
Influence of Zwitterionic Buffer Effects with Thermal Modification Treatments of Wood on Symbiotic Protists in Reticulitermes grassei Clément





Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Wood Treatment
2.2. Effects on Subterranean Termites
2.3. Scanning Electron Microscopy (SEM)
2.4. Effects on Symbiotic Protists
2.5. Statistical Analysis
3. Results
3.1. Efficacy against Subterranean Termites
3.2. Effects on Symbiotic Protists
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wright, M.S.; Lax, A.R. Combined effect of microbial and chemical control agents on subterranean termites. J. Microbiol. 2013, 51, 578–583. [Google Scholar] [CrossRef]
- Unsal, O.; Kartal, S.N.; Candan, Z.; Arango, R.A.; Clausen, C.A.; Green, F., III. Decay and termite resistance, water absorption and swelling of thermally compressed wood panels. Int. Biodeter. Biodegrad. 2009, 63, 548–552. [Google Scholar] [CrossRef]
- Oliver-Villanueva, J.; Gascón-Garrido, P.; Ibiza-Palacios, M.S. Evaluation of thermally-treated wood of beech (Fagus sylvatica L.) and ash (Fraxinus excelsior L.) against Mediterranean termites (Reticulitermes spp.). Eur. J. Wood Wood Prod. 2013, 71, 391–393. [Google Scholar] [CrossRef]
- Brito, F.M.S.; Paes, J.B.; Oliveira, J.T.S.; Arantes, M.D.C.; Dudecki, L. Chemical characterization and biological resistance of thermally treated bamboo. Constr. Build. Mater. 2020, 262, 120033. [Google Scholar] [CrossRef]
- Salman, S.; Thévenon, M.F.; Pétrissans, A.; Dumarçay, S.; Candelier, K.; Gérardin, P. Improvement of the durability of heat-treated wood against termites. Maderas-Cienc. Tecnol. 2017, 19, 317–328. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.; Sandberg, D.; Goli, G.; Todaro, L. Wood Modification in Europe: A State-of-the-Art about Processes, Products and Applications; Firenze University Press: Florence, Italy, 2019; 113p, ISBN 978-88-6453-970-6. (online PDF). [Google Scholar]
- Sandberg, D.; Kutnar, A.; Karlsson, O.; Jones, D. Wood Modification Technologies: Principles, Sustainability, and the Need for Innovation; Taylor and Francis Ltd.: Abingdon, UK, 2021; 450p, ISBN 9781138491779. [Google Scholar]
- Spear, M.; Binding, T.; Jenkins, D.; Nicholls, J.; Ormondroyd, G. Mild thermal modification to enhance the machinability of larch. In Proceedings of the 7th European Conference on Wood Modification, Lisbon, Portugal, 10–12 March 2014. [Google Scholar]
- Zhang, J.-W.; Liu, H.-H.; Yang, L.; Han, T.-Q.; Yin, Q. Effect of moderate temperature thermal modification combined with wax impregnation on wood properties. Appl. Sci. 2020, 10, 8231. [Google Scholar] [CrossRef]
- Humar, M.; Kržišnik, D.; Lesar, B.; Thaler, N.; Ugovšek, A.; Zupančič, K.; Žlahtič, M. Thermal modification of wax-impregnated wood to enhance its physical, mechanical, and biological properties. Holzforschung 2017, 71, 57–64. [Google Scholar] [CrossRef]
- Sun, B.; Wang, X.; Liu, J. Changes in dimensional stability and mechanical properties of Eucalyptus pellita by melamine–urea–formaldehyde resin impregnation and heat treatment. Eur. J. Wood Wood Prod. 2013, 71, 557–562. [Google Scholar] [CrossRef]
- Behr, G.; Bollmus, S.; Gellerich, A.; Militz, H. Improvement of mechanical properties of thermally modified hardwood through melamine treatment. Wood Mater. Sci. Eng. 2017, 13, 262–270. [Google Scholar] [CrossRef]
- Mubarok, M.; Dumarcay, S.; Militz, H.; Candelier, K.; Thévenon, M.-F.; Gérardin, P. Non-biocide antifungal and anti-termite wood preservation treatments based on combinations of thermal modification with different chemical additives. Eur. J. Wood Wood Prod. 2019, 77, 1125–1136. [Google Scholar] [CrossRef]
- Keey, R.B. Colour development on drying. Maderas-Cienc. Tecnol. 2005, 7, 3–16. [Google Scholar] [CrossRef]
- Marcos, M.; González-Penã, M.M.; Hale, M.D. Colour in thermally modified wood of beech, Norway spruce and Scots pine. Part 1: Colour evolution and colour changes. Holzforschung 2009, 63, 385–393. [Google Scholar] [CrossRef]
- Hauptmann, M.; Gindl-Altmutter, W.; Hansmann, C.; Bacher, M.; Rosenau, T.; Liebner, F.; D’Amico, S.; Schwanninger, M. Wood modification with tricine. Holzforschung 2015, 69, 985–991. [Google Scholar] [CrossRef]
- Peeters, K.; Larnøy, E.; Kutnar, A.; Hill, C.A.S. An examination of the potential for the use of the Maillard reaction to modify wood. Int. Wood Prod. J. 2018, 9, 108–114. [Google Scholar] [CrossRef] [Green Version]
- Popescu, C.-M.; Kržišnik, D.; Hočevar, M.; Humar, M.; Thaler, N.; Popescu, M.-C.; Jones, D. Infrared spectroscopy and chemometric methods for the evaluation of the thermal/chemical treatment effectiveness of hardwoods. In Proceedings of the Ninth European Conference on Wood Modification, Arnhem, The Netherlands, 17–18 September 2018; pp. 355–362. [Google Scholar]
- Popescu, C.-M.; Jones, D.; Kržišnik, D.; Humar, M. Determination of the effectiveness of a combined thermal/chemical wood modification by the use of FT–IR spectroscopy and chemometric methods. J. Mol. Struct. 2020, 1200, 127133. [Google Scholar] [CrossRef]
- Good, N.E.; Winget, G.D.; Winter, W.; Connolly, T.N.; Izawa, S.; Singh, R.M.M. Hydrogen ion buffers for biological research. Biochemistry 1966, 5, 467–477. [Google Scholar] [CrossRef]
- Ferreira, C.M.M.; Pinto, I.S.S.; Soares, E.V.; Soares, H.V.M. (Un)suitability of the use of pH buffers in biological, biochemical and environmental studies and their interaction with metal ions—A review. RSC Adv. 2015, 5, 30989–31003. [Google Scholar] [CrossRef] [Green Version]
- Soni, M.L.; Kapoor, R.C. Some thermodynamic parameters for hydroxyl amino acids: Bicine and tricine. Int. J. Quantum Chem. 1981, 20, 385–391. [Google Scholar] [CrossRef]
- Armenante, P.M.; Kafkewitz, D.; Jou, C.-J.; Lewandowski, G. Effect of pH on the anaerobic dechlorination of chlorophenols in a defined medium. Appl. Microbiol. Biotechnol. 1993, 39, 772–777. [Google Scholar] [CrossRef]
- Hicks, M.; Gebicki, J.M. Rate constants for reaction of hydroxyl radicals with Tris, Tricine and Hepes buffers. FEBS J. 1986, 199, 92–94. [Google Scholar] [CrossRef] [Green Version]
- Shiraishi, H.; Kataoka, M.; Morita, Y.; Umemoto, J. Interactions of hydroxyl radicals with tris (hydroxymethyl) aminomethane and Good’s buffers containing hydroxymethyl or hydroxyethyl residues produce formaldehyde. Free Rad. Res. Commun. 1993, 19, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Brune, A.; Friedrich, M. Microecology of the termite gut: Structure and function on a microscale. Curr. Opin. Microbiol. 2000, 3, 263–269. [Google Scholar] [CrossRef]
- Brune, A.; Emerson, D.; Breznak, J.A. The termite gut microflora as an oxygen sink: Microelectrode determination of oxygen and pH gradients in guts of lower and higher termites. Appl. Environ. Microbiol. 1995, 61, 2681–2687. [Google Scholar] [CrossRef] [Green Version]
- Brugerolle, G. Flagellar and cytoskeletal systems in amitochondrial flagellates: Archamoeba, Metamonada and Parabasala. Protoplasma 1991, 164, 70–90. [Google Scholar] [CrossRef]
- Čepička, I.; Hampl, V.; Kulda, J. Critical taxonomic revision of parabasalids with description of one new genus and three new species. Protist 2010, 161, 400–433. [Google Scholar] [CrossRef]
- Adl, S.M.; Simpson, A.G.B.; Lane, C.E.; Lukes, J.; Bass, D.; Bowser, S.S.; Brown, M.W.; Burki, F.; Dunthorn, M.; Hampl, V.; et al. The revised classification of Eukaryotes. J. Eukaryot. Microbiol. 2012, 59, 429–493. [Google Scholar] [CrossRef] [Green Version]
- Scharf, M.E. Termites as targets and models for biotechnology. Annu. Rev. Entomol. 2015, 60, 77–102. [Google Scholar] [CrossRef]
- Tanaka, H.; Aoyagi, H.; Shina, S.; Dodo, Y.; Yoshimura, T.; Nakamura, R.; Uchiyama, H. Influence of the diet components on the symbiotic microorganisms community in hindgut of Coptotermes formosanus Shiraki. Appl. Microbial. Cell Physiol. 2006, 71, 907–917. [Google Scholar] [CrossRef]
- Hu, X.P.; Song, D.; Gao, X. Biological changes in the Eastern subterranean termite, Reticulitermes flavipes (Isoptera, Rhinotermitidae) and its protozoa profile following starvation. Insectes Soc. 2011, 58, 39–45. [Google Scholar] [CrossRef]
- Raychoudhury, R.; Sen, R.; Cal, Y.; Sun, Y.; Lietze, V.U.; Boucias, D.G.; Scharf, M.E. Comparative metatranscriptomic signatures of wood and paper feeding in the gut of termite Reticulitermes flavipes (Isoptera: Rhinotermitidae). Insect Mol. Biol. 2013, 22, 155–171. [Google Scholar] [CrossRef]
- Arquette, T.J.; Champagne, D.E.; Brown, M.R.; Forschler, B.T. Evaluation of novel and traditional measures for vigor of laboratory-cultured termites, Reticulitermes flavipes (Kollar). J. Insect Physiol. 2006, 56, 51–66. [Google Scholar] [CrossRef]
- Rep, G.; Pohleven, F.; Košmerl, S. Development of industrial kiln for thermal wood modification by a procedure with an initial vacuum and commercialisation of modified Silvapro wood. In Proceedings of the Sixth European Conference on Wood Modification, Ljubljana, Slovenia, 17–18 September 2012; pp. 11–17. [Google Scholar]
- EN 117 2012. Wood Preservatives. Determination of Toxic Values against Reticulitermes Species (European Termites) (Laboratory Method); European Committee of Standardization: Brussels, Belgium.
- Esteves, B.; Ribeiro, F.; Cruz-Lopes, L.; Ferreira, J.; Domingos, I.; Duarte, M.; Duarte, S.; Nunes, L. Combined treatment by densification and heat treatment of maritime pine wood. Wood Res. 2017, 62, 373–388. [Google Scholar]
- Trager, W. The cultivation of a cellulose-digesting flagellate, Trichomonas termopsidis, and of certain other termite protozoa. Biol. Bull. 1934, 66, 182–190. [Google Scholar] [CrossRef]
- Duarte, S.; Nobre, T.; Borges, P.A.V.; Nunes, L. Symbiotic flagellate protists as cryptic drivers of adaptation and invasiveness of the subterranean termite Reticulitermes grassei Clément. Ecol. Evol. 2018, 8, 5242–5253. [Google Scholar] [CrossRef]
- Leidy, J. On intestinal parasites of Termes flavipes. Proc. Acad. Nat. Sci. Phila. 1877, 29, 146–149. [Google Scholar]
- Brugerolle, G.; Lee, J.J. Order Oxymonadida. Phylum Parabasalia. In The Illustrated Guide to the Protozoa; Lee, J.J., Leedale, G.F., Bradbury, P., Eds.; Society of Protozoologists: Lawrence, KS, USA, 2000; Volume II, pp. 1186–1250. [Google Scholar]
- Brugerolle, G.; Bordereau, C. Immunological and ultrastructural characterization of spirotrichonymphid flagellates from Reticulitermes grassei and R. flavipes (syn. R. santonensis), with special reference to Spirotrichonympha, Spironympha and Microjoenia. Org. Divers. Evol. 2006, 6, 109–123. [Google Scholar] [CrossRef] [Green Version]
- Lewis, J.L.; Forschler, B.T. A nondichotomous key to protist species identification of Reticulitermes (Isoptera: Rhinotermitidae). Ann. Entomol. Soc. Am. 2006, 99, 1028–1033. [Google Scholar] [CrossRef]
- Cornelius, M.L.; Osbrink, W.L.A. Bioassay design and length of time in the laboratory affect intercolonial interactions of the Formosan subterranean termite (Isoptera, Rhinotermitidae). Insectes Soc. 2009, 56, 203–211. [Google Scholar] [CrossRef] [Green Version]
- Windeisen, E.; Strobel, C.; Wegener, G. Chemical changes during the production of thermo-treated beech wood. Wood Sci. Technol. 2007, 41, 523–536. [Google Scholar] [CrossRef]
- Tai, V.; James, E.R.; Nalepa, C.A.; Scheffrahn, R.H.; Perlman, S.J.; Keeling, P.J. The role of host phylogeny varies in shaping microbial diversity in the hindguts of lower termites. Appl. Environ. Microbiol. 2015, 81, 1059–1070. [Google Scholar] [CrossRef] [Green Version]
- Duarte, S.; Duarte, M.; Borges, P.A.V.; Nunes, L. Dietary-driven variation effects on the symbiotic flagellate protist communities of the subterranean termite Reticulitermes grassei Clément. J. Appl. Entomol. 2017, 141, 300–307. [Google Scholar] [CrossRef]
- Bignell, D.E.; Anderson, J.M. Determination of pH and oxygen status in the guts of lower and higher termites. J. Insect Physiol. 1980, 26, 183–188. [Google Scholar] [CrossRef]


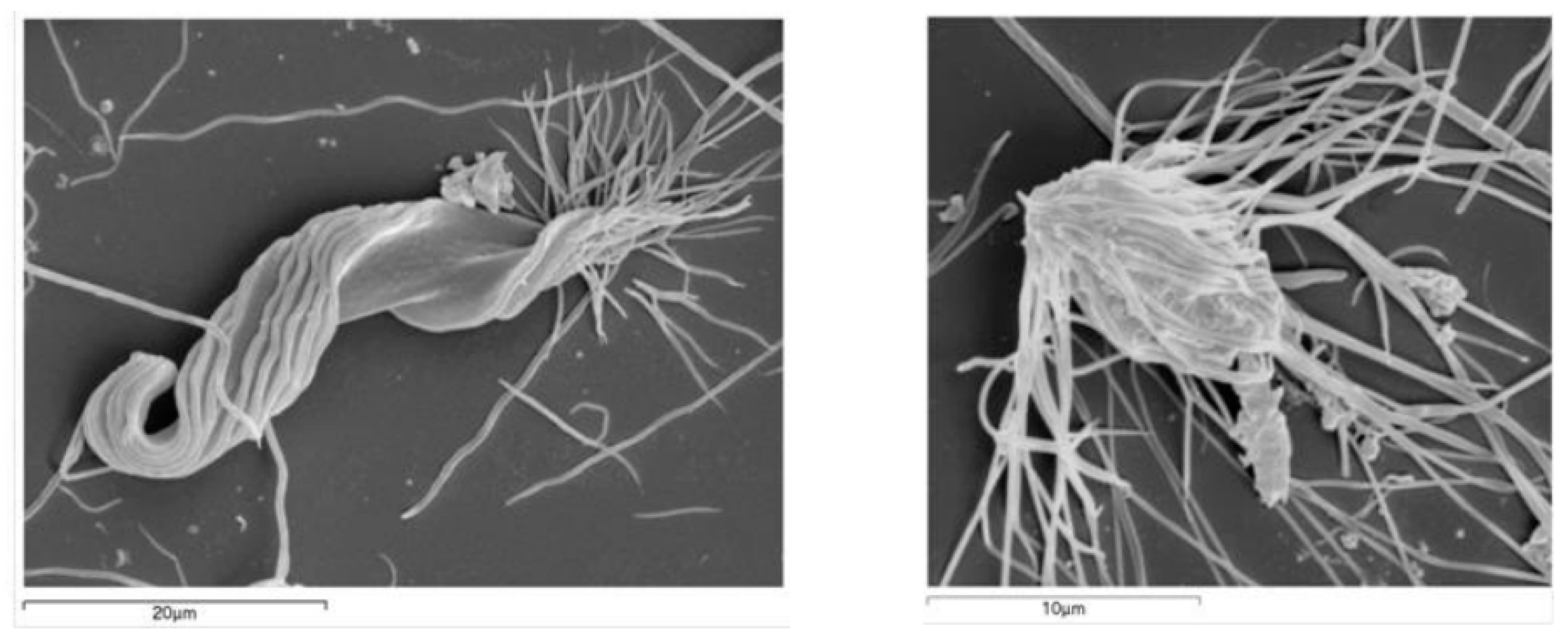{kind=link}
| Wood Species | Treatment | Survival (%) | Mass Loss (%) | ||
|---|---|---|---|---|---|
| Pine | Untreated | 84.4 ± 7.4 | - | 18.3 ± 2.9 | - |
| Spruce | Untreated | 88.4 ± 6.3 | a | 19.5 ± 0.8 | a |
| HT | 90.8 ± 4.5 | a | 19.0 ± 1.9 | a | |
| Bicine | 64.2 ± 38.9 | ab | 11.7 ± 1.0 | b | |
| Bicine HT | 21.1 ± 18.7 | b | 10.8 ± 1.5 | b | |
| Tricine | 11.3 ± 17.4 | b | 11.5 ± 1.1 | ab | |
| Tricine HT | 62.3 ± 31.2 | ab | 17.5 ± 0.8 | ab | |
| Beech | Untreated | 82.0 ± 9.7 | a | 15.9 ± 0.8 | a |
| HT | 87.2 ± 2.7 | a | 14.8 ± 0.8 | a | |
| Bicine | 38.0 ± 22.0 | a | 12.3 ± 0.3 | b | |
| Bicine HT | 57.6 ± 47.0 | a | 11.4 ± 1.3 | b | |
| Tricine | 71.4 ± 17.2 | a | 14.2 ± 0.3 | b | |
| Tricine HT | 79.9 ± 16.8 | a | 13.0 ± 0.0 | ab |
| Wood Species | Treatment | |||||
|---|---|---|---|---|---|---|
| Untreated | HT | Bicine | Bicine HT | Tricine | Tricine HT | |
| Spruce | 4 | 4 | 3 | 4 | 4 | 4 |
| 4 | 4 | 3 | 3 | 3 | 4 | |
| 4 | 4 | 3 | 1 | 2 | 4 | |
| Beech | 4 | 4 | 3 | 4 | 4 | 4 |
| 4 | 4 | 3 | 3 | 4 | 4 | |
| 4 | 4 | 3 | 2 | 3 | 4 | |
| Pine | 4 | - | - | - | - | - |
| 4 | - | - | - | - | - | |
| 4 | - | - | - | - | - | |
| Phylum | Class | Order | Family | Genus | Species | |
|---|---|---|---|---|---|---|
| n1 | Parabasalia | Trychonymphea | Trichonymphida | Trichonymphidae | Trichonympha | |
| n9 | Spirotrichonymphea | Spirotrichonymphida | Holomastigotoididae | Spirotrichonympha | S. flagellata | |
| n4 | Holomastigotes | H. elongatum | ||||
| n8 | Microjoenia | M. hexamitoides | ||||
| n6 | Hypotrichomonadea | Hypotrichomonadida | Hypotrichomonadidae | |||
| n2 | Preaxostyla | Oxymonadida | Pyrsonympha sp.1 | |||
| n7 | Pyrsonympha sp.2 | |||||
| n3 | Dinenympha | D. gracilis | ||||
| n5 | D. fimbriata |
| Treatment | n1 | n2 | n3 | n4 | n5 | n6 | n7 | n8 | n9 | |
|---|---|---|---|---|---|---|---|---|---|---|
| Pine | 0.86 ± 0.78 A | 12.06 ± 3.28 A | 2.62 ± 1.84 A | 0.24 ± 0.48 A | 0.30 ± 0.65 A | 0.06 ± 0.24 A | 0.22 ± 0.58 A | 4.56 ± 2.84 A | 1.42 ± 1.05 A | |
| Day 1 | Control | 0.64 ± 0.76 Aab | 8.24 ± 3.53 Ba | 2.16 ± 1.34 Aa | 0.08 ± 0.28 Aa | 0.24 ± 0.60 Aa | 0.20 ± 0.41 Aa | 0.20 ± 0.41 Aa | 1.88 ± 1.05 Ba | 1.28 ± 1.21 Aab |
| HT | 0.76 ± 0.66 Ab | 10.84 ± 4.04 Aa | 3.92 ± 1.53 Bb | 0.32 ± 0.48 Aa | 0.44 ± 0.58 Aa | 0.04 ± 0.20 Aa | 0.28 ± 0.46 Aa | 2.92 ± 1.68 Aa | 1.52 ± 1.19 Ab | |
| B | 0.44 ± 0.58 Aab | 9.40 ± 3.86 Ba | 2.36 ± 1.47 Aac | 0.28 ± 0.46 Aa | 0.16 ± 0.37 Aa | 0.12 ± 0.33 Aa | 0.08 ± 0.28 Aa | 3.40 ± 2.45 Aa | 0.88 ± 0.97 Aab | |
| BHT | 0.64 ± 0.81 Aab | 9.68 ± 3.58 Ba | 2.92 ± 1.53 Aab | 0.28 ± 0.46 Aa | 0.04 ± 0.20 Aa | 0.00 ± 0.00 Aa | 0.08 ± 0.28 Aa | 3.12 ± 1.90 Aa | 0.68 ± 0.69 Ba | |
| T | 0.20 ± 0.41 Ba | 9.76 ± 4.56 Aa | 1.72 ± 1.14 Aa | 0.12 ± 0.33 Aa | 0.20 ± 0.41 Aa | 0.08 ± 0.28 Aa | 0.20 ± 0.41 Aa | 2.40 ± 1.29 Ba | 0.64 ± 0.76 Ba | |
| THT | 0.36 ± 0.57 Bab | 10.32 ± 3.16 Aa | 3.56 ± 1.87 Abc | 0.24 ± 0.44 Aa | 0.20 ± 0.41 Aa | 0.12 ± 0.33 Aa | 0.08 ± 0.28 Aa | 2.80 ± 1.80 Ba | 1.24 ± 1.05 Aab | |
| Day 7 | Control | 0.60 ± 0.76 Aab | 8.00 ± 3.35 Ba | 1.85 ± 1.19 Aa | 0.15 ± 0.37 Aa | 0.10 ± 0.28 Aa | 0.10 ± 0.33 Aa | 0.25 ± 0.41 Aa | 2.90 ± 2.27 Ba | 1.30 ± 1.19 Aa |
| HT | 0.72 ± 0.68 Ab | 8.92 ± 2.43 Ba | 4.20 ± 2.18 Bb | 0.12 ± 0.33 Aa | 0.12 ± 0.33 Aa | 0.12 ± 0.33 Aa | 0.12 ± 0.33 Aa | 4.60 ± 3.44 Ba | 1.32 ± 1.07 Aa | |
| B | 0.28 ± 0.54 Bab | 7.68 ± 2.70 Ba | 1.24 ± 1.13 Bac | 0.20 ± 0.41 Aa | 0.00 ± 0.00 Ba | 0.12 ± 0.33 Aa | 0.04 ± 0.20 Aa | 2.64 ± 1.91 Ba | 0.56 ± 0.77 Bb | |
| BHT | 0.36 ± 0.57 Bab | 7.48 ± 3.92 Ba | 0.96 ± 0.89 Bc | 0.08 ± 0.28 Aa | 0.00 ± 0.00 Ba | 0.24 ± 0.44 Aa | 0.12 ± 0.33 Aa | 1.44 ± 1.56 Ba | 0.36 ± 0.57 Bab | |
| T | 0.20 ± 0.41 Ba | 7.32 ± 3.20 Ba | 2.32 ± 1.25 Aa | 0.08 ± 0.28 Aa | 0.04 ± 0.20 Aa | 0.20 ± 0.41 Aa | 0.04 ± 0.20 Aa | 2.88 ± 2.49 Aa | 0.84 ± 0.80 Bab | |
| THT | 0.44 ± 0.58 Aab | 8.64 ± 3.65 Ba | 2.16 ± 1.46 Aa | 0.08 ± 0.28 Aa | 0.00 ± 0.00 Ba | 0.04 ± 0.20 Ba | 0.08 ± 0.28 Aa | 3.12 ± 2.01 Aa | 0.72 ± 0.94 Bb | |
| Day 14 | Control | 0.92 ± 0.76 Aa | 9.88 ± 3.31 Bac | 4.20 ± 1.91 Ba | 0.32 ± 0.56 Aa | 0.08 ± 0.28 Aa | 0.24 ± 0.52 Aa | 0.32 ± 0.56 Aa | 3.40 ± 1.80 Aa | 1.84 ± 1.18 Aa |
| HT | 0.28 ± 0.46 Bb | 10.2 ± 2.36 Bac | 3.36 ± 1.58 Aa | 0.08 ± 0.28 Aa | 0.04 ± 0.20 Aa | 0.12 ± 0.33 Aa | 0.28 ± 0.54 Aa | 6.20 ± 3.44 Ab | 1.20 ± 1.12 Aab | |
| B | 0.24 ± 0.44 Bb | 6.32 ± 3.67 Bbc | 2.12 ± 1.62 Ab | 0.12 ± 0.33 Aa | 0.00 ± 0.00 Ba | 0.12 ± 0.33 Aa | 0.00 ± 0.00 Aa | 3.00 ± 2.45 Aa | 0.52 ± 0.59 Bb | |
| BHT | 0.08 ± 0.28 Bb | 7.84 ± 3.59 Bc | 2.08 ± 1.53 Ab | 0.00 ± 0.00 Aa | 0.00 ± 0.00 Ba | 0.08 ± 0.28 Aa | 0.12 ± 0.33 Aa | 2.84 ± 2.37 Ba | 0.76 ± 0.88 Bb | |
| T | 0.16 ± 0.37 Bb | 11.48 ± 3.72 Aa | 2.24 ± 1.13 Ab | 0.12 ± 0.33 Aa | 0.00 ± 0.00 Ba | 0.16 ± 0.47 Aa | 0.08 ± 0.28 Aa | 3.44 ± 2.81 Aa | 0.64 ± 0.70 Bb | |
| THT | 0.20 ± 0.41 Bb | 9.20 ± 3.77 Bac | 2.72 ± 1.70 Ab | 0.04 ± 0.20 Aa | 0.00 ± 0.00 Ba | 0.28 ± 0.54 Aa | 0.24 ± 0.52 Aa | 3.44 ± 1.53 Aa | 0.72 ± 1.02 Bb | |
| Day 21 | Control | 0.68 ± 0.75 Aa | 9.40 ± 4.70 Bac | 2.00 ± 0.96 Aab | 0.20 ± 0.41 Aa | 0.00 ± 0.00 Ba | 0.16 ± 0.37 Aa | 0.36 ± 0.49 Aa | 3.84 ± 2.29 Aa | 1.08 ± 1.00 Aa |
| HT | 0.48 ± 0.65 Aab | 11.20 ± 4.32 Aa | 2.84 ± 2.15 Aa | 0.24 ± 0.44 Aa | 0.00 ± 0.00 Ba | 0.16 ± 0.47 Aa | 0.20 ± 0.41 Aa | 3.44 ± 1.87 Aac | 1.92 ± 1.47 Aa | |
| B | 0.40 ± 0.58 Bab | 6.56 ± 3.18 Bb | 1.40 ± 1,41 Bbc | 0.04 ± 0.20 Aa | 0.00 ± 0.00 Ba | 0.04 ± 0.20 Aa | 0.04 ± 0.20 Ab | 2.80 ± 1.32 Bab | 0.16 ± 0.37 Bb | |
| BHT | 0.16 ± 0.37 Bb | 7.28 ± 3.29 Bc | 1.24 ± 1.05 Bc | 0.00 ± 0.00 Aa | 0.00 ± 0.00 Ba | 0.16 ± 0.37 Aa | 0.04 ± 0.20 Ab | 2.32 ± 2.17 Bb | 0.28 ± 0.54 Bb | |
| T | 0.32 ± 0.48 Bab | 10.36 ± 3.39 Ab | 1.88 ± 1.13 Aabc | 0.24 ± 0.52 Aa | 0.00 ± 0.00 Ba | 0.32 ± 0.48 Aa | 0.08 ± 0.28 Aa | 2.36 ± 1.93 Bbc | 0.40 ± 0.58 Bb | |
| THT | 0.20 ± 0.50 Bb | 5.84 ± 2.73 Bc | 1.44 ± 1.16 Babc | 0.08 ± 0.28 Aa | 0.00 ± 0.00 Ba | 0.20 ± 0.41 Aa | 0.16 ± 0.37 Aa | 2.52 ± 2.06 Bab | 0.44 ± 0.65 Bb | |
| Day 28 | Control | 0.92 ± 1.04 Aa | 6.84 ± 2.79 Ba | 3.24 ± 1.39 Aa | 0.28 ± 0.46 Aa | 0.08 ± 0.28 Aa | 0.24 ± 0.52 Aa | 0.28 ± 0.54 Aa | 2.48 ± 1.48 Ba | 0.76 ± 0.66 Bab |
| HT | 0.44 ± 0.58 Ba | 12.68 ± 5.43 Ab | 2.56 ± 1.61 Aa | 0.04 ± 0.20 Aa | 0.04 ± 0.20 Aa | 0.12 ± 0.33 Aa | 0.24 ± 0.44 Aa | 3.08 ± 2.20 Ba | 1.48 ± 1.48 Ab | |
| B | 0.08 ± 0.28 Bb | 5.00 ± 1.71 Bc | 1.20 ± 1.08 Bb | 0.04 ± 0.20 Aa | 0.00 ± 0.00 Ba | 0.00 ± 0.00 Aa | 0.04 ± 0.20 Aa | 0.48 ± 0.51 Bb | 0.08 ± 0.28 Bc | |
| BHT | 0.08 ± 0.28 Bb | 3.44 ± 2.18 Bd | 1.04 ± 0.89 Bb | 0.00 ± 0.00 Ab | 0.00 ± 0.00 Ba | 0.08 ± 0.28 Aa | 0.00 ± 0.00 Aa | 0.24 ± 0.52 Bb | 0.80 ± 1.22 Bab | |
| T | 0.36 ± 0.57 Bab | 6.32 ± 3.21 Bac | 0.88 ± 0.73 Bb | 0.12 ± 0.33 Aa | 0.00 ± 0.00 Ba | 0.04 ± 0.20 Aa | 0.08 ± 0.28 Aa | 1.20 ± 0.96 Bb | 0.80 ± 0.91 Bab | |
| THT | 0.36 ± 0.49 Ba | 8.32 ± 4.98 Ba | 1.44 ± 1.42 Bb | 0.00 ± 0.00 Ab | 0.00 ± 0.00 Ba | 0.16 ± 0.37 Aa | 0.12 ± 0.44 Aa | 2.00 ± 1.35 Ba | 0.52 ± 0.65 Ba |
| Treatment | n1 | n2 | n3 | n4 | n5 | n6 | n7 | n8 | n9 | |
|---|---|---|---|---|---|---|---|---|---|---|
| Pine | 0.86 ± 0.78 A | 12.06 ± 3.28 A | 2.62 ± 1.84 A | 0.24 ± 0.48 A | 0.30 ± 0.65 A | 0.06 ± 0.24 A | 0.22 ± 0.58 A | 4.56 ± 2.84 A | 1.42 ± 1.05 A | |
| Day 1 | Control | 0.52 ± 0.65 Aa | 9.32 ± 2.91 Ba | 4.08 ± 2.33 Ba | 0.24 ± 0.44 Aa | 0.12 ± 0.33 Aa | 0.12 ± 0.33 Aa | 0.16 ± 0.37 Aa | 2.44 ± 1.73 Ba | 1.36 ± 1.04 Aa |
| HT | 0.72 ± 0.68 Aa | 8.88 ± 2.91 Bab | 3.04 ± 1.43 Aa | 0.20 ± 0.50 Aa | 0.28 ± 0.46 Aa | 0.12 ± 0.33 Aa | 0.28 ± 0.46 Aa | 3.28 ± 2.21 Aa | 1.44 ± 1.26 Aa | |
| B | 0.64 ± 0.70 Aa | 7.20 ± 3.37 Bab | 3.12 ± 1.81 Aa | 0.20 ± 0.41 Aa | 0.16 ± 0.47 Aa | 0.12 ± 0.33 Aa | 0.16 ± 0.37 Aa | 2.52 ± 2.33 Ba | 0.64 ± 0.86 Bb | |
| BHT | 0.64 ± 0.86 Aa | 7.32 ± 4.41 Bb | 2.80 ± 1.66 Aab | 0.32 ± 0.56 Aa | 0.24 ± 0.44 Aa | 0.16 ± 0.37 Aa | 0.08 ± 0.28 Aa | 2.12 ± 1.42 Ba | 0.72 ± 0.79 Bab | |
| T | 0.40 ± 0.50 Aa | 7.48 ± 2.87 Bab | 3.48 ± 1.83 Aa | 0.08 ± 0.28 Aa | 0.08 ± 0.28 Aa | 0.04 ± 0.20 Aa | 0.16 ± 0.37 Aa | 1.96 ± 1,37 Ba | 0.88 ± 0.88 Aab | |
| THT | 0.48 ± 0.65 Aa | 4.72 ± 1.40 Bc | 1.76 ± 1.16 Ab | 0.04 ± 0.20 Aa | 0.04 ± 0.20 Aa | 0.04 ± 0.20 Aa | 0.08 ± 0.28 Aa | 1.96 ± 1,49 Ba | 1.28 ± 0.89 Aa | |
| Day 7 | Control | 0.72 ± 0.84 Aa | 10.32 ± 3.41 Ba | 2.80 ± 1.71 Aa | 0.16 ± 0.37 Aa | 0.00 ± 0.00 Aa | 0.12 ± 0.33 Aa | 0.32 ± 0.56 Aa | 3.72 ± 3.70 Aa | 1.28 ± 0.98 Aac |
| HT | 0.48 ± 0.51 Aa | 10.2 ± 3.50 Bac | 2.48 ± 1.56 Aab | 0.32 ± 0.63 Aa | 0.00 ± 0.00 Aa | 0.04 ± 0.20 Aa | 0.16 ± 0.47 Aa | 2.76 ± 1.20 Aa | 1.20 ± 0.87 Aac | |
| B | 0.64 ± 0.70 Aa | 8.28 ± 2.64 Bb | 1.60 ± 1.26 Abc | 0.16 ± 0.37 Aa | 0.00 ± 0.00 Aa | 0.12 ± 0.33 Aa | 0.20 ± 0.41 Aa | 3.60 ± 3.23 Aa | 0.84 ± 0.75 Bb | |
| BHT | 0.64 ± 0.76 Aa | 9.48 ± 4.45 Bbc | 2.52 ± 1.73 Aa | 0.12 ± 0.44 Aa | 0.08 ± 0.28 Aa | 0.20 ± 0.50 Aa | 0.20 ± 0.50 Aa | 3.16 ± 1.97 Aa | 1.16 ± 0.99 Bab | |
| T | 0.36 ± 0.57 Bab | 10.6 ± 5.02 Bab | 3.04 ± 1.81 Aa | 0.08 ± 0.28 Aa | 0.04 ± 0.20 Aa | 0.08 ± 0.28 Aa | 0.24 ± 0.44 Aa | 6.28 ± 2.70 Ba | 0.88 ± 0.73 Bbc | |
| THT | 0.16 ± 0.47 Bb | 6.96 ± 2.37 Bc | 1.52 ± 0.96 Bc | 0.08 ± 0.28 Aa | 0.00 ± 0.00 Ba | 0.16 ± 0.37 Aa | 0.12 ± 0.33 Aa | 4.60 ± 2.50 Ba | 0.36 ± 0.70 Bb | |
| Day 14 | Control | 0.72 ± 0.74 Aa | 12.40 ± 3.52 Aa | 3.44 ± 1.87 Aa | 0.24 ± 0.44 Aa | 0.00 ± 0.00 Ba | 0.28 ± 0.46 Aa | 0.20 ± 0.50 Aa | 3.84 ± 2.15 Aa | 1.88 ± 1.30 Aa |
| HT | 0.60 ± 0.71 Aab | 8.64 ± 3.38 Bbc | 2.84 ± 1.68 Aa | 0.16 ± 0.37 Aa | 0.04 ± 0.20 Aa | 0.08 ± 0.28 Aa | 0.16 ± 0.37 Aa | 4.56 ± 2.57 Aa | 1.56 ± 1.33 Aa | |
| B | 0.32 ± 0.48 Bab | 8.40 ± 3.55 Bbc | 2.36 ± 1.55 Aa | 0.12 ± 0.33 Aa | 0.00 ± 0.00 Ba | 0.20 ± 0.50 Aa | 0.12 ± 0.33 Aa | 3.56 ± 2.72 Aa | 0.40 ± 0.58 Bb | |
| BHT | 0.28 ± 0.54 Bab | 12.4 ± 3.99 Aa | 3.64 ± 2.27 Aa | 0.20 ± 0.41 Aa | 0.00 ± 0.00 Ba | 0.08 ± 0.28 Aa | 0.24 ± 0.44 Aa | 3.96 ± 2.84 Aa | 0.72 ± 0.68 Bb | |
| T | 0.52 ± 0.71 Aab | 9.16 ± 2.84 Bb | 2.32 ± 1.49 Aa | 0.12 ± 0.33 Aa | 0.00 ± 0.00 Ba | 0.24 ± 0.52 Aa | 0.24 ± 0.52 Aa | 3.96 ± 2.79 Aa | 0.64 ± 0.76 Bb | |
| THT | 0.20 ± 0.41 Bb | 7.24 ± 2.59 Bc | 2.92 ± 2.06 Aa | 0.08 ± 0.28 Aa | 0.00 ± 0.00 Ba | 0.12 ± 0.33 Aa | 0.28 ± 0.61 Aa | 4.32 ± 2.85 Aa | 0.84 ± 0.90 Ba | |
| Day 21 | Control | 0.64 ± 0.70 Aa | 9.80 ± 3.32 Ba | 3.48 ± 1.83 Ba | 0.24 ± 0.52 Aa | 0.00 ± 0.00 Ba | 0.20 ± 0.41 Aa | 0.20 ± 0.41 Aa | 3.00 ± 1.76 Bab | 1.40 ± 1.00 Aa |
| HT | 0.48 ± 0.59 Aa | 8.84 ± 3.30 Bab | 3.68 ± 2.17 Bab | 0.12 ± 0.33 Aa | 0.04 ± 0.20 Aa | 0.12 ± 0.33 Aa | 0.24 ± 0.44 Aa | 3.56 ± 2.26 Bb | 1.32 ± 1.38 Aa | |
| B | 0.20 ± 0.41 Ba | 10.28 ± 4.77 Ba | 1.48 ± 0.87 Abc | 0.04 ± 0.20 Aa | 0.00 ± 0.00 Ba | 0.24 ± 0.52 Aa | 0.12 ± 0.33 Aa | 2.08 ± 1.35 Ba | 0.48 ± 0.71 Bb | |
| BHT | 0.36 ± 0.64 Aa | 8.60 ± 2.92 Bb | 1.76 ± 1.16 Abc | 0.08 ± 0.28 Aa | 0.00 ± 0.00 Ba | 0.08 ± 0.28 Aa | 0.04 ± 0.20 Aa | 1.84 ± 1.72 Ba | 0.84 ± 1.52 Bb | |
| T | 0.48 ± 0.65 Aa | 8.56 ± 3.81 Bb | 2.16 ± 1.97 Ab | 0.12 ± 0.33 Aa | 0.00 ± 0.00 Ba | 0.08 ± 0.28 Aa | 0.12 ± 0.33 Aa | 2.04 ± 2.41 Ba | 1.36 ± 1.82 Aab | |
| THT | 0.16 ± 0.37 Ba | 7.80 ± 2.00 Bc | 1.72 ± 1.49 Bc | 0.16 ± 0.37 Aa | 0.00 ± 0.00 Ba | 0.12 ± 0.33 Aa | 0.04 ± 0.20 Aa | 3.00 ± 2.31 Bab | 0.80 ± 1.12 Aab | |
| Day 28 | Control | 0.76 ± 0.72 Aa | 8.72 ± 3.47 Bab | 2.52 ± 1.42 Aa | 0.40 ± 0.87 Aa | 0.00 ± 0.00 Ba | 0.36 ± 1.04 Aa | 0.16 ± 0.37 Aa | 2.12 ± 1.13 Ba | 0.92 ± 0.91 Ba |
| HT | 0.28 ± 0.46 Ba | 9.28 ± 2.69 Bb | 2.84 ± 1.86 Aa | 0.04 ± 0.20 Aab | 0.00 ± 0.00 Ba | 0.16 ± 0.37 Aa | 0.08 ± 0.28 Aa | 1.04 ± 0.93 Bbd | 1.16 ± 0.94 Aa | |
| B | 0.12 ± 0.33 Ba | 7.32 ± 4.49 Bac | 1.32 ± 1.25 Bb | 0.00 ± 0.00 Ab | 0.00 ± 0.00 Ba | 0.20 ± 0.50 Aa | 0.24 ± 0.44 Aa | 1.12 ± 1.09 Bbd | 0.16 ± 0.47 Bb | |
| BHT | 0.24 ± 0.44 Ba | 5.40 ± 2.52 Bc | 1.76 ± 1.64 Ab | 0.20 ± 0.41 Aab | 0.00 ± 0.00 Ba | 0.08 ± 0.28 Aa | 0.08 ± 0.28 Aa | 0.88 ± 0.97 Bbc | 0.08 ± 0.40 Bb | |
| T | 0.52 ± 0.51 Aa | 5.84 ± 2.27 Bc | 1.48 ± 1.16 Bb | 0.00 ± 0.00 Ab | 0.00 ± 0.00 Ba | 0.04 ± 0.20 Aa | 0.12 ± 0.44 Aa | 0.40 ± 0.71 Bc | 0.16 ± 0.37 Bb | |
| THT | 0.08 ± 0.28 Ba | 6.80 ± 2.25 Bac | 1.40 ± 1.08 Bb | 0.04 ± 0.20 Aab | 0.00 ± 0.00 Ba | 0.12 ± 0.33 Aa | 0.08 ± 0.28 Aa | 1.68 ± 1.25 Bad | 0.08 ± 0.28 Bb |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duarte, S.; Nunes, L.; Kržišnik, D.; Humar, M.; Jones, D. Influence of Zwitterionic Buffer Effects with Thermal Modification Treatments of Wood on Symbiotic Protists in Reticulitermes grassei Clément. Insects 2021, 12, 139. https://doi.org/10.3390/insects12020139
Duarte S, Nunes L, Kržišnik D, Humar M, Jones D. Influence of Zwitterionic Buffer Effects with Thermal Modification Treatments of Wood on Symbiotic Protists in Reticulitermes grassei Clément. Insects. 2021; 12(2):139. https://doi.org/10.3390/insects12020139
Chicago/Turabian StyleDuarte, Sónia, Lina Nunes, Davor Kržišnik, Miha Humar, and Dennis Jones. 2021. "Influence of Zwitterionic Buffer Effects with Thermal Modification Treatments of Wood on Symbiotic Protists in Reticulitermes grassei Clément" Insects 12, no. 2: 139. https://doi.org/10.3390/insects12020139
APA StyleDuarte, S., Nunes, L., Kržišnik, D., Humar, M., & Jones, D. (2021). Influence of Zwitterionic Buffer Effects with Thermal Modification Treatments of Wood on Symbiotic Protists in Reticulitermes grassei Clément. Insects, 12(2), 139. https://doi.org/10.3390/insects12020139