D.; Burgher-MacLellan, K.L.; Rogers, R.E.L. Infra-population and -community dynamics of the parasites Nosema apis and Nosema ceranae, and consequences for honey bee (Apis mellifera) hosts. PLoS ONE 2014, 9, e99456. [Google Scholar] [CrossRef] [Green Version]
- Fries, I. Nosema ceranae in European honey bees (Apis mellifera). J. Invertebr. Pathol. 2010, 103, 73–79. [Google Scholar] [CrossRef]
- Kurtti, T.J.; Ross, S.E.; Liu, Y.; Munderloh, U.G. In vitro developmental biology and spore production in Nosema furnacalis (Microspora: Nosematidae). J. Invertebr. Pathol. 1994, 63, 188–196. [Google Scholar] [CrossRef]
- Ishihara, R. The life cycle of Nosema bombycis as revealed in tissue culture cells of Bombyx mori. J. Invertebr. Pathol. 1969, 14, 316–320. [Google Scholar] [CrossRef]
- Malysh, J.M.; Ignatieva, A.N.; Artokhin, K.S.; Frolov, A.N.; Tokarev, Y.S. Natural infection of the beet webworm Loxostege sticticalis L. (Lepidoptera: Crambidae) with three Microsporidia and host switching in Nosema ceranae. Parasitol. Res. 2018, 117, 3039–3044. [Google Scholar] [CrossRef] [PubMed]
- Preston, C.E.; Agnello, A.M.; Hajek, A.E. Nosema maddoxi (Microsporidia: Nosematidae) in brown marmorated stink bug, Halyomorpha halys (Hemiptera: Pentatomidae), populations in the United States. Biol. Control 2020, 144, 104213. [Google Scholar] [CrossRef]
- Milbrath, M.O.; van Tran, T.; Huang, W.-F.; Solter, L.F.; Tarpy, D.R.; Lawrence, F.; Huang, Z.Y. Comparative virulence and competition between Nosema apis and Nosema ceranae in honey bees (Apis mellifera). J. Invertebr. Pathol. 2015, 125, 9–15. [Google Scholar] [CrossRef]
- Gisder, S.; Horchler, L.; Pieper, F.; Schüler, V.; Šima, P.; Genersch, E. Rapid gastrointestinal passage may protect Bombus terrestris from becoming a true host for Nosema ceranae. Appl. Environ. Microbiol. 2020, 86, e00629-20. [Google Scholar] [CrossRef]
- Wilson-Rich, N.; Spivak, M.; Fefferman, N.H.; Starks, P.T. Genetic, individual, and group facilitation of disease resistance in insect societies. Annu. Rev. Entomol. 2009, 54, 405–423. [Google Scholar] [CrossRef] [Green Version]
- Alaux, C.; Brunet, J.-L.; Dussaubat, C.; Mondet, F.; Tchamitchan, S.; Cousin, M.; Brillard, J.; Baldy, A.; Belzunces, L.P.; Conte, Y.L. Interactions between Nosema microspores and a neonicotinoid weaken honeybees (Apis mellifera). Environ. Microbiol. 2010, 12, 774–782. [Google Scholar] [CrossRef] [Green Version]
- Dunphy, G.; Halwani, A. Haemolymph proteins of larvae of Galleria mellonella detoxify endotoxins of the insect pathogenic bacteria Xenorhabdus nematophilus (Enterobacteriaceae). J. Insect Physiol. 1997, 43, 1023–1029. [Google Scholar] [CrossRef]
- Zdybicka-Barabas, A.; Cytryńska, M. Phenoloxidase activity in hemolymph of Galleria mellonella larvae challenged with Aspergillus oryzae. Ann. UMCS Biol. 2010, 65, 49–57. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Özgör, E. The Effects of Nosema apis and Nosema ceranae Infection on Survival and Phenoloxidase Gene Expression in Galleria mellonella (Lepidoptera: Galleriidae) Compared to Apis mellifera. Insects 2021, 12, 953. https://doi.org/10.3390/insects12100953
Özgör E. The Effects of Nosema apis and Nosema ceranae Infection on Survival and Phenoloxidase Gene Expression in Galleria mellonella (Lepidoptera: Galleriidae) Compared to Apis mellifera. Insects. 2021; 12(10):953. https://doi.org/10.3390/insects12100953
Chicago/Turabian StyleÖzgör, Erkay. 2021. "The Effects of Nosema apis and Nosema ceranae Infection on Survival and Phenoloxidase Gene Expression in Galleria mellonella (Lepidoptera: Galleriidae) Compared to Apis mellifera" Insects 12, no. 10: 953. https://doi.org/10.3390/insects12100953
APA StyleÖzgör, E. (2021). The Effects of Nosema apis and Nosema ceranae Infection on Survival and Phenoloxidase Gene Expression in Galleria mellonella (Lepidoptera: Galleriidae) Compared to Apis mellifera. Insects, 12(10), 953. https://doi.org/10.3390/insects12100953