
Physiological and Population Responses of Nilaparvata lugens after Feeding on Drought-Stressed Rice

 , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Biomaterials and Design
2.2. Drought Stress Treatments
2.3. BPH Population Numbers
2.4. Impact of Drought Stress on the Expression of Ovarian Development Genes
2.5. Anatomy of BPH Ovaries
2.6. Determination of Water Content in BPH
2.7. Determination of Osmotic Pressure in BPH Hemolymph
2.8. Glucose and Total Amino Acid Content in BPH
2.9. Determination of Glutathione S-Transferase Activity in BPH
2.10. Ultrastructure of Flight Muscles in BPH
2.11. Analysis of Transmission Electron Micrographs
2.12. Statistical Analysis
3. Results
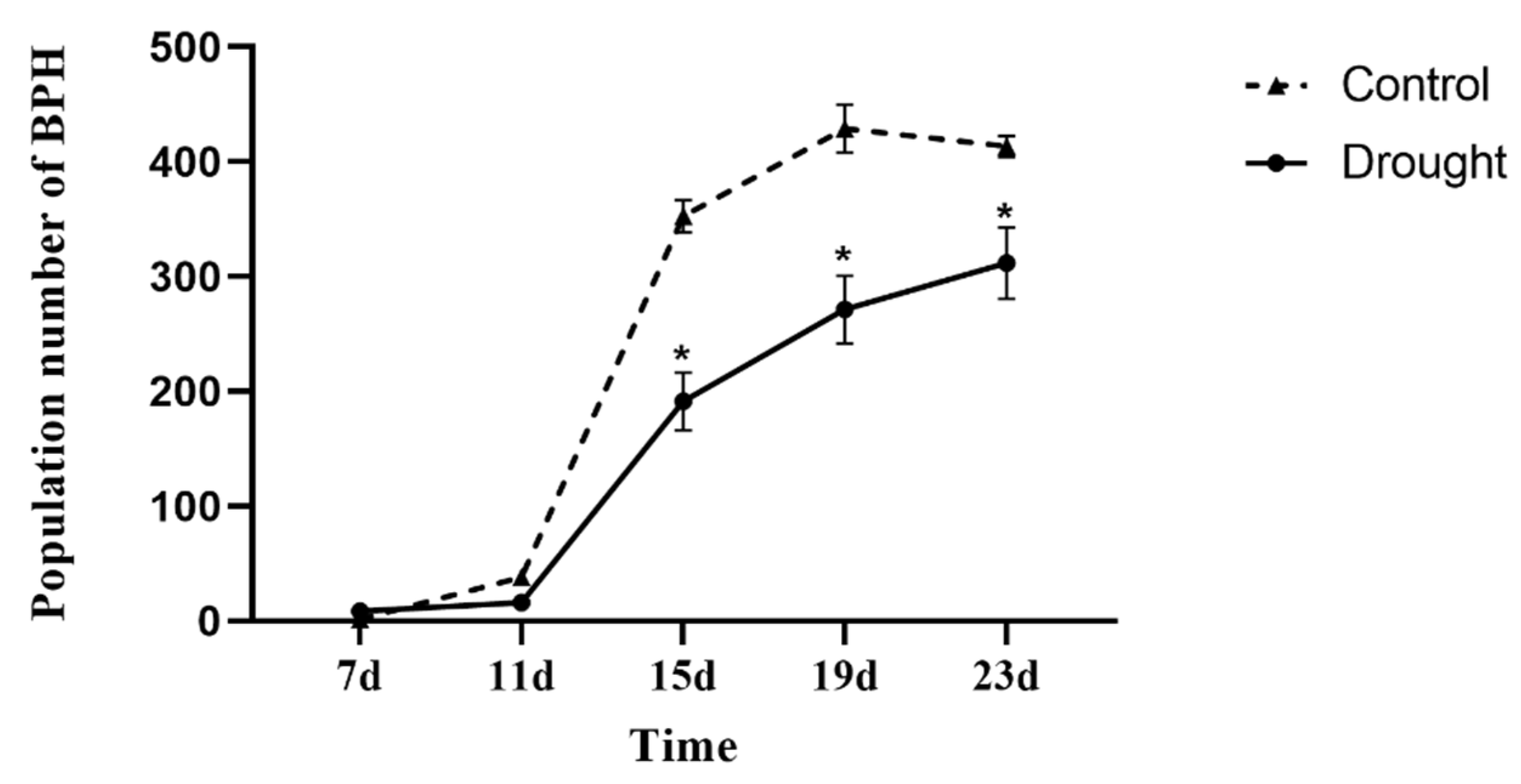
3.1. Population Dynamics of BPH
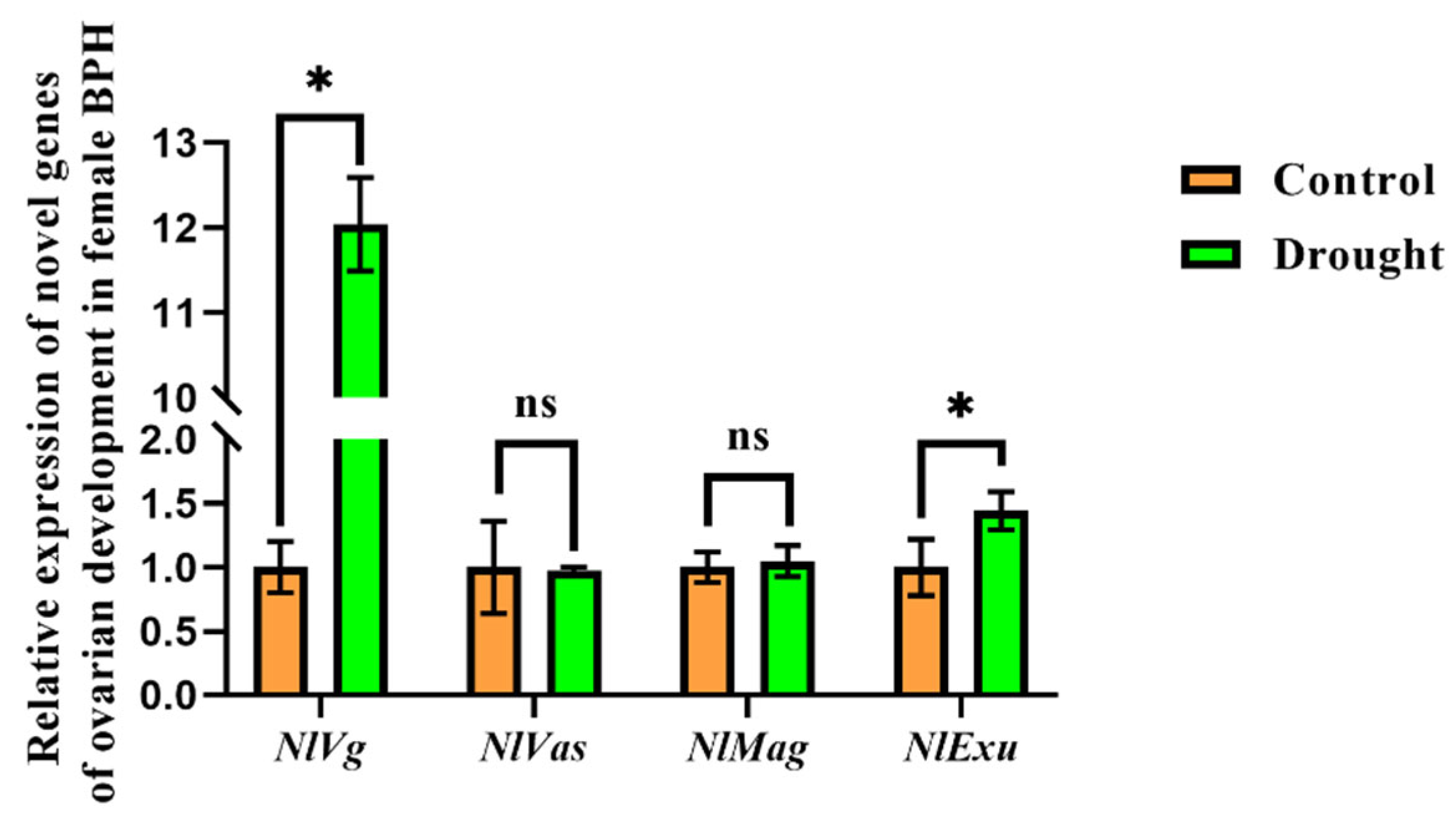
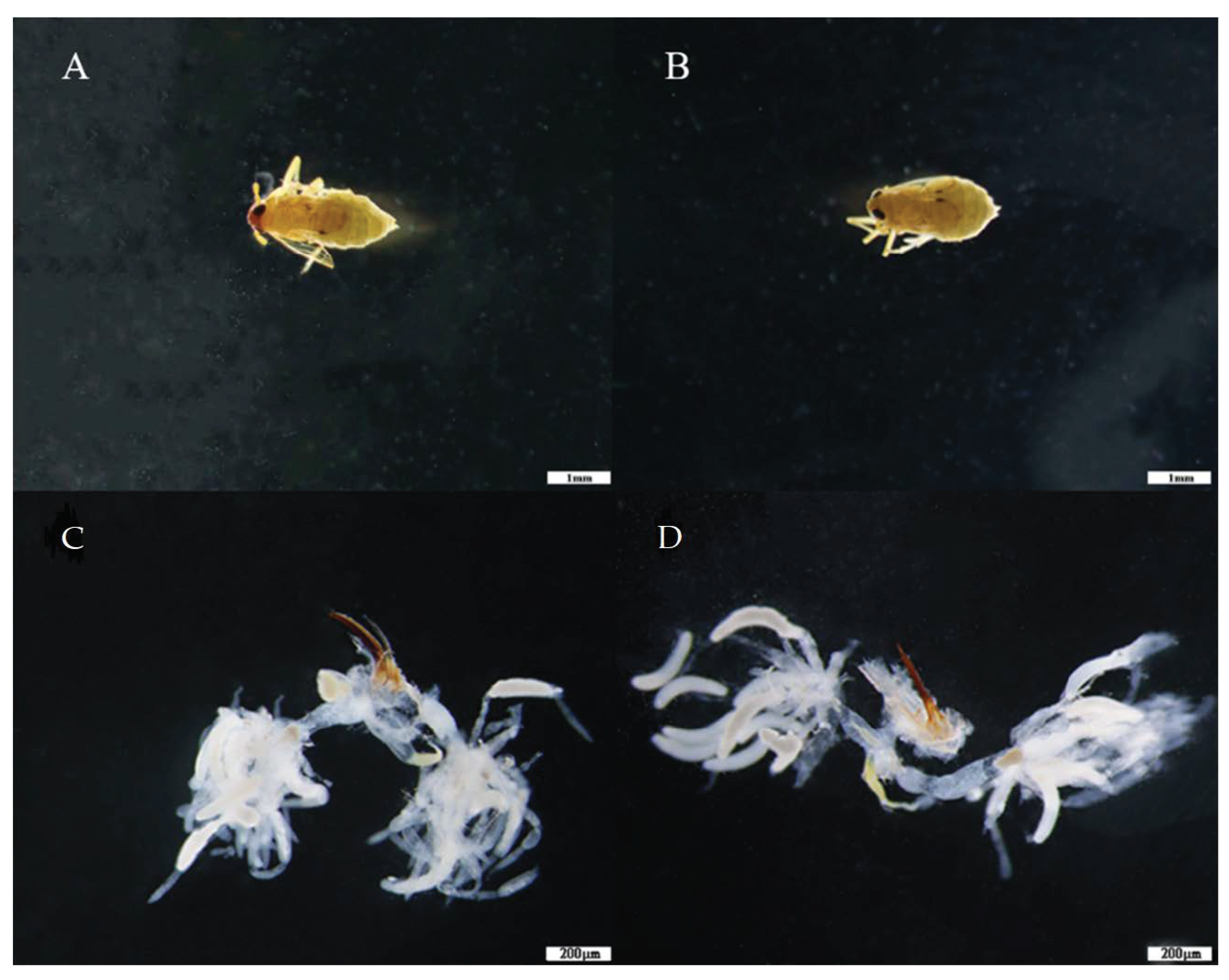
3.2. Effects of Drought Stress on the Fecundity of BPH
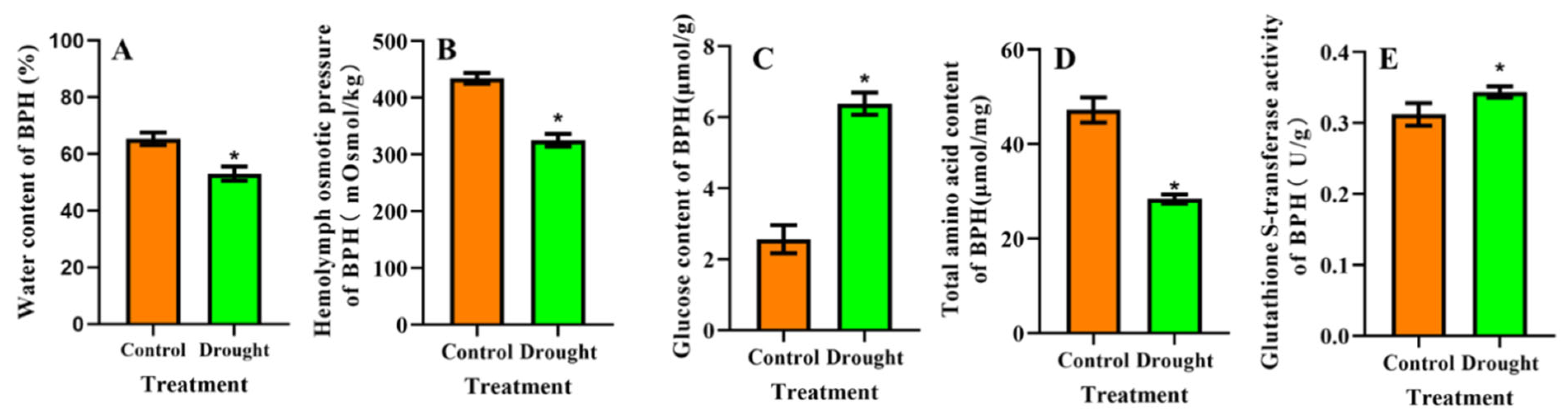
3.3. Physiological Changes in BPH after Feeding on Drought-Stressed Rice
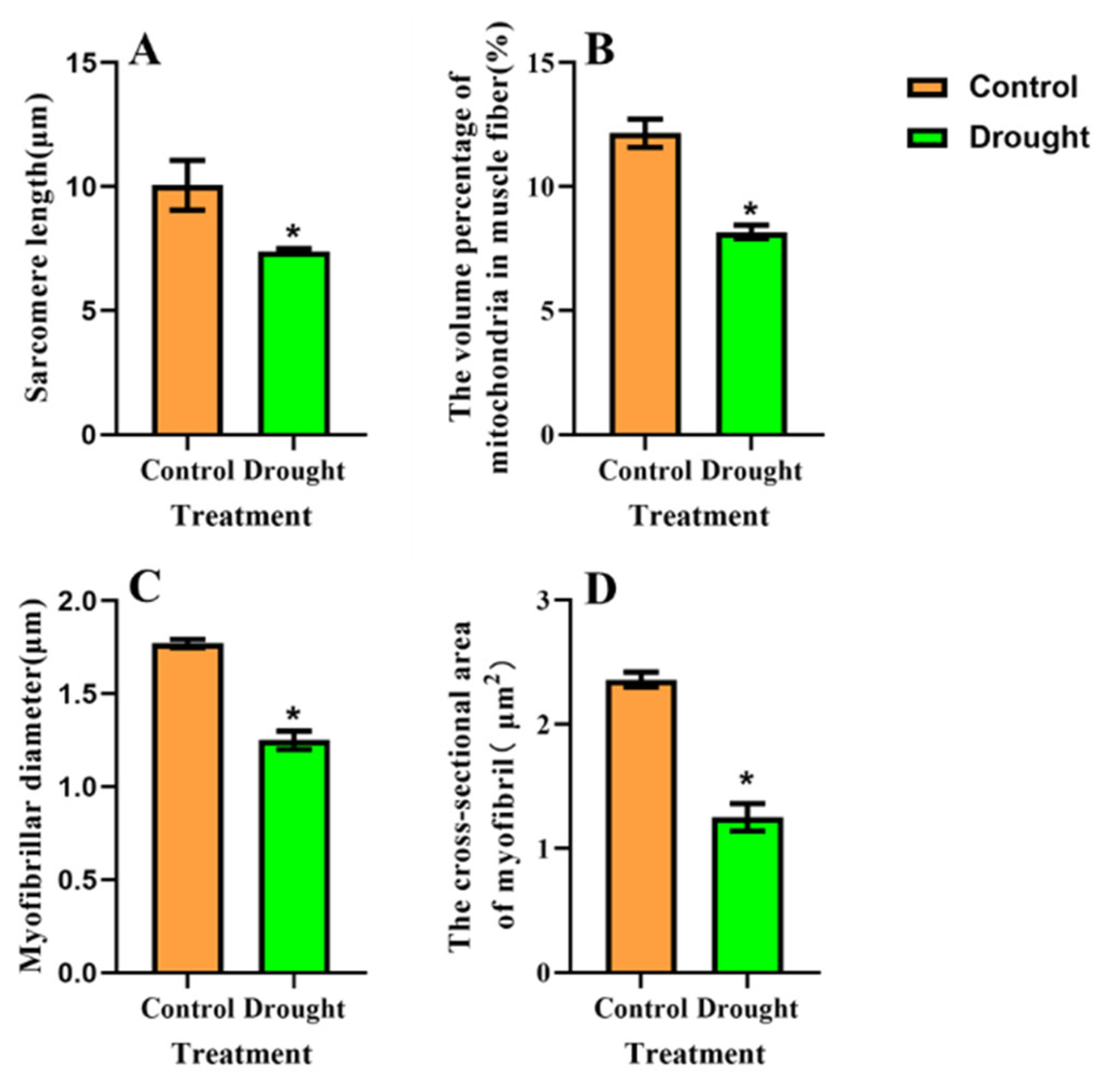
3.4. Ultrastructural Changes in BPH Flight Muscles after Feeding on Drought-Stressed Rice
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhou, R.; Yu, X.; Ottosen, C.O.; Rosenqvist, E.; Zhao, L.; Wang, Y.; Yu, W.; Zhao, T.; Wu, Z. Drought stress had a predominant effect over heat stress on three tomato cultivars subjected to combined stress. BMC Plant Biol. 2017, 17, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deutsch, C.A.; Tewksbury, J.J.; Tigchelaar, M.; Battisti, D.S.; Merrill, S.C.; Huey, R.B.; Naylor, R.L. Increase in crop losses to insect pests in a warming climate. Science 2018, 361, 916–919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, C.B.; Chirumamilla, A.; Hartman, G.L. Resistance and virulence in the soybean-Aphis glycines interaction. Euphytica 2012, 186, 635–646. [Google Scholar] [CrossRef]
- Su, Z.; Ma, X.; Guo, H.; Sukiran, N.L.; Guo, B.; Assmann, S.M.; Ma, H. Flower development under drought stress: Morphological and transcriptomic analyses reveal acute responses and long-term acclimation in Arabidopsis. Plant Cell 2013, 25, 3785–3807. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDowell, N.G. Mechanisms linking drought, hydraulics, carbon metabolism, and vegetation mortality. Plant Physiol. 2011, 155, 1051–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huberty, A.F.; Denno, R.F. Plant water stress and its consequences for herbivorous insects: A new synthesis. Ecology 2004, 85, 1383–1398. [Google Scholar] [CrossRef]
- Colinet, H.; Sinclair, B.J.; Vernon, P.; Renault, D. Insects in fluctuating thermal environments. Annu. Rev. Entomol. 2015, 60, 123–140. [Google Scholar] [CrossRef] [Green Version]
- Ma, G.; Rudolf, V.H.; Ma, C.S. Extreme temperature events alter demographic rates, relative fitness, and community structure. Glob. Chang. Biol. 2015, 21, 1794–1808. [Google Scholar] [CrossRef]
- Addo-Bediako, A.; Steven, L.C.; Gaston, K.J. Revisiting water loss in insects: A large-scale view. J. Insect Physiol. 2001, 47, 1377–1388. [Google Scholar] [CrossRef]
- Pritchard, J.; Griffiths, B.; Hunt, E.J. Can the plant-mediated impacts on aphids of elevated CO2 and drought be predicted. Glob. Change Biol. 2007, 13, 1616–1629. [Google Scholar] [CrossRef]
- Sun, L.J.; Chen, J.; Zheng, C. Effect of drought during the root nourishing period of Chinese chives on the population dynamics of the chive gnat, Bradysia odoriphaga (Diptera: Sciaridae). Acta Entomol. Sin. 2019, 62, 79–85. [Google Scholar]
- Wei, Q.; Su, J. Research advances in carbohydrate and lipid metabolism in insects. Acta Entomol. Sin. 2016, 59, 906–916. [Google Scholar]
- Yang, L. A Preliminary Study on Enzymes Involving in Amino Acid Biosynthesis in Two Rice Planthoppers. Master’s Dissertation, Nanjing Agricultural University, Nanjing, China, 2012. [Google Scholar]
- Wen, Z.; Zeng, R.S.; Niu, G. Ecological significance of induction of broad-substrate cytochrome P450s by natural and synthetic inducers in Helicoverpa zea. J. Chem. Ecol. 2009, 35, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Pelz-Stelinski, K.; Stelinski, L.L. Effect of Candidatus Liberibacter asiaticus infection on susceptibility of Asian citrus psyllid, Diaphorina citri, to selected insecticides. Pest Manag. Sci. 2011, 67, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Qian, L.; Ke, R.; Chen, X.; Li, Z.; Gui, F. Effects of elevated carbon dioxide on the activities of physiological enzymes in thrips Frankliniella occidentalis and F. intonsa fed on different host plants. J. Plant Protec. 2017, 1, 45–53. [Google Scholar]
- Zhang, Z.; Cui, B.; Yan, S. Evaluation of tricin, a stylet probing stimulant of brown planthopper, in infested and non-infested rice plants. J. Appl. Entomol. 2017, 141, 393–401. [Google Scholar] [CrossRef]
- Hu, G.; Lu, M.H.; Reynolds, D.R.; Wang, H.K.; Chen, X.; Liu, W.C.; Zhu, F.; Wu, X.W.; Xia, F.; Xie, M.C.; et al. Long-term seasonal forecasting of a major migrant insect pest: The brown planthopper in the Lower Yangtze River Valley. J. Pest Sci. 2019, 92, 417–428. [Google Scholar] [CrossRef] [Green Version]
- Jones, C.M.; Parry, H.; Tay, W.T.; Reynolds, D.R.; Chapman, J.W. Movement ecology of pest Helicoverpa: Implications for ongoing spread. Ann. Rev. Entomol. 2019, 64, 277–295. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.L.; Fu, X.W.; Zhao, S.Y.; Shen, X.J.; Wyckhuys, K.A.G.; Wu, K.M. Long-term shifts in abundance of (migratory) crop-feeding and beneficial insect species in northeastern Asia. J. Pest Sci. 2020, 93, 583–594. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Yin, J.; Cao, Y.; Li, K. The effect of feeding and mating on the development of flight muscle in Agrotis ypsilon. Chin. J. Appl. Entomol. 2013, 50, 1573–1585. [Google Scholar]
- Newmark, P.A.; Boswell, R.E. The mago nashi locus encodes an essential product required for germ plasm assembly in Drosophila. Development 1994, 120, 303–1313. [Google Scholar] [CrossRef] [PubMed]
- Cha, B.J.; Koppetsch, B.S.; Theurkauf, W.E. In vivo analysis of Drosophila bicoidm RNA localization reveals a novelmicrotubule-dependent axis specification pathway. Cell 2001, 106, 35–46. [Google Scholar] [CrossRef] [Green Version]
- Parkash, R.; Lambhod, C.; Singh, D. Ecological significance of wing spot dimorphism in Drosophila biarmipes (Diptera, Drosophilidae). Acta Entomol. Sin. 2013, 56, 1267–1274. [Google Scholar]
- Saffman, E.; Lasko, P. Germline development in vertebrates and invertebrates. Cell. Mol. Life Sci. 1999, 55, 1141–1163. [Google Scholar] [CrossRef] [PubMed]
- Raikhel, A.S.; Dhadialla, T. Accumulation of yolk proteins in insect oocytes. Ann. Rev. Entomol. 1992, 37, 217–251. [Google Scholar] [CrossRef]
- Hagedorn, H.H.; Maddison, D.R.; Tu, Z. The evolution of vitellogenins, cyclorrhaphan yolk proteins and related molecules. Adv. Insect Physiol. 1998, 27, 335–384. [Google Scholar]
- Sappington, T.W.; Raikhel, A.S. Molecular characteristics of insect vitellogenins and vitellogenin receptors. Insect Biochem. Mol. Biol. 1998, 28, 277–300. [Google Scholar] [CrossRef]
- Giorgi, F.; Bradley, J.; Nordin, J. Differential vitellin polypeptide processing in insect embryos. Micron 1999, 30, 579–596. [Google Scholar] [CrossRef]
- Guidugli, K.R.; Nascimento, A.M.; Amdam, G.V.; Barchuk, A.R.; Omholt, S.; Simoes, Z.L.P.; Hartfelder, K. Vitellogenin regulates hormonal dynamics in the worker caste of a eusocial insect. FEBS Lett. 2005, 579, 4961–4965. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Shang, K.; Liu, J.; Jiang, T.; Hu, D.; Hua, H. Multi-generational effects of rice harboring Bph15 on brown planthopper, Nilaparvata lugens. Pest Manag. Sci. 2014, 70, 310–317. [Google Scholar] [CrossRef]
- Tan, Y. Effects of Water Stress on Feeding Behavior and Physiology of Nilaparvata lugens. Master’s Dissertation, Jiliang University, Hangzhou, China, 2014. [Google Scholar]
- An, N.T.T.; Michaud, D.; Cloutier, C. Proteomic profiling of aphid Macrosiphum euphorbiae responses to host-plant-mediated stress induced by defoliation and water deficit. J. Insect Physiol. 2007, 53, 601–611. [Google Scholar]
- Ge, L.Q.; Gu, H.T.; Huang, B.; Song, Q.S.; Stanley, D.; Liu, F.; Yang, G.Q.; Wu, J.C. An adenylyl cyclase like-9 gene (NlAC9) influences growth and fecundity in the brown planthopper, Nilaparvata lugens (Stål) (Hemiptera: Delphacidae). PLoS ONE 2017, 12, e0189214. [Google Scholar] [CrossRef] [PubMed]
- Karley, A.J.; Ashford, D.A.; Minto, L.M.; Pritchard, J.; Douglas, A.E. The significance of gut sucrase activity for osmoregulation in the pea aphid, Acyrthosiphon pisum. J. Insect Physiol. 2005, 51, 1313–1319. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.G.; Jiang, C.X.; Mao, M.; Liu, C.; Li, Q.; Wang, X.G.; Yang, Q.F.; Wang, H.J. Effect of the insecticide Dinotefuran on the ultrastructure of the flight muscle of female Sogatella furcifera (Hemiptera: Delphacidae). J. Econ. Entomol. 2017, 2, 632. [Google Scholar] [CrossRef]
- Wan, D.J.; Chen, J.; Jiang, L.B.; Ge, L.Q.; Wu, J.C. Effects of the insecticide triazophos on the ultrastructure of the flight muscle of the brown planthopper Nilaparvata lugens Stål (Hemiptera: Delphacidae). Crop Protec. 2013, 43, 54–59. [Google Scholar] [CrossRef]
- Steven, L.C.; Jesper, G.S.; John, S.T. Water loss in insects: An environmental change perspective. J. Insect Physiol. 2011, 57, 1070–1084. [Google Scholar]
- Rani, P.U.; Prasannalaxmi, K. Water stress induced physiological and biochemical changes in Piper betle L. and Ricinus communis L. plants and their effects on Spodoptera litura. Allelopath. J. 2014, 33, 25–41. [Google Scholar]
- Luo, D.; Yang, Y.; Xu, H.; Zheng, X.; Lv, Z. Minireview on the influences of drought stress on herbivores and their natural enemies. J. Environ. Entomol. 2012, 34, 229–235. [Google Scholar]
- Wu, J.L. Physiological Response to Drought Stress and Effect on Biodiversity of Key Diseases and Insects in Transgenic Drought-Resistant Soybean. Master’s Dissertation, Collage of Northeast Agricultural University, Harbin, China, 2015. [Google Scholar]
- Simpson, K.L.S.; Jackson, G.E.; Grace, J. The response of aphids to plant water stress–the case of Myzus persicae and Brassica oleracea var. capitata. Entomol. Exp. Appl. 2012, 142, 191–202. [Google Scholar] [CrossRef] [Green Version]
- Maron, J.L. Plant water stress and previous herbivore damage affect insect performance. Ecol. Entomol. 2018, 43, 47–54. [Google Scholar]
- Suárez-Vidal, E.; Sampedro, L.; Voltas, J.; Serrano, L.; Notivol, E.; Zas, R. Drought stress modifies early effective resistance and induced chemical defences of Aleppo pine against a chewing insect herbivore. Environ. Exp. Bot. 2019, 162, 550–559. [Google Scholar] [CrossRef]
- Lenhart, P.A.; Eubanks, M.D.; Behmer, S.T. Water stress in grasslands: Dynamic responses of plants and insect herbivores. Oikos 2015, 124, 381–390. [Google Scholar] [CrossRef]
- Dang, Z.; Chen, F. Responses of insects to rainfall and drought. Acta Entomol. Sin. 2011, 48, 1161–1169. [Google Scholar]
- Yin, X.W. Responses of leaf nitrogen concentration and specific leaf area to atmospheric CO2 enrichment: A retrospective synthesis across 62 species. Glob. Chang. Biol. 2002, 8, 631–642. [Google Scholar] [CrossRef]
- Caldeira, M.C.; Fernandéz, V.; Tomé, J.; Pereira, J.S. Positive effect of drought on longicorn borer larval survival and growth on eucalyptus trunks. Ann. For. Sci. 2002, 59, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; He, L.; Liu, H.; Yan, R.; Wan, Q. The changes of amino acid, SOD and CAT of Musca domestica larva during diapause. Sichuan J. Zool. 2009, 4, 105–107. [Google Scholar]
- Moheb, A.; Grondin, M.; Ibrahim, R.K. Winter wheat hull (husk) is a valuable source for tricin, a potential selective cytotoxic agent. Food Chem. 2013, 138, 931–937. [Google Scholar] [CrossRef] [PubMed]
- Gong, G.; Cui, B.; Dai, Z.; Bei, J.; Li, Y.; Zhang, Y.; Wu, W.; Zhang, Z. Salivary proteomics response of rice brown planthopper Nilaparvata lugens (Stål) to stimulation of tricin, a key rice insect-resistant compound. J. Environ. Entomol. 2019, 41, 50–61. [Google Scholar]
- Chen, X.; Wang, Z.; Li, X.; Zhu, Y.; Liu, L.; Chen, W.; Chen, Q. Research Progress on Glutathione S-Transferases; Northeast Agricultural University: Harbin, China, 2013; Volume 1, pp. 155–159. [Google Scholar]
- Bao, Y.X.; Huang, J.Y.; Xie, X.J.; Lu, M.H. Influence of monsoon’s advancing, retreating and conversion on migrations of Nilaparvata lugens (Stål) in China. Acta Ecol. Sin. 2013, 33, 4864–4877. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequences (5′–3′) | GenBank Accession No. |
|---|---|---|
| NlActin-qF | TGGACTTCGAGCAGGAAATGG | EU179846.1 |
| NlActin-qR | ACGTCGCACTTCAGATCGAG | |
| NlVg-qF | GTGGCTCGTTCAAGGTTATGG | AB353856 |
| NlVg-qR | GCAATCTCTGGGTGCTGTTG | |
| NlExu-qF | GGGCGCTCAGGGATAAGACT | DB823987 |
| NlExu-qR | GGGCATCATGACAAAGCAGAA | |
| NlMag-qF | CCGGATGGCAAGTTGAGGTA | DB843362 |
| NlMag-qR | TGTGGCCAAGGTGAGTCATCT | |
| NlVas-qF | CCGATGCGGTGGATGTACTC | CG3056 |
| NlVas-qR | GGCGCTGCATCTCTTCAAGT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, X.; Chen, L.; Lan, X.; Liao, G.; Feng, L.; Li, J.; Fan, W.; Wang, S.; Liu, J. Physiological and Population Responses of Nilaparvata lugens after Feeding on Drought-Stressed Rice. Insects 2022, 13, 355. https://doi.org/10.3390/insects13040355
Liang X, Chen L, Lan X, Liao G, Feng L, Li J, Fan W, Wang S, Liu J. Physiological and Population Responses of Nilaparvata lugens after Feeding on Drought-Stressed Rice. Insects. 2022; 13(4):355. https://doi.org/10.3390/insects13040355
Chicago/Turabian StyleLiang, Xinyan, Lin Chen, Xiaoying Lan, Guangrong Liao, Ling Feng, Jitong Li, Wenyan Fan, Shuang Wang, and Jinglan Liu. 2022. "Physiological and Population Responses of Nilaparvata lugens after Feeding on Drought-Stressed Rice" Insects 13, no. 4: 355. https://doi.org/10.3390/insects13040355
APA StyleLiang, X., Chen, L., Lan, X., Liao, G., Feng, L., Li, J., Fan, W., Wang, S., & Liu, J. (2022). Physiological and Population Responses of Nilaparvata lugens after Feeding on Drought-Stressed Rice. Insects, 13(4), 355. https://doi.org/10.3390/insects13040355