
Ozone Induces Distress Behaviors in Fig Wasps with a Reduced Chance of Recovery



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Biological Model
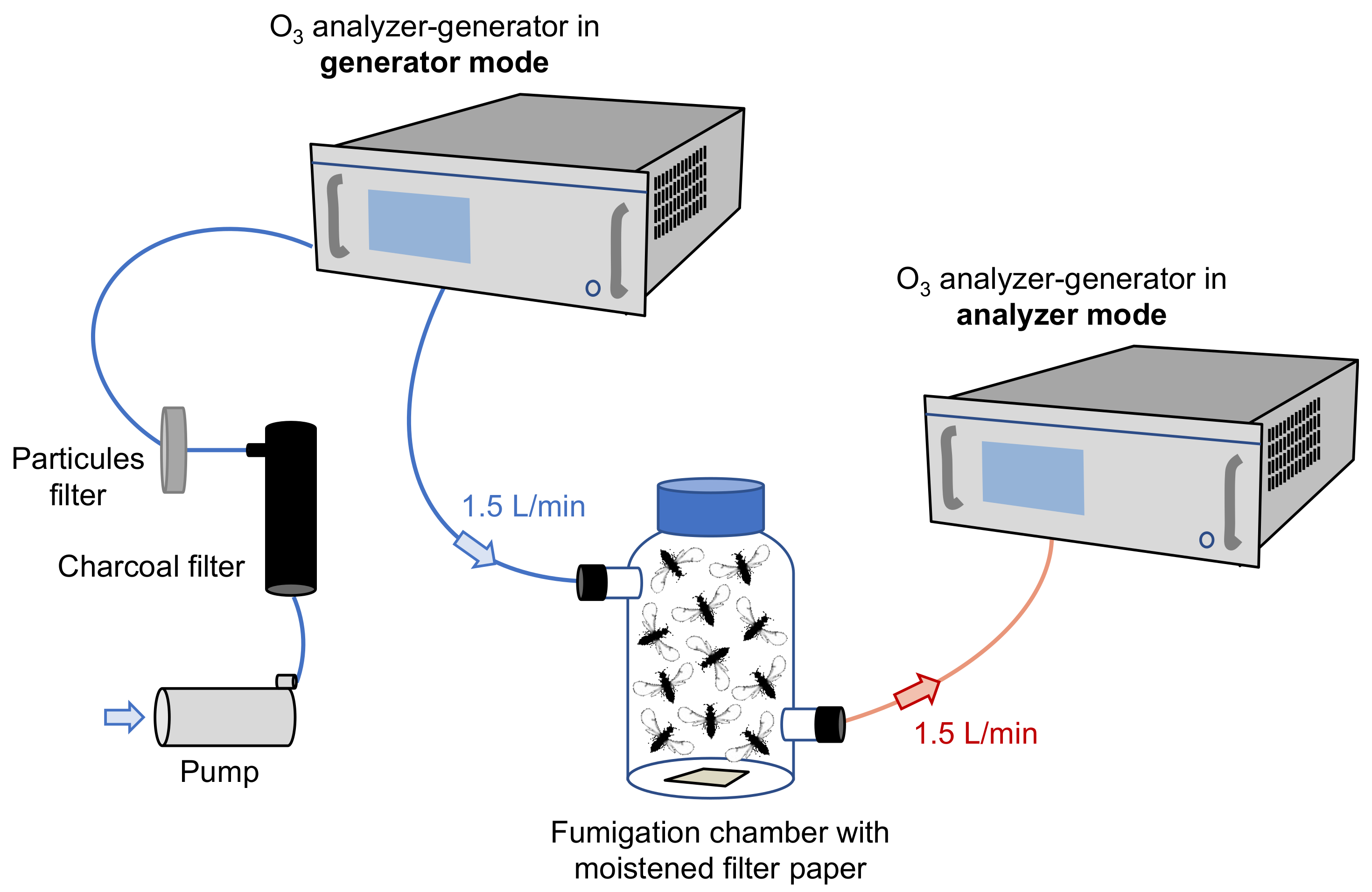
2.2. Ozone Exposure and Behavioral Assays
2.3. Statistical Analyses
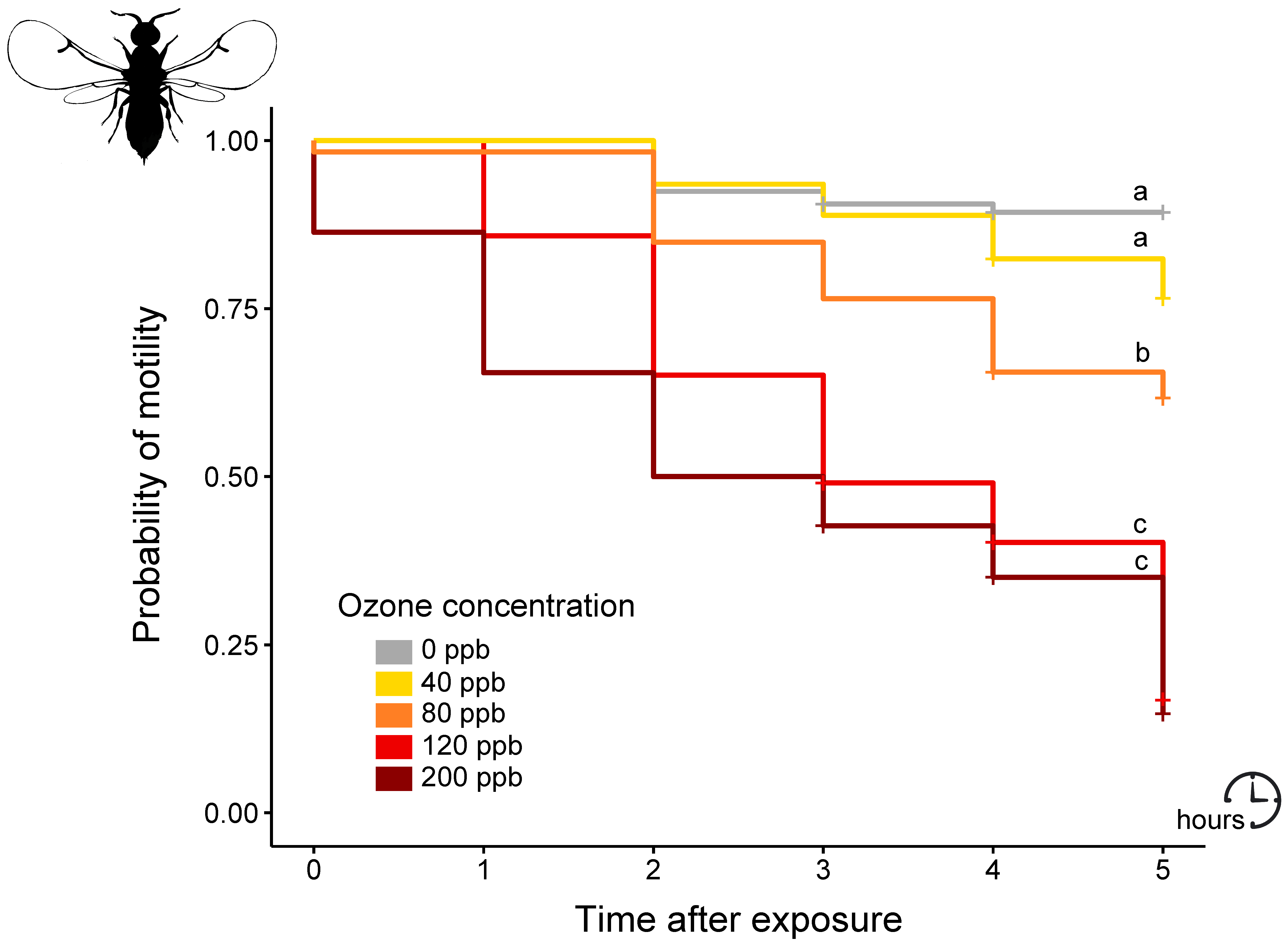
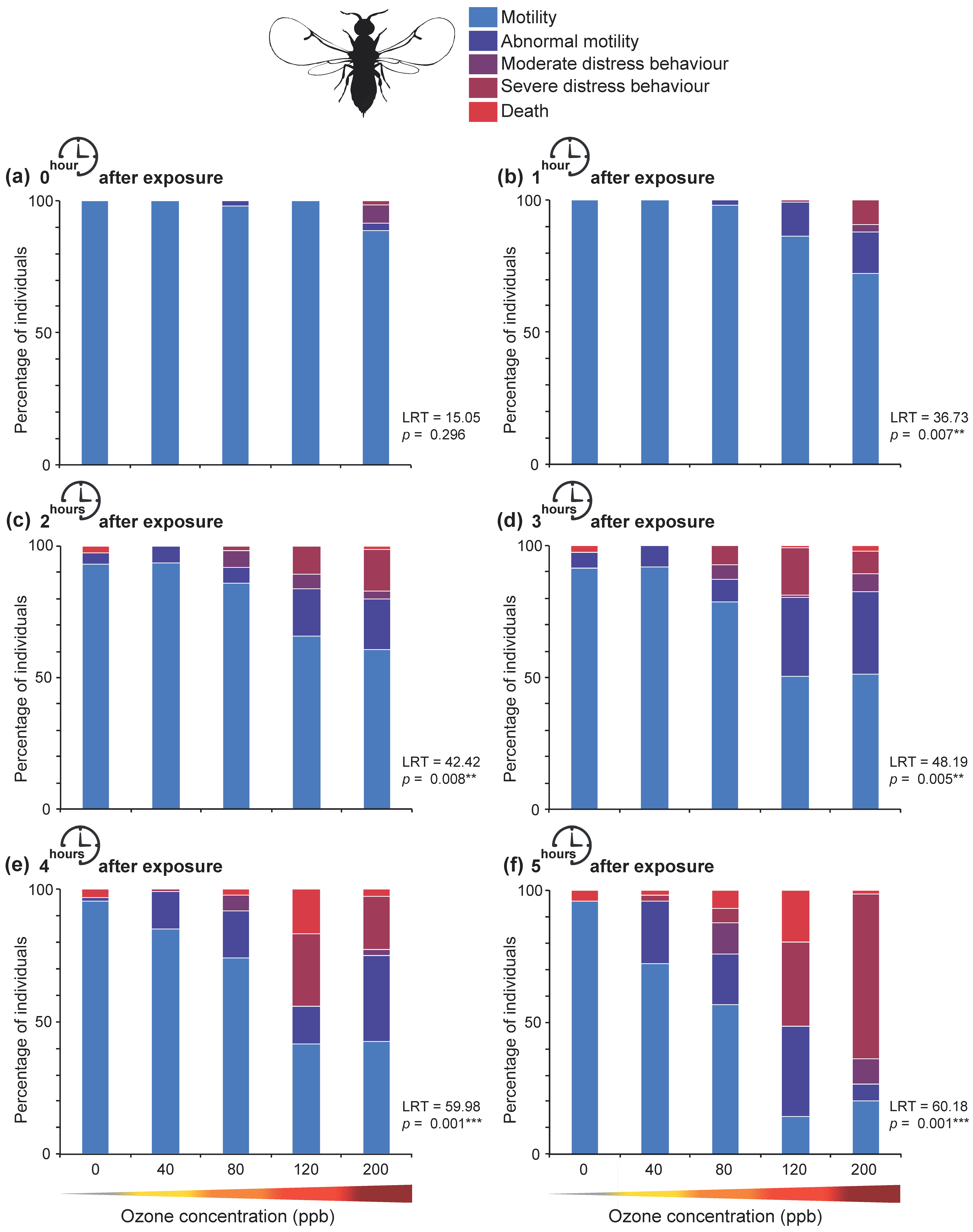
3. Results and Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
| Exposure at 0 ppb O3 (n = 10 with 11 ± 3 Individuals per Trial) | ||||||
| 0 h | 1 h | 2 h | 3 h | 4 h | 5 h | |
| Motility | 11 ± 3 | 11 ± 3 | 10 ± 3 | 10 ± 3 | 9 ± 3 | 9 ± 3 |
| Abnormal motility | 0 ± 0 | 0 ± 0 | 1 ± 2 | 1 ± 2 | 0 ± 0 | 0 ± 0 |
| Moderate distress behavior | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| Severe distress behavior | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| Death | 0 ± 0 | 0 ± 0 | 0 ± 1 | 0 ± 1 | 0 ± 1 | 0 ± 1 |
| Exposure at 40 ppb O3 (n = 10 with 11 ± 2 Individuals per Trial) | ||||||
| 0 h | 1 h | 2 h | 3 h | 4 h | 5 h | |
| Motility | 11 ± 2 | 11 ± 2 | 10 ± 2 | 10 ± 2 | 9 ± 3 | 8 ± 3 |
| Abnormal motility | 0 ± 0 | 0 ± 0 | 1 ± 1 | 1 ± 1 | 2 ± 2 | 3 ± 2 |
| Moderate distress behavior | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| Severe distress behavior | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| Death | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| Exposure at 80 ppb O3 (n = 11 with 11 ± 3 Individuals per Trial) | ||||||
| 0 h | 1 h | 2 h | 3 h | 4 h | 5 h | |
| Motility | 11 ± 3 | 11 ± 3 | 9 ± 3 | 8 ± 3 | 8 ± 2 | 7 ± 3 |
| Abnormal motility | 0 ± 0 | 0 ± 0 | 1 ± 1 | 1 ± 1 | 2 ± 2 | 2 ± 3 |
| Moderate distress behavior | 0 ± 0 | 0 ± 0 | 1 ± 2 | 1 ± 1 | 1 ± 1 | 1 ± 2 |
| Severe distress behavior | 0 ± 0 | 0 ± 0 | 0 ± 1 | 1 ± 3 | 0 ± 0 | 1 ± 2 |
| Death | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 1 | 1 ± 1 |
| Exposure at 120 ppb O3 (n = 9 with 12 ± 1 Individuals per Trial) | ||||||
| 0 h | 1 h | 2 h | 3 h | 4 h | 5 h | |
| Motility | 12 ± 1 | 10 ± 2 | 8 ± 4 | 6 ± 5 | 5 ± 5 | 2 ± 3 |
| Abnormal motility | 0 ± 0 | 2 ± 2 | 2 ± 3 | 4 ± 4 | 2 ± 2 | 4 ± 5 |
| Moderate distress behavior | 0 ± 0 | 0 ± 0 | 1 ± 1 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
| Severe distress behavior | 0 ± 0 | 0 ± 0 | 1 ± 2 | 2 ± 4 | 3 ± 5 | 4 ± 5 |
| Death | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 2 ± 5 | 3 ± 6 |
| Exposure at 200 ppb O3 (n = 10 with 11 ± 3 Individuals per Trial) | ||||||
| 0 h | 1 h | 2 h | 3 h | 4 h | 5 h | |
| Motility | 9 ± 4 | 7 ± 5 | 6 ± 3 | 5 ± 3 | 4 ± 2 | 2 ± 2 |
| Abnormal motility | 0 ± 1 | 2 ± 4 | 3 ± 4 | 4 ± 4 | 4 ± 4 | 1 ± 1 |
| Moderate distress behavior | 1 ± 3 | 0 ± 1 | 0 ± 1 | 1 ± 3 | 0 ± 1 | 2 ± 4 |
| Severe distress behavior | 0 ± 1 | 1 ± 3 | 2 ± 4 | 1 ± 4 | 3 ± 5 | 7 ± 4 |
| Death | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 | 0 ± 0 |
References
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global Pollinator Declines: Trends, Impacts and Drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Goulson, D.; Lye, G.; Darvill, B. Decline and Conservation of Bumble Bees. Annu. Rev. Entomol. 2008, 53, 191–208. [Google Scholar] [CrossRef] [PubMed]
- Winfree, R.; Aguilar, R.; Vázquez, D.P.; Lebuhn, G.; Aizen, M. A Meta-Analysis of Bees’Responses to Anthropogenic Disturbance. Ecology 2009, 90, 2068–2076. [Google Scholar] [CrossRef] [PubMed]
- Kevan, P.G.; Greco, C.F.; Belaoussoff, S. Log-Normality of Biodiversity and Abundance in Diagnosis and Measuring of Ecosystemic Health: Pesticide Stress on Pollinators on Blueberry Heaths. J. Appl. Ecol. 1997, 34, 1122. [Google Scholar] [CrossRef]
- Halm, M.-P.; Rortais, A.; Arnold, G.; Touffet-Briens, F. Modes of Honeybees Exposure to Systemic Insecticides: Estimated amounts of Contaminated Pollen and Nectar Consumed by Different Categories of Bees. Apidologie 2005, 36, 71–83. [Google Scholar] [CrossRef] [Green Version]
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemüller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel Declines in Pollinators and Insect-Pollinated Plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef]
- Williams, P.H.; Araújo, M.B.; Rasmont, P. Can Vulnerability Among British Bumblebee (Bombus) Species Be Explained by Niche Position and Breadth? Biol. Conserv. 2007, 138, 493–505. [Google Scholar] [CrossRef]
- Dormann, C.F.; Schweiger, O.; Arens, P.; Augenstein, I.; Aviron, S.; Bailey, D.; Baudry, J.; Billeter, R.; Bugter, R.; Bukácek, R.; et al. Prediction Uncertainty of Environmental Change Effects on Temperate European Biodiversity. Ecol. Lett. 2008, 11, 235–244. [Google Scholar] [CrossRef]
- Agathokleous, E.; Feng, Z.; Oksanen, E.; Sicard, P.; Wang, Q.; Saitanis, C.J.; Araminiene, V.; Blande, J.D.; Hayes, F.; Calatayud, V.; et al. Ozone Affects Plant, Insect, and Soil Microbial Communities: A Threat to Terrestrial Ecosystems and Biodiversity. Sci. Adv. 2020, 6, eabc1176. [Google Scholar] [CrossRef] [PubMed]
- Blande, J.D. Effects of Air Pollution on Plant–Insect Interactions Mediated by Olfactory and Visual Cues. Curr. Opin. Environ. Sci. Health 2021, 19, 100228. [Google Scholar] [CrossRef]
- Wong, B.B.M.; Candolin, U. Behavioral Responses to Changing Environments. Behav. Ecol. 2015, 26, 665–673. [Google Scholar] [CrossRef] [Green Version]
- Gérard, M.; Vanderplanck, M.; Wood, T.; Michez, D. Global Warming and Plant–Pollinator Mismatches. Emerg. Top. Life Sci. 2020, 4, 77–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furlan, E.; Stoklosa, J.; Griffiths, J.; Gust, N.; Ellis, R.; Huggins, R.M.; Weeks, A.R. Small Population Size and Extremely Low Levels of Genetic Diversity in Island Populations of the Platypus, Ornithorhynchus anatinus. Ecol. Evol. 2012, 2, 844–857. [Google Scholar] [CrossRef] [PubMed]
- Augusiak, J.; Brink, P.V.D. The Influence of Insecticide Exposure and Environmental Stimuli on the Movement Behaviour and Dispersal of a Freshwater Isopod. Ecotoxicology 2016, 25, 1338–1352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iriti, M.; Faoro, F. Oxidative Stress, the Paradigm of Ozone Toxicity in Plants and Animals. Water Air Soil Pollut. 2008, 187, 285–301. [Google Scholar] [CrossRef]
- Vanderplanck, M.; Lapeyre, B.; Brondani, M.; Opsommer, M.; Dufay, M.; Hossaert-McKey, M.; Proffit, M. Ozone Pollution Alters Olfaction and Behavior of Pollinators. Antioxidants 2021, 10, 636. [Google Scholar] [CrossRef]
- Cooper, O.R.; Parrish, D.D.; Ziemke, J.R.; Balashov, N.V.; Cupeiro, M.; Galbally, I.; Gilge, S.; Horowitz, L.W.; Jensen, N.R.; Lamarque, J.-F.; et al. Global Distribution and Trends of Tropospheric Ozone: An Observation-Based Review. Elem. Sci. Anth. 2014, 2, 000029. [Google Scholar] [CrossRef]
- Kjellberg, F.; Doumesche, B.; Bronstein, J.L. Longevity of a Fig Wasp (Blastophaga psenes). Proc. K. Ned. Akad. Wet. Ser. C 1988, 91, 117–122. [Google Scholar]
- Proffit, M.; Lapeyre, B.; Buatois, B.; Deng, X.; Arnal, P.; Gouzerh, F.; Carrasco, D.; Hossaert-Mckey, M. Chemical Signal is in the Blend: Bases of Plant-Pollinator Encounter in a Highly Specialized Interaction. Sci. Rep. 2020, 10, 10071. [Google Scholar] [CrossRef] [PubMed]
- Vautard, R.; Honore, C.; Beekmann, M.; Rouil, L. Simulation of Ozone During the August 2003 Heat Wave and Emission Control Scenarios. Atmospheric Environ. 2005, 39, 2957–2967. [Google Scholar] [CrossRef]
- Solberg, S.; Hov, O.; Søvde, A.; Isaksen, I.S.A.; Coddeville, P.; De Backer, H.; Forster, C.; Orsolini, Y.; Uhse, K. European Surface Ozone in the Extreme Summer 2003. J. Geophys. Res. Space Phys. 2008, 113. [Google Scholar] [CrossRef] [Green Version]
- Team, R.R. A Language and Environment for Statistical Computing, Vienna, R Foundation for Statistical Computing. 2017. Available online: http://www.R-project.org2009 (accessed on 1 October 2021).
- Wang, Y.; Naumann, U.; Eddelbuettel, D.; Wilshire, J.; Warton, D.I. Mvabund Statistical Methods for Analysing Multivariate Abundance Data. R Package Version 4.0.1. 2019. Available online: https://CRAN.R-project.org/package=mvabund (accessed on 1 October 2021).
- Wang, Y.; Naumann, U.; Wright, S.; Warton, D.I. Mvabund—An R Package for Model-Based Analysis of Multivariate Abundance Data. Methods Ecol. Evol. 2012, 3, 471–474. [Google Scholar] [CrossRef]
- Warton, D.I.; Wright, S.T.; Wang, Y. Distance-Based Multivariate Analyses Confound Location and Dispersion Effects. Methods Ecol. Evol. 2012, 3, 89–101. [Google Scholar] [CrossRef]
- Warton, D.I. Regularized Sandwich Estimators for Analysis of High-Dimensional Data Using Generalized Estimating Equations. Biometrics 2011, 67, 116–123. [Google Scholar] [CrossRef]
- García, L.V. Escaping the Bonferroni Iron Claw in Ecological Studies. Oikos 2004, 105, 657–663. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, S. A Farewell to Bonferroni: The Problems of Low Statistical Power and Publication Bias. Behav. Ecol. 2004, 15, 1044–1045. [Google Scholar] [CrossRef]
- Kodrík, D.; Bednářová, A.; Zemanová, M.; Krishnan, N. Hormonal Regulation of Response to Oxidative Stress in Insects—An Update. Int. J. Mol. Sci. 2015, 16, 25788–25816. [Google Scholar] [CrossRef] [Green Version]
- Telesnicki, M.C.; Martínez-Ghersa, M.A.; Arneodo, J.D.; Ghersa, C.M. Direct Effect of Ozone Pollution on Aphids: Revisiting the Evidence at Individual and Population Scales. Entomol. Exp. Appl. 2015, 155, 71–79. [Google Scholar] [CrossRef]
- Beaulieu, M.; Gillen, E.; Hahn, S.; Pape, J.M.; Fischer, K. Behavioural Antioxidant Strategies to Cope with High Temperatures: A Study in a Tropical Butterfly. Anim. Behav. 2015, 109, 89–99. [Google Scholar] [CrossRef]
- Bonvehí, J.S.; Torrentó, M.S.; Lorente, E.C. Evaluation of Polyphenolic and Flavonoid Compounds in Honeybee-Collected Pollen Produced in Spain. J. Agric. Food Chem. 2001, 49, 1848–1853. [Google Scholar] [CrossRef]
- Johnson, K.S.; Felton, G.W. Plant Phenolics as Dietary Antioxidants for Herbivorous Insects: A Test with Genetically Modified Tobacco. J. Chem. Ecol. 2001, 27, 2579–2597. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vanderplanck, M.; Lapeyre, B.; Lucas, S.; Proffit, M. Ozone Induces Distress Behaviors in Fig Wasps with a Reduced Chance of Recovery. Insects 2021, 12, 995. https://doi.org/10.3390/insects12110995
Vanderplanck M, Lapeyre B, Lucas S, Proffit M. Ozone Induces Distress Behaviors in Fig Wasps with a Reduced Chance of Recovery. Insects. 2021; 12(11):995. https://doi.org/10.3390/insects12110995
Chicago/Turabian StyleVanderplanck, Maryse, Benoit Lapeyre, Shéhérazade Lucas, and Magali Proffit. 2021. "Ozone Induces Distress Behaviors in Fig Wasps with a Reduced Chance of Recovery" Insects 12, no. 11: 995. https://doi.org/10.3390/insects12110995
APA StyleVanderplanck, M., Lapeyre, B., Lucas, S., & Proffit, M. (2021). Ozone Induces Distress Behaviors in Fig Wasps with a Reduced Chance of Recovery. Insects, 12(11), 995. https://doi.org/10.3390/insects12110995