
Exploring the Use of Entomopathogenic Nematodes and the Natural Products Derived from Their Symbiotic Bacteria to Control the Grapevine Moth, Lobesia botrana (Lepidoptera: Tortricidae)

 ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Nematode Rearing
2.2. Symbiotic Bacterial Isolation and Natural Products Generation
2.3. Larvicidal and Pupicidal Assays
2.4. Statistical Analysis
3. Results
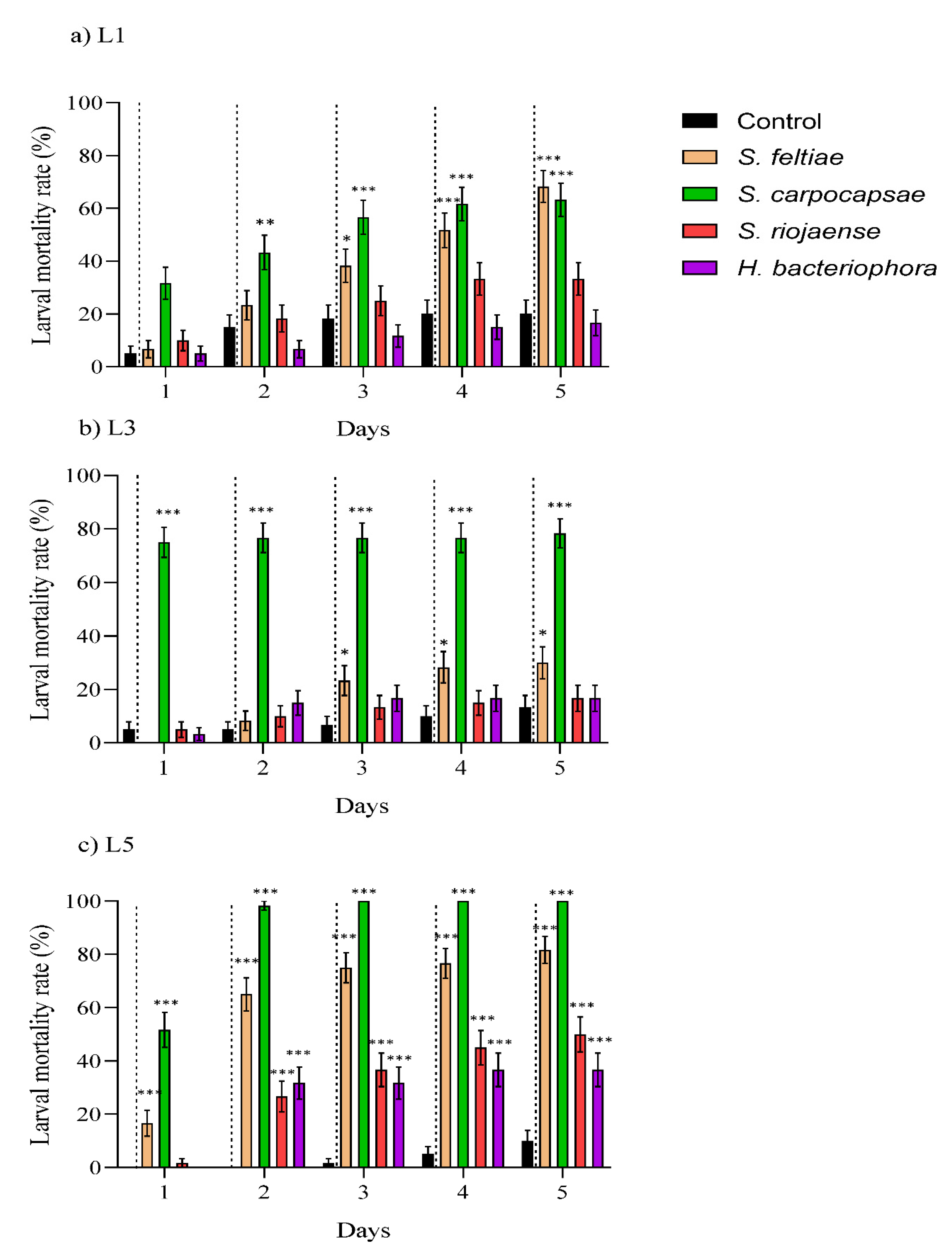
3.1. Larvicidal Effect by Entomopathogenic Nematodes
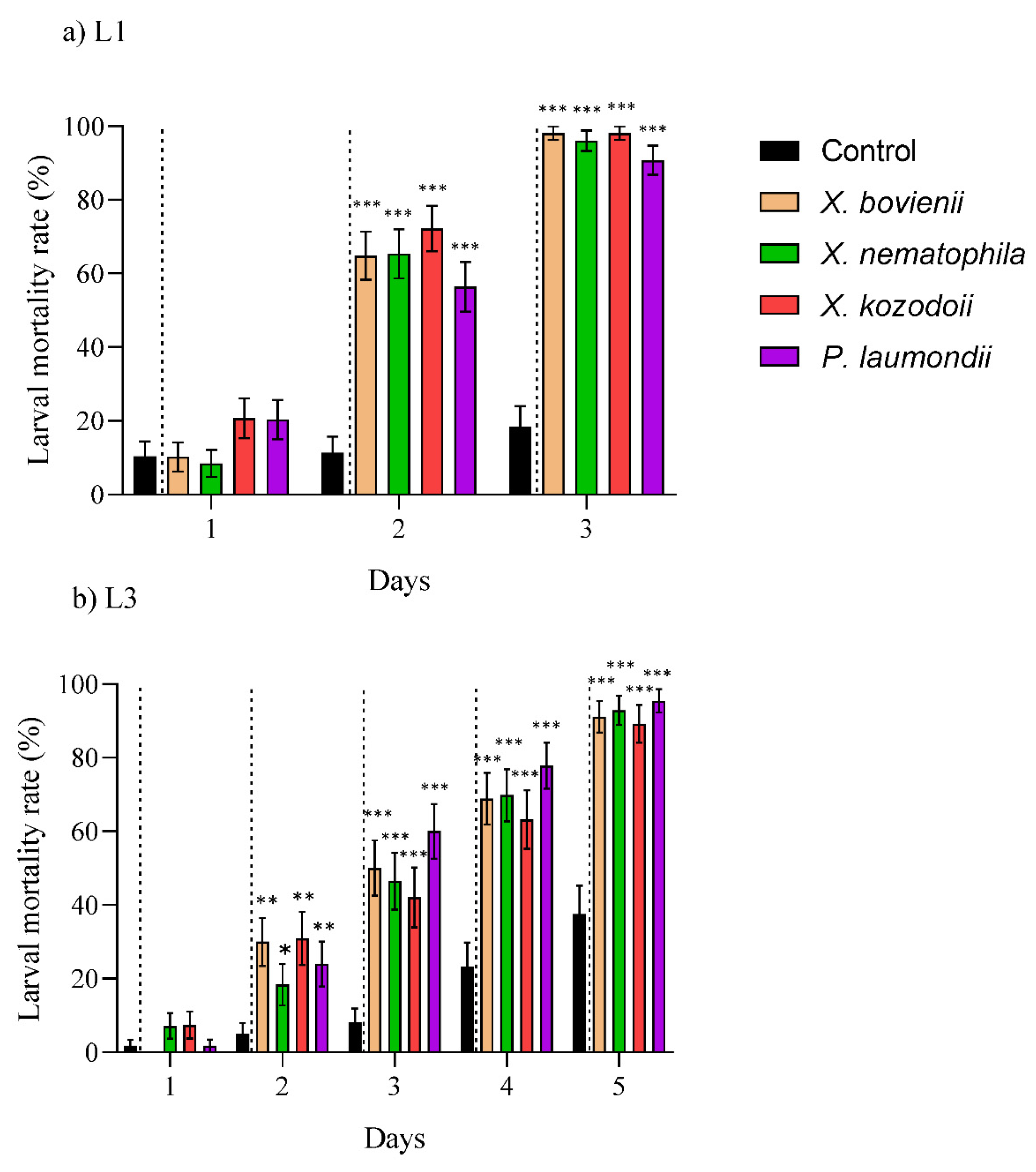
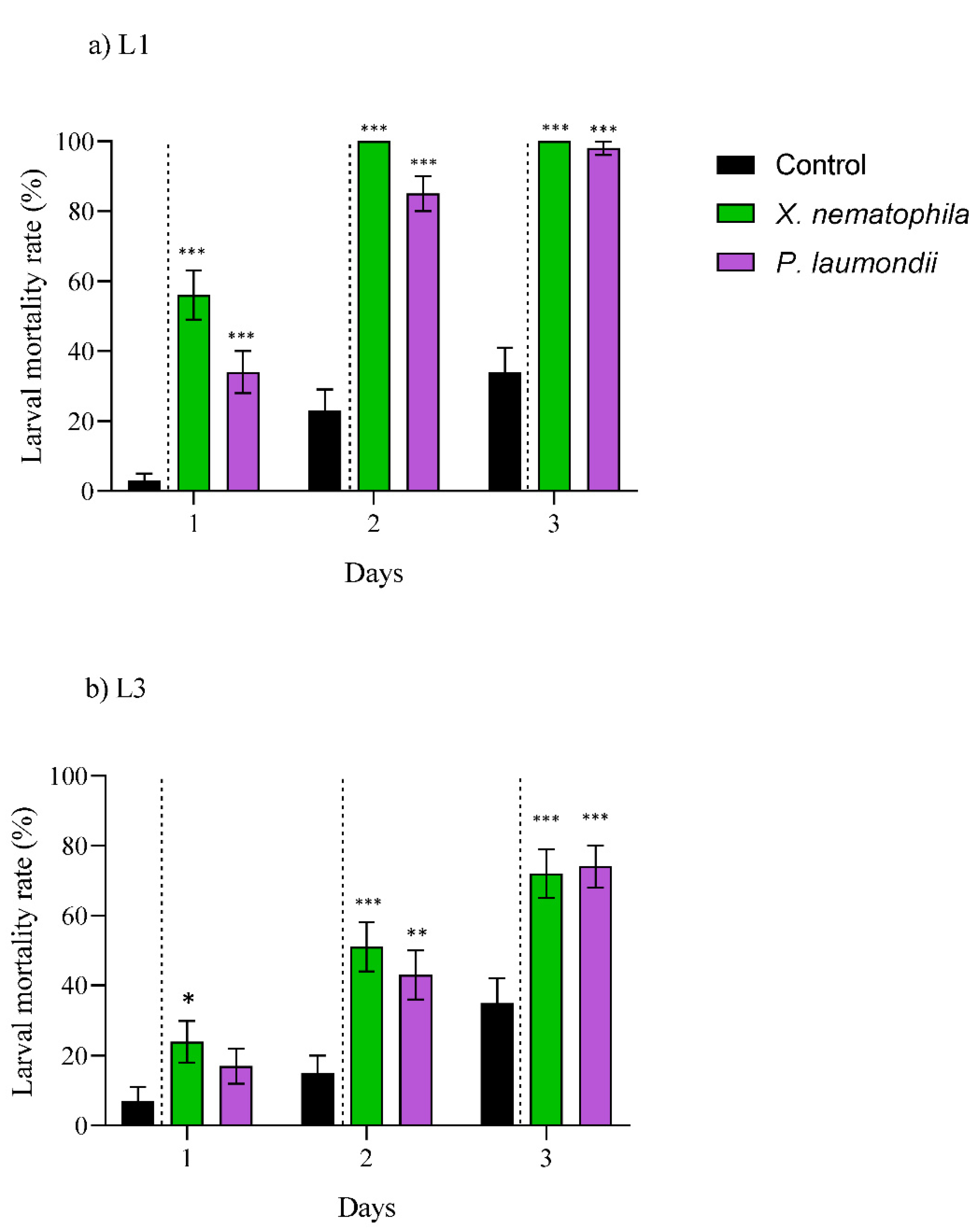
3.2. Larvicidal Effect by Bioactive Compounds Generated by the Symbiotic Bacteria
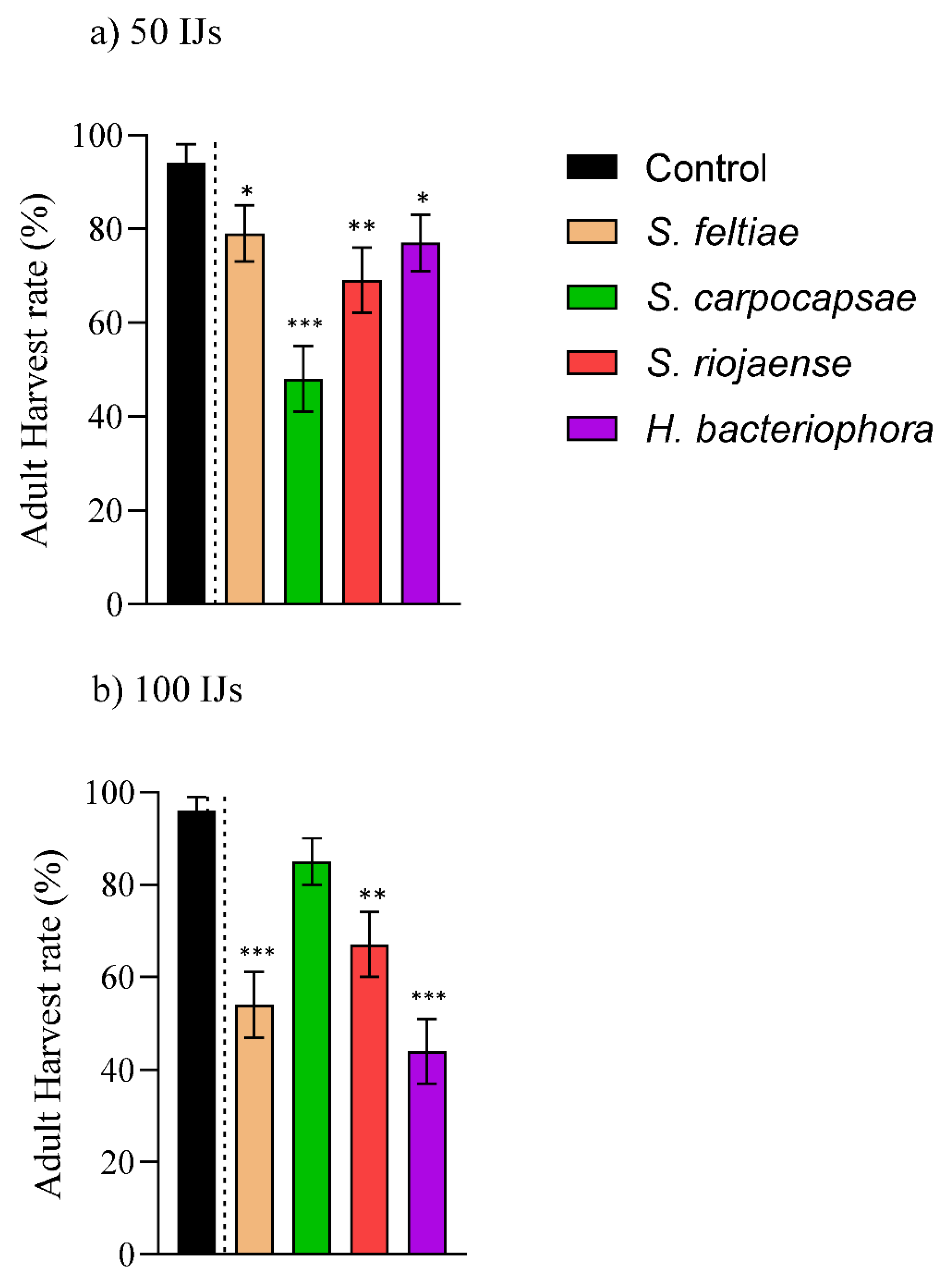
3.3. Pupicidal Effect by the Entomopathogenic Nematodes
4. Discussion
4.1. Entomopathogenic Nematodes as Biological Control Agents against Larvae and Pupae of the European Grapevine Moth
4.2. Natural Products Derived from Xenorhabdus and Photorhabdus Have Toxic Effect on Larvae
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Santos, J.A.; Fraga, H.; Malheiro, A.C.; Moutinho-Pereira, J.; Dinis, L.-T.; Correia, C.; Moriondo, M.; Leolini, L.; Dibari, C.; Costafreda-Aumedes, S.; et al. A Review of the Potential Climate Change Impacts and Adaptation Options for European Viticulture. Appl. Sci. 2020, 10, 3092. [Google Scholar] [CrossRef]
- Pertot, I.; Caffi, T.; Rossi, V.; Mugnai, L.; Hoffmann, C.; Grando, M.S.; Gary, C.; Lafond, D.; Duso, C.; Thiery, D.; et al. A critical review of plant protection tools for reducing pesticide use on grapevine and new perspectives for the implementation of IPM in viticulture. Crop Prot. 2017, 97, 70–84. [Google Scholar] [CrossRef]
- Provost, C.; Pedneault, K. The organic vineyard as a balanced ecosystem: Improved organic grape management and impacts on wine quality. Sci. Hortic. 2016, 208, 43–56. [Google Scholar] [CrossRef]
- Martín Gil, Á.; Ramos Sáez de Ojer, J.L.; Pérez, M.R. Guía de Gestión Integrada de Plagas: Uva de Transformación; Ministerio de Agricultura, Alimentación y Medio Ambiente: Madrid, Spain, 2014; ISBN 9788449113888. [Google Scholar]
- Zehnder, G.; Gurr, G.M.; Kühne, S.; Wade, M.R.; Wratten, S.D.; Wyss, E. Arthropod pest management in organic crops. Annu. Rev. Entomol. 2007, 52, 57–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacometti, M.A.; Wratten, S.D.; Walter, M. Review: Alternatives to synthetic fungicides for Botrytis cinerea management in vineyards. Aust. J. Grape Wine Res. 2010, 16, 154–172. [Google Scholar] [CrossRef]
- Gonzalez, M. Lobesia botrana: Polilla de la uva. Enología 2010, 7, 1–5. [Google Scholar]
- Varela, L.G.; Lucchi, A.; Bagnoli, B.; Nicolini, G.; Ioriatti, C. Impacts of standard wine-making process on the survival of Lobesia botrana larvae (Lepidoptera: Tortricidae) in infested grape clusters. J. Econ. Entomol. 2013, 106, 2349–2353. [Google Scholar] [CrossRef] [Green Version]
- Gilligan, T.M.; Epstein, M.E.; Passoa, S.C.; Powell, J.A.; Sage, O.C.; Brown, J.W. Discovery of Lobesia botrana ([Denis & Schiffermller]) in California: An invasive species new to North America (Lepidoptera: Tortricidae). Proc. Entomol. Soc. Washingt. 2011, 113, 14–30. [Google Scholar] [CrossRef]
- Rank, A.; Ramos, R.S.; da Silva, R.S.; Soares, J.R.S.; Picanço, M.C.; Fidelis, E.G. Risk of the introduction of Lobesia botrana in suitable areas for Vitis vinifera. J. Pest Sci. 2020, 93, 1167–1179. [Google Scholar] [CrossRef]
- Gutierrez, A.P.; Ponti, L.; Gilioli, G.; Baumgärtner, J. Climate warming effects on grape and grapevine moth (Lobesia botrana) in the Palearctic region. Agric. For. Entomol. 2018, 20, 255–271. [Google Scholar] [CrossRef] [Green Version]
- Martín-Vertedor, D.; Ferrero-García, J.J.; Torres-Vila, L.M. Global warming affects phenology and voltinism of Lobesia botrana in Spain. Agric. For. Entomol. 2010, 12, 169–176. [Google Scholar] [CrossRef]
- Reis, S.; Martins, J.; Gonçalves, F.; Carlos, C.; Santos, A.J. European grapevine moth in the Douro region: Voltinism and climatic scenarios. OENO One 2021, 55, 335–351. [Google Scholar] [CrossRef]
- Mondani, L.; Palumbo, R.; Tsitsigiannis, D.; Perdikis, D.; Mazzoni, E.; Battilani, P. Pest Management and Ochratoxin A Contamination in Grapes: A review. Toxins 2020, 12, 303. [Google Scholar] [CrossRef] [PubMed]
- López Plantey, R.; Papura, D.; Couture, C.; Thiéry, D.; Pizzuolo, P.H.; Bertoldi, M.V.; Lucero, G.S. Characterization of entomopathogenic fungi from vineyards in Argentina with potential as biological control agents against the European grapevine moth Lobesia botrana. BioControl 2019, 64, 501–511. [Google Scholar] [CrossRef]
- Castex, V.; Beniston, M.; Calanca, P.; Fleury, D.; Moreau, J. Pest management under climate change: The importance of understanding tritrophic relations. Sci. Total Environ. 2018, 616–617, 397–407. [Google Scholar] [CrossRef] [PubMed]
- Lacey, L.A.; Grzywacz, D.; Shapiro-Ilan, D.I.; Frutos, R.; Brownbridge, M.; Goettel, M.S. Insect pathogens as biological control agents: Back to the future. J. Invertebr. Pathol. 2015, 132, 1–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, S.; Koitabashi, M.; Yaginuma, D.; Anzai, M.; Fukuda, M. Potential of bioinsecticidal Bacillus thuringiensis inoculum to suppress gray mold in tomato based on induced systemic resistance. J. Phytopathol. 2019, 167, 679–685. [Google Scholar] [CrossRef]
- Shapira, I.; Keasar, T.; Harari, A.R.; Gavish-Regev, E.; Kishinevsky, M.; Steinitz, H.; Sofer-Arad, C.; Tomer, M.; Avraham, A.; Sharon, R. Does mating disruption of Planococcus ficus and Lobesia botrana affect the diversity, abundance and composition of natural enemies in Israeli vineyards? Pest Manag. Sci. 2018, 74, 1837–1844. [Google Scholar] [CrossRef]
- Ioriatti, C.; Anfora, G.; Tasin, M.; De Cristofaro, A.; Witzgall, P.; Lucchi, A. Chemical ecology and management of Lobesia botrana (Lepidoptera: Tortricidae). J. Econ. Entomol. 2011, 104, 1125–1137. [Google Scholar] [CrossRef]
- Sammaritano, J.A.; Deymié, M.; Herrera, M.; Vazquez, F.; Cuthbertson, A.G.S.; López-Lastra, C.; Lechner, B. The entomopathogenic fungus, Metarhizium anisopliae for the european grapevine moth, Lobesia botrana Den. & Schiff. (Lepidoptera: Tortricidae) and its effect to the phytopathogenic fungus, Botrytis cinerea. Egypt. J. Biol. Pest Control 2018, 28, 83. [Google Scholar] [CrossRef]
- Pérez Moreno, I.; Marco Mancebón, V.; Sáenz de Cabezón, F. Evaluación del parasitismo natural sobre crisálidas hibernants de polilla del racimo (Lobesia botrana Den. y Schiff.) en viñedos de La Rioja. Boletín Sanid. Veg. Plagas 2000, 26, 715–722. [Google Scholar]
- Gökçe, A.; Isaacs, R.; Whalon, M.E. Ovicidal, larvicidal and anti-ovipositional activities of Bifora radians and other plant extracts on the grape berry moth Paralobesia viteana (Clemens). J. Pest Sci. 2011, 84, 487–493. [Google Scholar] [CrossRef]
- Ifoulis, A.A.; Savopoulou-Soultani, M. Biological control of Lobesia botrana (Lepidoptera: Tortricidae) larvae by using different formulations of Bacillus thuringiensis in 11 vine cultivars under field conditions. J. Econ. Entomol. 2004, 97, 340–343. [Google Scholar] [CrossRef] [PubMed]
- Shapiro-Ilan, D.I.; Han, R.; Dolinksi, C. Entomopathogenic nematode production and application technology. J. Nematol. 2012, 44, 206–217. [Google Scholar]
- Griffin, C.T. Behaviour and population dynamics of entomopathogenic nematodes following application. In Nematode Pathogenesis of Insects and Other Pests; Springer: Cham, Switzerland, 2015; pp. 57–95. ISBN 9783319182667. [Google Scholar]
- Da Silva, W.J.; Pilz-Júnior, H.L.; Heermann, R.; Da Silva, O.S. The great potential of entomopathogenic bacteria Xenorhabdus and Photorhabdus for mosquito control: A review. Parasites Vectors 2020, 13, 376. [Google Scholar] [CrossRef] [PubMed]
- Karthik Raja, R.; Arun, A.; Touray, M.; Hazal Gulsen, S.; Cimen, H.; Gulcu, B.; Hazir, C.; Aiswarya, D.; Ulug, D.; Cakmak, I.; et al. Antagonists and defense mechanisms of entomopathogenic nematodes and their mutualistic bacteria. Biol. Control 2021, 152, 104452. [Google Scholar] [CrossRef]
- Boemare, N. Biology, taxonomy and systematics of Xenorhabdus and Photorhabdus. In Entomopathogenic Nematology; Gaugle, R., Ed.; CABI Publishing: Wallingford, UK, 2002; pp. 35–56. [Google Scholar]
- Adams, B.J.; Fodor, A.; Koppenhöfer, H.S.; Stackebrandt, E.; Patricia Stock, S.; Klein, M.G. Biodiversity and systematics of nematode-bacterium entomopathogens. Biol. Control 2006, 37, 32–49. [Google Scholar] [CrossRef]
- Dillman, A.R.; Chaston, J.M.; Adams, B.J.; Ciche, T.A.; Goodrich-Blair, H.; Stock, S.P.; Sternberg, P.W. An entomopathogenic nematode by any other name. PLoS Pathog. 2012, 8, e1002527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campos-Herrera, R.; Vicente-Díez, I.; Blanco-Pérez, R.; Chelkha, M.; del Mar Gonzalez-Trujillo, M.; Puelles, M.; Čepulitè, R.; Pou, A. Positioning entomopathogenic nematodes for the future viticulture: Exploring their use against biotic threats and as bioindicators of soil health. Turk. J. Zool. 2021, 45, 335–346. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.; Dolinksi, C. Entomopathogenic Nematode Application Technology. In Nematode Pathogenesis of Insects and Other Pests: Ecology and Applied Technologies for Sustainable Plant and Crop Protection; Campos-Herrera, R., Ed.; Springer International Publishing: Cham, Switzerland; Heidelberg, Germany; New York, NY, USA; Dordrecht, The Netherlands; London, UK, 2015; ISBN 9783319182667. [Google Scholar]
- Manrakhan, A.; Daneel, J.H.; Moore, S.D. The impact of naturally occurring entomopathogenic nematodes on false codling moth, Thaumatotibia leucotreta (Lepidoptera: Tortricidae), in citrus orchards. Biocontrol Sci. Technol. 2014, 24, 241–245. [Google Scholar] [CrossRef]
- Odendaal, D.; Addison, M.F.; Malan, A.P. Control of diapausing codling moth, Cydia pomonella (Lepidoptera: Tortricidae) in wooden fruit bins, using entomopathogenic nematodes (Heterorhabditidae and Steinernematidae). Biocontrol Sci. Technol. 2016, 26, 1504–1515. [Google Scholar] [CrossRef]
- de Waal, J.Y.; Addison, M.F.; Malan, A.P. Potential of Heterorhabditis zealandica (Rhabditida: Heterorhabditidae) for the control of codling moth, Cydia pomonella (Lepidoptera: Tortricidae) in semi-field trials under South African conditions. Int. J. Pest Manag. 2018, 64, 102–109. [Google Scholar] [CrossRef]
- Malan, A.P.; Diest, J.I.V.; Moore, S.D.; Addison, P. Control Options for False Codling Moth, Thaumatotibia leucotreta (Lepidoptera: Tortricidae), in South Africa, with Emphasis on the Potential Use of Entomopathogenic Nematodes and Fungi. Afr. Entomol. 2018, 26, 14–29. [Google Scholar] [CrossRef]
- Yağci, M.; Özdem, A.; Erdoğuş, F.D.; Ayan, E. Efficiency of entomopathogenic nematodes (Rhabditida: Heterorhabditidae and Steinernematidae) on the codling moth (Cydia pomonella L.) (Lepidoptera: Tortricidae) under controlled conditions. Egypt. J. Biol. Pest Control 2021, 31, 75. [Google Scholar] [CrossRef]
- du Preez, F.; Malan, A.P.; Addison, P. Potential of in vivo- and in vitro-cultured entomopathogenic nematodes to infect Lobesia vanillana (Lepidoptera: Tortricidae) under laboratory conditions. PLoS ONE 2021, 16, e90972. [Google Scholar] [CrossRef] [PubMed]
- Vicente-Díez, I.; Blanco-Pérez, R.; del Mar Gonzalez-Trujillo, M.; Pou, A.; Campos-Herrera, R. Insecticidal Effect of Entomopathogenic Nematodes and the Cell-Free Supernatant from Their Symbiotic Bacteria against Philaenus spumarius (Hemiptera: Aphrophoridae) Nymphs. Insects 2021, 12, 448. [Google Scholar] [CrossRef] [PubMed]
- Malan, A.P.; Knoetze, R.; Moore, S.D. Isolation and identification of entomopathogenic nematodes from citrus orchards in South Africa and their biocontrol potential against false codling moth. J. Invertebr. Pathol. 2011, 108, 115–125. [Google Scholar] [CrossRef]
- Steyn, V.M.; Malan, A.P.; Addison, P. Efficacy of entomopathogens against Thaumatotibia leucotreta under laboratory conditions. Entomol. Exp. Appl. 2021, 169, 449–461. [Google Scholar] [CrossRef]
- Bastidas, B.; Portillo, E.; San-Blas, E. Size does matter: The life cycle of Steinernema spp. in micro-insect hosts. J. Invertebr. Pathol. 2014, 121, 46–55. [Google Scholar] [CrossRef]
- Stock, S.P. Diversity, biology and evolutionary relationships. In Nematode Pathogenesis of Insects and Other Pests; Campos-Herrera, R., Ed.; Springer International Publishing: Cham, Switzerland; Heidelberg, Germany; New York, NY, USA; Dordrecht, The Netherlands; London, UK, 2015; pp. 3–27. [Google Scholar]
- Campbell, J.F.; Lewis, E.E.; Stock, S.P.; Nadler, S.; Kaya, H.K. Evolution of host search strategies in entomopathogenic nematodes. J. Nematol. 2003, 35, 142–145. [Google Scholar]
- Půža, V.; Campos-Herrera, R.; Blanco-Pérez, R.; Jakubíková, H.; Vicente-Díez, I.; Nermut’, J. Steinernema riojaense n. sp., a new entomopathogenic nematode (Nematoda: Steinernematidae) from Spain. Nematology 2020, 22, 825–841. [Google Scholar] [CrossRef]
- Filgueiras, C.C.; Willett, D.S. Non-lethal effects of entomopathogenic nematode infection. Sci. Rep. 2021, 11, 17090. [Google Scholar] [CrossRef]
- Chacón-Orozco, J.G.; Bueno, C.J.; Shapiro-Ilan, D.I.; Hazir, S.; Leite, L.G.; Harakava, R. Antifungal activity of Xenorhabdus spp. and Photorhabdus spp. against the soybean pathogenic Sclerotinia sclerotiorum. Sci. Rep. 2020, 10, 20649. [Google Scholar] [CrossRef]
- Eroglu, C.; Cimen, H.; Ulug, D.; Karagoz, M.; Hazir, S.; Cakmak, I. Acaricidal effect of cell-free supernatants from Xenorhabdus and Photorhabdus bacteria against Tetranychus urticae (Acari: Tetranychidae). J. Invertebr. Pathol. 2019, 160, 61–66. [Google Scholar] [CrossRef]
- Bussaman, P.; Sobanboa, S.; Grewal, P.S.; Chandrapatya, A. Pathogenicity of additional strains of Photorhabdus and Xenorhabdus (Enterobacteriaceae) to the mushroom mite Luciaphorus perniciosus (Acari: Pygmephoridae). Appl. Entomol. Zool. 2009, 44, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Chandler, D.; Bailey, A.S.; Mark Tatchell, G.; Davidson, G.; Greaves, J.; Grant, W.P. The development, regulation and use of biopesticides for integrated pest management. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 1987–1998. [Google Scholar] [CrossRef] [PubMed]
- Schünemann, R.; Knaak, N.; Fiuza, L.M. Mode of Action and Specificity of Bacillus thuringiensis Toxins in the Control of Caterpillars and Stink Bugs in Soybean Culture. ISRN Microbiol. 2014, 2014, 135675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orozco, R.A.; Molnár, I.; Bode, H.; Stock, S.P. Bioprospecting for secondary metabolites in the entomopathogenic bacterium Photorhabdus luminescens subsp. sonorensis. J. Invertebr. Pathol. 2016, 141, 45–52. [Google Scholar] [CrossRef]
- Crowder, D.W.; Jabbour, R. Relationships between biodiversity and biological control in agroecosystems: Current status and future challenges. Biol. Control 2014, 75, 8–17. [Google Scholar] [CrossRef]
- Blanco-Pérez, R.; Sáenz-Romo, M.G.; Vicente-Díez, I.; Ibáñez-Pascual, S.; Martínez-Villar, E.; Marco-Mancebón, V.S.; Pérez-Moreno, I.; Campos-Herrera, R. Impact of vineyard ground cover management on the occurrence and activity of entomopathogenic nematodes and associated soil organisms. Agric. Ecosyst. Environ. 2020, 301, 107028. [Google Scholar] [CrossRef]
- Karimi, B.; Cahurel, J.Y.; Gontier, L.; Charlier, L.; Chovelon, M.; Mahé, H.; Ranjard, L. A meta-analysis of the ecotoxicological impact of viticultural practices on soil biodiversity. Environ. Chem. Lett. 2020, 18, 1947–1966. [Google Scholar] [CrossRef]
- Sáenz-Romo, M.G.; Veas-Bernal, A.; Martínez-García, H.; Campos-Herrera, R.; Ibáñez-Pascual, S.; Martínez-Villar, E.; Pérez-Moreno, I.; Marco-Mancebón, V.S. Ground cover management in a Mediterranean vineyard: Impact on insect abundance and diversity. Agric. Ecosyst. Environ. 2019, 283, 106571. [Google Scholar] [CrossRef]
- Mnif, I.; Ghribi, D. Potential of bacterial derived biopesticides in pest management. Crop Prot. 2015, 77, 52–64. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Principal Target | References | ||||
|---|---|---|---|---|---|
| Biological control agents | Entomopathogenic fungi | Beauviera bassiana | Pupae | López Plantey et al., 2019 [15] | |
| Metarhizium spp. (M. robertsii; M. anisopliae) | Larvae | López Plantey et al., 2019; Sammaritano et al., 2018 [15,21] | |||
| Bacterias | B. thuringiensis | Larvae Adults | Ioriatti et al., 2011 [20] | ||
| Artropods | Predator | Chrysoperla sp. | Eggs Pupae | Castex et al., 2018 [16] | |
| Parasitoids | Trichogramma sp. | Eggs | Pérez Moreno et al., 2000 [22] | ||
| Dibrachis sp. | Pupae | ||||
| Entomopathogenic nematodes | Steinernema sp. Heterohabditis sp. | Larvae Pupae | Current research | ||
| Biotechnical control tools | Pheromone-mediated Malting Disruption (MD) | Adults | Ioriatti et al., 2011; Shapira et al., 2018 [19,20] | ||
| Insecticidal-plants extracts | Bifora radians | Larvae | Gökçe et al., 2011 [23] | ||
| Insecticidal-bacterium based product | Bacillus thuringiensis | Larvae | Ifoulis and Savopoulou-Soultani, 2004 [24] | ||
| Xhenorhabdus sp. Photorhabdus sp. | Larvae | Current reserch | |||
| Entomopathogenic Nematodes Species | Population | ITS-Gen Bank Accession Number | Bacterial Species | ITS-Gen Bank Accession Number |
|---|---|---|---|---|
| Steinernema feltiae | RM-107 | MW480131 | Xenorhabdus bovienii | MW467374 |
| Steinernema carpocapsae | All | MW574913 | Xenorhabdus nematophila | MW574906 |
| Steinernema riojaense | RM-30 | MK503133 | Xenorhabdus kozodoii | MW467375 |
| Heterorhabditis bacteriophora | RM-102 | MW480132 | Photorabhdus laumondii subsp. laumondii | MW574908 |
| Day | Treatment | Tested Stage of EGVM | Number of Insect Tested | Slope ± SE | LC50 (95% Confidence Intervals) | LC90 (95% Confidence Intervals) | X2 |
|---|---|---|---|---|---|---|---|
| 3 | Sfe | L5 | 300 | 0.918 ± 0.222 | 5.229 (3.469–10.297) | 130.362 (38.089–3766.676) | 1.383 |
| Sca | L5 | 300 | 2.112 ± 0.623 | 0.352 (0.047–0.629) | 1.424 (0.954–2.074) | 1.398 | |
| 4 | Sfe | L5 | 300 | 1.001 ± 0.223 | 3. 692 (2.499–5.821) | 70.318 (26.189–801.379) | 4.395 |
| Sca | L5 | 300 | 3.248 ± 1.073 | 0.530 (0.110–0.769) | 1.315 (1.024–1.887) | 0.065 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vicente-Díez, I.; Blanco-Pérez, R.; Chelkha, M.; Puelles, M.; Pou, A.; Campos-Herrera, R. Exploring the Use of Entomopathogenic Nematodes and the Natural Products Derived from Their Symbiotic Bacteria to Control the Grapevine Moth, Lobesia botrana (Lepidoptera: Tortricidae). Insects 2021, 12, 1033. https://doi.org/10.3390/insects12111033
Vicente-Díez I, Blanco-Pérez R, Chelkha M, Puelles M, Pou A, Campos-Herrera R. Exploring the Use of Entomopathogenic Nematodes and the Natural Products Derived from Their Symbiotic Bacteria to Control the Grapevine Moth, Lobesia botrana (Lepidoptera: Tortricidae). Insects. 2021; 12(11):1033. https://doi.org/10.3390/insects12111033
Chicago/Turabian StyleVicente-Díez, Ignacio, Rubén Blanco-Pérez, Maryam Chelkha, Miguel Puelles, Alicia Pou, and Raquel Campos-Herrera. 2021. "Exploring the Use of Entomopathogenic Nematodes and the Natural Products Derived from Their Symbiotic Bacteria to Control the Grapevine Moth, Lobesia botrana (Lepidoptera: Tortricidae)" Insects 12, no. 11: 1033. https://doi.org/10.3390/insects12111033
APA StyleVicente-Díez, I., Blanco-Pérez, R., Chelkha, M., Puelles, M., Pou, A., & Campos-Herrera, R. (2021). Exploring the Use of Entomopathogenic Nematodes and the Natural Products Derived from Their Symbiotic Bacteria to Control the Grapevine Moth, Lobesia botrana (Lepidoptera: Tortricidae). Insects, 12(11), 1033. https://doi.org/10.3390/insects12111033