
Time-Dependent Odorant Sensitivity Modulation in Insects


{kind=link}
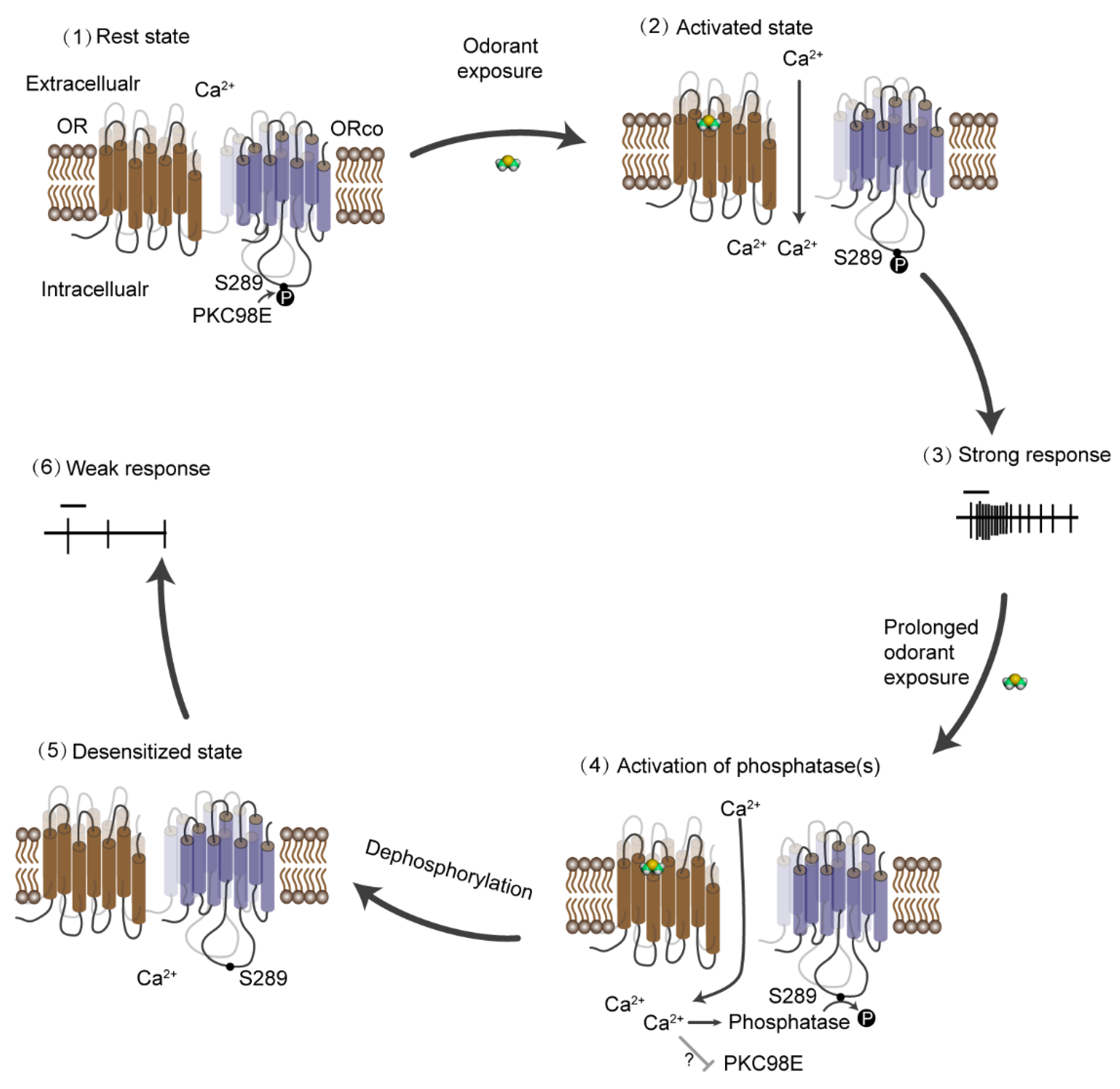{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Desensitization of ORNs
2.1. Short-Term Desensitization
2.2. Long-Term Desensitization
3. Molecular Mechanism of Olfactory Desensitization
3.1. Short-Term Desensitization
3.2. Long-Term Desensitization
4. Future Directions
4.1. Factors Activating PKC98E
4.2. Phosphatase Dephosphorylating Orco S289
4.3. Molecular Mechanism of Desensitization of ORNs
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cardé, R.T. Navigation Along Windborne Plumes of Pheromone and Resource-Linked Odors. Annu. Rev. Entomol. 2021, 66, 317–336. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Wang, C.-Z. The Ethological Significance and Olfactory Detection of Herbivore-Induced Plant Volatiles in Interactions of Plants, Herbivorous Insects, and Parasitoids. Arthropod-Plant Interact. 2019, 13, 161–179. [Google Scholar] [CrossRef]
- Bruce, T.J.A.; Wadhams, L.J.; Woodcock, C.M. Insect Host Location: A Volatile Situation. Trends Plant Sci. 2005, 10, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Hansson, B.S.; Stensmyr, M.C. Evolution of Insect Olfaction. Neuron 2011, 72, 698–711. [Google Scholar] [CrossRef] [Green Version]
- Kaissling, K.-E. The Sensitivity of the Insect Nose: The Example of Bombyx Mori. In Biologically Inspired Signal Processing for Chemical Sensing; Gutiérrez, A., Marco, S., Eds.; Studies in Computational Intelligence; Springer: Berlin/Heidelberg, Germany, 2009; pp. 45–52. ISBN 978-3-642-00176-5. [Google Scholar]
- Carey, A.F.; Carlson, J.R. Insect Olfaction from Model Systems to Disease Control. Proc. Natl. Acad. Sci. USA 2011, 108, 12987–12995. [Google Scholar] [CrossRef] [Green Version]
- Stocker, R.F. The Organization of the Chemosensory System in Drosophila melanogaster: A Rewiew. Cell Tissue Res. 1994, 275, 3–26. [Google Scholar] [CrossRef]
- Hallem, E.A.; Carlson, J.R. The Odor Coding System of Drosophila. Trends Genet. 2004, 20, 453–459. [Google Scholar] [CrossRef]
- Vosshall, L.B.; Stocker, R.F. Molecular Architecture of Smell and Taste in Drosophila. Annu. Rev. Neurosci. 2007, 30, 505–533. [Google Scholar] [CrossRef] [Green Version]
- Leal, W.S. Odorant Reception in Insects: Roles of Receptors, Binding Proteins, and Degrading Enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef]
- Ronderos, D.; Smith, D. Diverse Signaling Mechanisms Mediate Volatile Odorant Detection in Drosophila. Fly 2009, 3, 290–297. [Google Scholar] [CrossRef] [Green Version]
- Joseph, R.M.; Carlson, J.R. Drosophila Chemoreceptors: A Molecular Interface Between the Chemical World and the Brain. Trends Genet. 2015, 31, 683–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, C.-Y.; Menuz, K.; Carlson, J.R. Olfactory Perception: Receptors, Cells, and Circuits. Cell 2009, 139, 45–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakagawa, T.; Vosshall, L.B. Controversy and Consensus: Noncanonical Signaling Mechanisms in the Insect Olfactory System. Curr. Opin. Neurobiol. 2009, 19, 284–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallem, E.A.; Ho, M.G.; Carlson, J.R. The Molecular Basis of Odor Coding in the Drosophila Antenna. Cell 2004, 117, 965–979. [Google Scholar] [CrossRef] [Green Version]
- Xiao, S.; Sun, J.S.; Carlson, J.R. Robust Olfactory Responses in the Absence of Odorant Binding Proteins. eLife 2019, 8, e51040. [Google Scholar] [CrossRef]
- Ronderos, D.S.; Lin, C.-C.; Potter, C.J.; Smith, D.P. Farnesol-Detecting Olfactory Neurons in Drosophila. J. Neurosci. 2014, 34, 3959–3968. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.; Atkinson, R.; Jones, D.N.M.; Smith, D.P. Drosophila OBP LUSH Is Required for Activity of Pheromone-Sensitive Neurons. Neuron 2005, 45, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Haverkamp, A.; Hansson, B.S.; Knaden, M. Combinatorial Codes and Labeled Lines: How Insects Use Olfactory Cues to Find and Judge Food, Mates, and Oviposition Sites in Complex Environments. Front. Physiol. 2018, 9, 49. [Google Scholar] [CrossRef]
- Kurtovic, A.; Widmer, A.; Dickson, B.J. A Single Class of Olfactory Neurons Mediates Behavioural Responses to a Drosophila Sex Pheromone. Nature 2007, 446, 542–546. [Google Scholar] [CrossRef]
- Ha, T.S.; Smith, D.P. A Pheromone Receptor Mediates 11-Cis-Vaccenyl Acetate-Induced Responses in Drosophila. J. Neurosci. 2006, 26, 8727–8733. [Google Scholar] [CrossRef]
- Stensmyr, M.C.; Dweck, H.K.M.; Farhan, A.; Ibba, I.; Strutz, A.; Mukunda, L.; Linz, J.; Grabe, V.; Steck, K.; Lavista-Llanos, S.; et al. A Conserved Dedicated Olfactory Circuit for Detecting Harmful Microbes in Drosophila. Cell 2012, 151, 1345–1357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelosi, P. Odorant-Binding Proteins. Crit. Rev. Biochem. Mol. Biol. 1994, 29, 199–228. [Google Scholar] [CrossRef] [PubMed]
- Klein, U. Sensillum-Lymph Proteins from Antennal Olfactory Hairs of the Moth Antheraea Polyphemus (Saturniidae). Insect Biochem. 1987, 17, 1193–1204. [Google Scholar] [CrossRef]
- Vogt, R.G.; Riddiford, L.M. Pheromone Binding and Inactivation by Moth Antennae. Nature 1981, 293, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Guo, P.-P.; Sun, Y.-L.; Huang, L.-Q.; Wang, C.-Z. Contribution of Odorant Binding Proteins to Olfactory Detection of (Z)-11-Hexadecenal in Helicoverpa armigera. Insect Biochem. Mol. Biol. 2021, 131, 103554. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Huang, L.-Q.; Pelosi, P.; Wang, C.-Z. Three Pheromone-Binding Proteins Help Segregation between Two Helicoverpa Species Utilizing the Same Pheromone Components. Insect Biochem. Mol. Biol. 2012, 42, 708–716. [Google Scholar] [CrossRef]
- Pelosi, P.; Zhou, J.-J.; Ban, L.P.; Calvello, M. Soluble Proteins in Insect Chemical Communication. Cell. Mol. Life Sci. 2006, 63, 1658–1676. [Google Scholar] [CrossRef]
- Laughlin, J.D.; Ha, T.S.; Jones, D.N.M.; Smith, D.P. Activation of Pheromone-Sensitive Neurons Is Mediated by Conformational Activation of Pheromone-Binding Protein. Cell 2008, 133, 1255–1265. [Google Scholar] [CrossRef] [Green Version]
- Sun, J.S.; Xiao, S.; Carlson, J.R. The Diverse Small Proteins Called Odorant-Binding Proteins. Open Biol. 2018, 8, 180208. [Google Scholar] [CrossRef] [Green Version]
- Syed, Z.; Ishida, Y.; Taylor, K.; Kimbrell, D.A.; Leal, W.S. Pheromone Reception in Fruit Flies Expressing a Moth’s Odorant Receptor. Proc. Natl. Acad. Sci. USA 2006, 103, 16538–16543. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.-H.; Zheng, M.-Y.; Sun, J.-B.; Khuhro, S.A.; Yan, Q.; Huang, Y.; Syed, Z.; Dong, S.-L. CRISPR/Cas9 Mediated Gene Knockout Reveals a More Important Role of PBP1 than PBP2 in the Perception of Female Sex Pheromone Components in Spodoptera litura. Insect Biochem. Mol. Biol. 2019, 115, 103244. [Google Scholar] [CrossRef] [PubMed]
- Larter, N.K.; Sun, J.S.; Carlson, J.R. Organization and Function of Drosophila Odorant Binding Proteins. eLife 2016, 5, e20242. [Google Scholar] [CrossRef] [PubMed]
- Scheuermann, E.A.; Smith, D.P. Odor-Specific Deactivation Defects in a Drosophila Odorant-Binding Protein Mutant. Genetics 2019, 213, 897–909. [Google Scholar] [CrossRef] [PubMed]
- Vosshall, L.B.; Amrein, H.; Morozov, P.S.; Rzhetsky, A.; Axel, R. A Spatial Map of Olfactory Receptor Expression in the Drosophila Antenna. Cell 1999, 96, 725–736. [Google Scholar] [CrossRef] [Green Version]
- Gao, Q.; Chess, A. Identification of Candidate Drosophila Olfactory Receptors from Genomic DNA Sequence. Genomics 1999, 60, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Clyne, P.J.; Warr, C.G.; Freeman, M.R.; Lessing, D.; Kim, J.; Carlson, J.R. A Novel Family of Divergent Seven-Transmembrane Proteins: Candidate Odorant Receptors in Drosophila. Neuron 1999, 22, 327–338. [Google Scholar] [CrossRef] [Green Version]
- Benton, R.; Vannice, K.S.; Gomez-Diaz, C.; Vosshall, L.B. Variant Ionotropic Glutamate Receptors as Chemosensory Receptors in Drosophila. Cell 2009, 136, 149–162. [Google Scholar] [CrossRef] [Green Version]
- Kwon, J.Y.; Dahanukar, A.; Weiss, L.A.; Carlson, J.R. The Molecular Basis of CO2 Reception in Drosophila. Proc. Natl. Acad. Sci. USA 2007, 104, 3574–3578. [Google Scholar] [CrossRef] [Green Version]
- Jones, W.D.; Cayirlioglu, P.; Grunwald Kadow, I.; Vosshall, L.B. Two Chemosensory Receptors Together Mediate Carbon Dioxide Detection in Drosophila. Nature 2007, 445, 86–90. [Google Scholar] [CrossRef]
- de Bruyne, M.; Foster, K.; Carlson, J.R. Odor Coding in the Drosophila Antenna. Neuron 2001, 30, 537–552. [Google Scholar] [CrossRef] [Green Version]
- Dobritsa, A.A.; van Naters, W.V.D.G.; Warr, C.G.; Steinbrecht, R.A.; Carlson, J.R. Integrating the Molecular and Cellular Basis of Odor Coding in the Drosophila Antenna. Neuron 2003, 37, 827–841. [Google Scholar] [CrossRef] [Green Version]
- Yao, C.A.; Ignell, R.; Carlson, J.R. Chemosensory Coding by Neurons in the Coeloconic Sensilla of the Drosophila Antenna. J. Neurosci. 2005, 25, 8359–8367. [Google Scholar] [CrossRef] [PubMed]
- Buck, L.; Axel, R. A Novel Multigene Family May Encode Odorant Receptors: A Molecular Basis for Odor Recognition. Cell 1991, 65, 175–187. [Google Scholar] [CrossRef]
- Troemel, E.R.; Chou, J.H.; Dwyer, N.D.; Colbert, H.A.; Bargmann, C.I. Divergent Seven Transmembrane Receptors Are Candidate Chemosensory Receptors in C. elegans. Cell 1995, 83, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Lundin, C.; Käll, L.; Kreher, S.A.; Kapp, K.; Sonnhammer, E.L.; Carlson, J.R.; von Heijne, G.; Nilsson, I. Membrane Topology of the Drosophila OR83b Odorant Receptor. FEBS Lett. 2007, 581, 5601–5604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benton, R.; Sachse, S.; Michnick, S.W.; Vosshall, L.B. Atypical Membrane Topology and Heteromeric Function of Drosophila Odorant Receptors in vivo. PLoS Biol. 2006, 4, e20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smart, R.; Kiely, A.; Beale, M.; Vargas, E.; Carraher, C.; Kralicek, A.V.; Christie, D.L.; Chen, C.; Newcomb, R.D.; Warr, C.G. Drosophila Odorant Receptors Are Novel Seven Transmembrane Domain Proteins That Can Signal Independently of Heterotrimeric G Proteins. Insect Biochem. Mol. Biol. 2008, 38, 770–780. [Google Scholar] [CrossRef]
- Kaupp, U.B. Olfactory Signalling in Vertebrates and Insects: Differences and Commonalities. Nat. Rev. Neurosci. 2010, 11, 188–200. [Google Scholar] [CrossRef]
- Task, D.; Lin, C.-C.; Vulpe, A.; Afify, A.; Ballou, S.; Brbić, M.; Schlegel, P.; Jefferis, G.S.X.E.; Li, H.; Menuz, K.; et al. Chemoreceptor Co-Expression in Drosophila Olfactory Neurons. bioRxiv 2021, 11, 355651. [Google Scholar]
- Kumar, A.; Tauxe, G.M.; Perry, S.; Scott, C.A.; Dahanukar, A.; Ray, A. Contributions of the Conserved Insect Carbon Dioxide Receptor Subunits to Odor Detection. Cell Rep. 2020, 31, 107510. [Google Scholar] [CrossRef]
- Ning, C.; Yang, K.; Xu, M.; Huang, L.-Q.; Wang, C.-Z. Functional Validation of the Carbon Dioxide Receptor in Labial Palps of Helicoverpa Armigera Moths. Insect Biochem. Mol. Biol. 2016, 73, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Pellegrino, M.; Nakagawa, T.; Nakagawa, T.; Vosshall, L.B.; Touhara, K. Insect Olfactory Receptors Are Heteromeric Ligand-Gated Ion Channels. Nature 2008, 452, 1002–1006. [Google Scholar] [CrossRef] [PubMed]
- Wicher, D.; Schäfer, R.; Bauernfeind, R.; Stensmyr, M.C.; Heller, R.; Heinemann, S.H.; Hansson, B.S. Drosophila Odorant Receptors Are Both Ligand-Gated and Cyclic-Nucleotide-Activated Cation Channels. Nature 2008, 452, 1007–1011. [Google Scholar] [CrossRef] [PubMed]
- Neuhaus, E.M.; Gisselmann, G.; Zhang, W.; Dooley, R.; Störtkuhl, K.; Hatt, H. Odorant Receptor Heterodimerization in the Olfactory System of Drosophila melanogaster. Nat. Neurosci. 2005, 8, 15–17. [Google Scholar] [CrossRef]
- Yao, C.A.; Carlson, J.R. Role of G-Proteins in Odor-Sensing and CO2-Sensing Neurons in Drosophila. J. Neurosci. 2010, 30, 4562–4572. [Google Scholar] [CrossRef] [PubMed]
- Kain, P.; Chakraborty, T.S.; Sundaram, S.; Siddiqi, O.; Rodrigues, V.; Hasan, G. Reduced Odor Responses from Antennal Neurons of Gqα, Phospholipase Cβ, and RdgA Mutants in Drosophila Support a Role for a Phospholipid Intermediate in Insect Olfactory Transduction. J. Neurosci. 2008, 28, 4745–4755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breer, H.; Boekhoff, I.; Tareilus, E. Rapid Kinetics of Second Messenger Formation in Olfactory Transduction. Nature 1990, 345, 65–68. [Google Scholar] [CrossRef]
- Boekhoff, I.; Raming, K.; Breer, H. Pheromone-Induced Stimulation of Inositol-Trisphosphate Formation in Insect Antennae Is Mediated by G-Proteins. J. Comp. Physiol. B 1990, 160, 99–103. [Google Scholar] [CrossRef]
- Boekhoff, I.; Strotmann, J.; Raming, K.; Tareilus, E.; Breer, H. Odorant-Sensitive Phospholipase C in Insect Antennae. Cell Signal. 1990, 2, 49–56. [Google Scholar] [CrossRef]
- Gomez-Diaz, C.; Martin, F.; Alcorta, E. The CAMP Transduction Cascade Mediates Olfactory Reception in Drosophila melanogaster. Behav. Genet. 2004, 34, 395–406. [Google Scholar] [CrossRef]
- Miazzi, F.; Hansson, B.S.; Wicher, D. Odor-Induced CAMP Production in Drosophila melanogaster Olfactory Sensory Neurons. J. Exp. Biol. 2016, 219, 1798–1803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziegelberger, G.; van den, B.M.; Kaissling, K.E.; Klumpp, S.; Schultz, J.E. Cyclic GMP Levels and Guanylate Cyclase Activity in Pheromone-Sensitive Antennae of the Silkmoths Antheraea polyphemus and Bombyx mori. J. Neurosci. 1990, 10, 1217–1225. [Google Scholar] [CrossRef] [PubMed]
- Boekhoff, I.; Seifert, E.; Göggerle, S.; Lindemann, M.; Krüger, B.-W.; Breer, H. Pheromone-Induced Second-Messenger Signaling in Insect Antennae. Insect Biochem. Mol. Biol. 1993, 23, 757–762. [Google Scholar] [CrossRef]
- Butterwick, J.A.; del Mármol, J.; Kim, K.H.; Kahlson, M.A.; Rogow, J.A.; Walz, T.; Ruta, V. Cryo-EM Structure of the Insect Olfactory Receptor Orco. Nature 2018, 560, 447–452. [Google Scholar] [CrossRef]
- del Mármol, J.; Yedlin, M.A.; Ruta, V. The Structural Basis of Odorant Recognition in Insect Olfactory Receptors. Nature 2021, 597, 126–131. [Google Scholar] [CrossRef]
- Brand, P.; Robertson, H.M.; Lin, W.; Pothula, R.; Klingeman, W.E.; Jurat-Fuentes, J.L.; Johnson, B.R. The Origin of the Odorant Receptor Gene Family in Insects. eLife 2018, 7, e38340. [Google Scholar] [CrossRef]
- McKenzie, S.K.; Kronauer, D.J.C. The Genomic Architecture and Molecular Evolution of Ant Odorant Receptors. Genome Res. 2018, 28, 1757–1765. [Google Scholar] [CrossRef] [Green Version]
- Robertson, H.M.; Wanner, K.W. The Chemoreceptor Superfamily in the Honey Bee, Apis Mellifera: Expansion of the Odorant, but Not Gustatory, Receptor Family. Genome Res. 2006, 16, 1395–1403. [Google Scholar] [CrossRef] [Green Version]
- Jones, W.D.; Nguyen, T.-A.T.; Kloss, B.; Lee, K.J.; Vosshall, L.B. Functional Conservation of an Insect Odorant Receptor Gene across 250 Million Years of Evolution. Curr. Biol. 2005, 15, R119–R121. [Google Scholar] [CrossRef] [Green Version]
- Larsson, M.C.; Domingos, A.I.; Jones, W.D.; Chiappe, M.E.; Amrein, H.; Vosshall, L.B. Or83b Encodes a Broadly Expressed Odorant Receptor Essential for Drosophila Olfaction. Neuron 2004, 43, 703–714. [Google Scholar] [CrossRef] [Green Version]
- Stengl, M.; Funk, N.W. The Role of the Coreceptor Orco in Insect Olfactory Transduction. J. Comp. Physiol. A 2013, 199, 897–909. [Google Scholar] [CrossRef] [PubMed]
- Cardé, R.T.; Willis, M.A. Navigational Strategies Used by Insects to Find Distant, Wind-Borne Sources of Odor. J. Chem. Ecol. 2008, 34, 854–866. [Google Scholar] [CrossRef] [PubMed]
- Conchou, L.; Lucas, P.; Meslin, C.; Proffit, M.; Staudt, M.; Renou, M. Insect Odorscapes: From Plant Volatiles to Natural Olfactory Scenes. Front. Physiol. 2019, 10, 972. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.I. Early Olfactory Processing in Drosophila: Mechanisms and Principles. Annu. Rev. Neurosci. 2013, 36, 217–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaissling, K.-E.; Strausfeld, C.Z.; Rumbo, E.R. Adaptation Processes in Insect Olfactory Receptors. Ann. N. Y. Acad. Sci. 1987, 510, 104–112. [Google Scholar] [CrossRef]
- Guo, H.; Smith, D.P. Odorant Receptor Desensitization in Insects. J. Exp. Neurosci. 2017, 11, 1179069517748600. [Google Scholar] [CrossRef]
- Zufall, F.; Leinders-Zufall, T. The Cellular and Molecular Basis of Odor Adaptation. Chem. Senses 2000, 25, 473–481. [Google Scholar] [CrossRef]
- Guo, H.; Kunwar, K.; Smith, D. Odorant Receptor Sensitivity Modulation in Drosophila. J. Neurosci. 2017, 37, 9465–9473. [Google Scholar] [CrossRef] [Green Version]
- Dolzer, J.; Fischer, K.; Stengl, M. Adaptation in Pheromone-Sensitive Trichoid Sensilla of the Hawkmoth Manduca sexta. J. Exp. Biol. 2003, 206, 1575–1588. [Google Scholar] [CrossRef] [Green Version]
- Gorur-Shandilya, S.; Demir, M.; Long, J.; Clark, D.A.; Emonet, T. Olfactory Receptor Neurons Use Gain Control and Complementary Kinetics to Encode Intermittent Odorant Stimuli. eLife 2017, 6, e27670. [Google Scholar] [CrossRef]
- Murmu, M.S.; Stinnakre, J.; Réal, E.; Martin, J.-R. Calcium-Stores Mediate Adaptation in Axon Terminals of Olfactory Receptor Neurons in Drosophila. BMC Neurosci. 2011, 12, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olsen, S.R.; Wilson, R.I. Lateral Presynaptic Inhibition Mediates Gain Control in an Olfactory Circuit. Nature 2008, 452, 956–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ignell, R.; Root, C.M.; Birse, R.T.; Wang, J.W.; Nässel, D.R.; Winther, Å.M.E. Presynaptic Peptidergic Modulation of Olfactory Receptor Neurons in Drosophila. Proc. Natl. Acad. Sci. USA 2009, 106, 13070–13075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baker, T.C.; Hansson, B.S.; Löfstedt, C.; Löfqvist, J. Adaptation of Antennal Neurons in Moths Is Associated with Cessation of Pheromone-Mediated Upwind Flight. Proc. Natl. Acad. Sci. USA 1988, 85, 9826–9830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Störtkuhl, K.F.; Hovemann, B.T.; Carlson, J.R. Olfactory Adaptation Depends on the Trp Ca2+ Channel in Drosophila. J. Neurosci. 1999, 19, 4839–4846. [Google Scholar] [CrossRef] [Green Version]
- Martelli, C.; Carlson, J.R.; Emonet, T. Intensity Invariant Dynamics and Odor-Specific Latencies in Olfactory Receptor Neuron Response. J. Neurosci. 2013, 33, 6285–6297. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.-H.; Jing, B.-Y.; Yang, D.; Zeng, X.; Shen, Y.; Tu, Y.; Luo, D.-G. Distinct Signaling of Drosophila Chemoreceptors in Olfactory Sensory Neurons. Proc. Natl. Acad. Sci. USA 2016, 113, E902–E911. [Google Scholar] [CrossRef] [Green Version]
- Cobb, M.; Domain, I. Olfactory Coding in a Simple System: Adaptation in Drosophila Larvae. Proc. R. Soc. Lond. B 2000, 267, 2119–2125. [Google Scholar] [CrossRef] [Green Version]
- Deshpande, M.; Venkatesh, K.; Rodrigues, V.; Hasan, G. The Inositol 1,4,5-Trisphosphate Receptor Is Required for Maintenance of Olfactory Adaptation in Drosophila Antennae. J. Neurobiol. 2000, 43, 282–288. [Google Scholar] [CrossRef]
- de Bruyne, M.; Clyne, P.J.; Carlson, J.R. Odor Coding in a Model Olfactory Organ: The Drosophila Maxillary Palp. J. Neurosci. 1999, 19, 4520–4532. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, S.R.; Mohapatra, P.; Nagalla, M.; Sindvani, R.; Amaya, D.; Dickson, H.A.; Menuz, K. Identification and Characterization of CYPs Induced in the Drosophila Antenna by Exposure to a Plant Odorant. Sci. Rep. 2021, 11, 20530. [Google Scholar] [CrossRef]
- Colbert, H.A.; Bargmann, C.I. Odorant-Specific Adaptation Pathways Generate Olfactory Plasticity in C. elegans. Neuron 1995, 14, 803–812. [Google Scholar] [CrossRef] [Green Version]
- Bargmann, C.I.; Hartwieg, E.; Horvitz, H.R. Odorant-Selective Genes and Neurons Mediate Olfaction in C. elegans. Cell 1993, 74, 515–527. [Google Scholar] [CrossRef]
- Stelinski, L.L.; Miller, J.R.; Gut, L.J. Presence of Long-Lasting Peripheral Adaptation in Oblique-Banded Leafroller, Choristoneura Rosaceana and Absence of Such Adaptation in Redbanded Leafroller, Argyrotaenia velutinana. J. Chem. Ecol. 2003, 29, 405–423. [Google Scholar] [CrossRef]
- Stanczyk, N.M.; Brookfield, J.F.Y.; Field, L.M.; Logan, J.G. Aedes aegypti Mosquitoes Exhibit Decreased Repellency by DEET Following Previous Exposure. PLoS ONE 2013, 8, e54438. [Google Scholar] [CrossRef] [PubMed]
- Schild, D.; Restrepo, D. Transduction Mechanisms in Vertebrate Olfactory Receptor Cells. Physiol. Rev. 1998, 78, 429–466. [Google Scholar] [CrossRef] [PubMed]
- Kato, A.; Touhara, K. Mammalian Olfactory Receptors: Pharmacology, G Protein Coupling and Desensitization. Cell. Mol. Life Sci. 2009, 66, 3743. [Google Scholar] [CrossRef]
- Mashukova, A.; Spehr, M.; Hatt, H.; Neuhaus, E.M. β-Arrestin2-Mediated Internalization of Mammalian Odorant Receptors. J. Neurosci. 2006, 26, 9902–9912. [Google Scholar] [CrossRef] [Green Version]
- Nagel, K.I.; Wilson, R.I. Biophysical Mechanisms Underlying Olfactory Receptor Neuron Dynamics. Nat. Neurosci. 2011, 14, 208–216. [Google Scholar] [CrossRef] [Green Version]
- Chertemps, T.; François, A.; Durand, N.; Rosell, G.; Dekker, T.; Lucas, P.; Maïbèche-Coisne, M. A Carboxylesterase, Esterase-6, Modulates Sensory Physiological and Behavioral Response Dynamics to Pheromone in Drosophila. BMC Biol. 2012, 10, 56. [Google Scholar] [CrossRef] [Green Version]
- Sargsyan, V.; Getahun, M.; Lavista Llanos, S.; Olsson, S.; Hansson, B.; Wicher, D. Phosphorylation via PKC Regulates the Function of the Drosophila Odorant Co-Receptor. Front. Cell. Neurosci. 2011, 5, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poudel, S.; Guo, H.; Smith, D.P. PKC98E Regulates Odorant Responses in Drosophila melanogaster. J. Neurosci. 2021, 41, 3948–3957. [Google Scholar] [CrossRef] [PubMed]
- Tremmel, D.M.; Resad, S.; Little, C.J.; Wesley, C.S. Notch and PKC Are Involved in Formation of the Lateral Region of the Dorso-Ventral Axis in Drosophila Embryos. PLoS ONE 2013, 8, e67789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, X.; Ha, T.S.; Smith, D.P. SNMP Is a Signaling Component Required for Pheromone Sensitivity in Drosophila. Proc. Natl. Acad. Sci. USA 2008, 105, 10996–11001. [Google Scholar] [CrossRef] [Green Version]
- Schaeffer, E.; Smith, D.; Mardon, G.; Quinn, W.; Zuker, C. Isolation and Characterization of Two New Drosophila Protein Kinase C Genes, Including One Specifically Expressed in Photoreceptor Cells. Cell 1989, 57, 403–412. [Google Scholar] [CrossRef]
- Johnson, J.E.; Giorgione, J.; Newton, A.C. The C1 and C2 Domains of Protein Kinase C Are Independent Membrane Targeting Modules, with Specificity for Phosphatidylserine Conferred by the C1 Domain. Biochemistry 2000, 39, 11360–11369. [Google Scholar] [CrossRef]
- Steinberg, S.F. Structural Basis of Protein Kinase C Isoform Function. Physiol. Rev. 2008, 88, 1341–1378. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-C.; Pearce, M.W.; Honda, T.; Johnson, T.K.; Charlu, S.; Sharma, K.R.; Imad, M.; Burke, R.E.; Zinsmaier, K.E.; Ray, A.; et al. The Drosophila melanogaster Phospholipid Flippase DATP8B Is Required for Odorant Receptor Function. PLoS Genet. 2014, 10, e1004209. [Google Scholar] [CrossRef]
- Ha, T.S.; Xia, R.; Zhang, H.; Jin, X.; Smith, D.P. Lipid Flippase Modulates Olfactory Receptor Expression and Odorant Sensitivity in Drosophila. Proc. Natl. Acad. Sci. USA 2014, 111, 7831–7836. [Google Scholar] [CrossRef] [Green Version]
- Getahun, M.N.; Olsson, S.B.; Lavista-Llanos, S.; Hansson, B.S.; Wicher, D. Insect Odorant Response Sensitivity Is Tuned by Metabotropically Autoregulated Olfactory Receptors. PLoS ONE 2013, 8, e58889. [Google Scholar] [CrossRef] [Green Version]
- Jain, K.; Lavista-Llanos, S.; Grabe, V.; Hansson, B.S.; Wicher, D. Calmodulin Regulates the Olfactory Performance in Drosophila melanogaster. Sci. Rep. 2021, 11, 3747. [Google Scholar] [CrossRef] [PubMed]
- Bahk, S.; Jones, W.D. Insect Odorant Receptor Trafficking Requires Calmodulin. BMC Biol. 2016, 14, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, H.; Smith, D.P. Time-Dependent Odorant Sensitivity Modulation in Insects. Insects 2022, 13, 354. https://doi.org/10.3390/insects13040354
Guo H, Smith DP. Time-Dependent Odorant Sensitivity Modulation in Insects. Insects. 2022; 13(4):354. https://doi.org/10.3390/insects13040354
Chicago/Turabian StyleGuo, Hao, and Dean P. Smith. 2022. "Time-Dependent Odorant Sensitivity Modulation in Insects" Insects 13, no. 4: 354. https://doi.org/10.3390/insects13040354
APA StyleGuo, H., & Smith, D. P. (2022). Time-Dependent Odorant Sensitivity Modulation in Insects. Insects, 13(4), 354. https://doi.org/10.3390/insects13040354