
Models Applied to Grapevine Pests: A Review


Abstract
:Simple Summary
Abstract
1. Introduction
2. What Is a Model?
3. Predicted Phenomena in Entomology
3.1. Pest Population Growth and Dynamic
3.2. Pest Invasion Risk
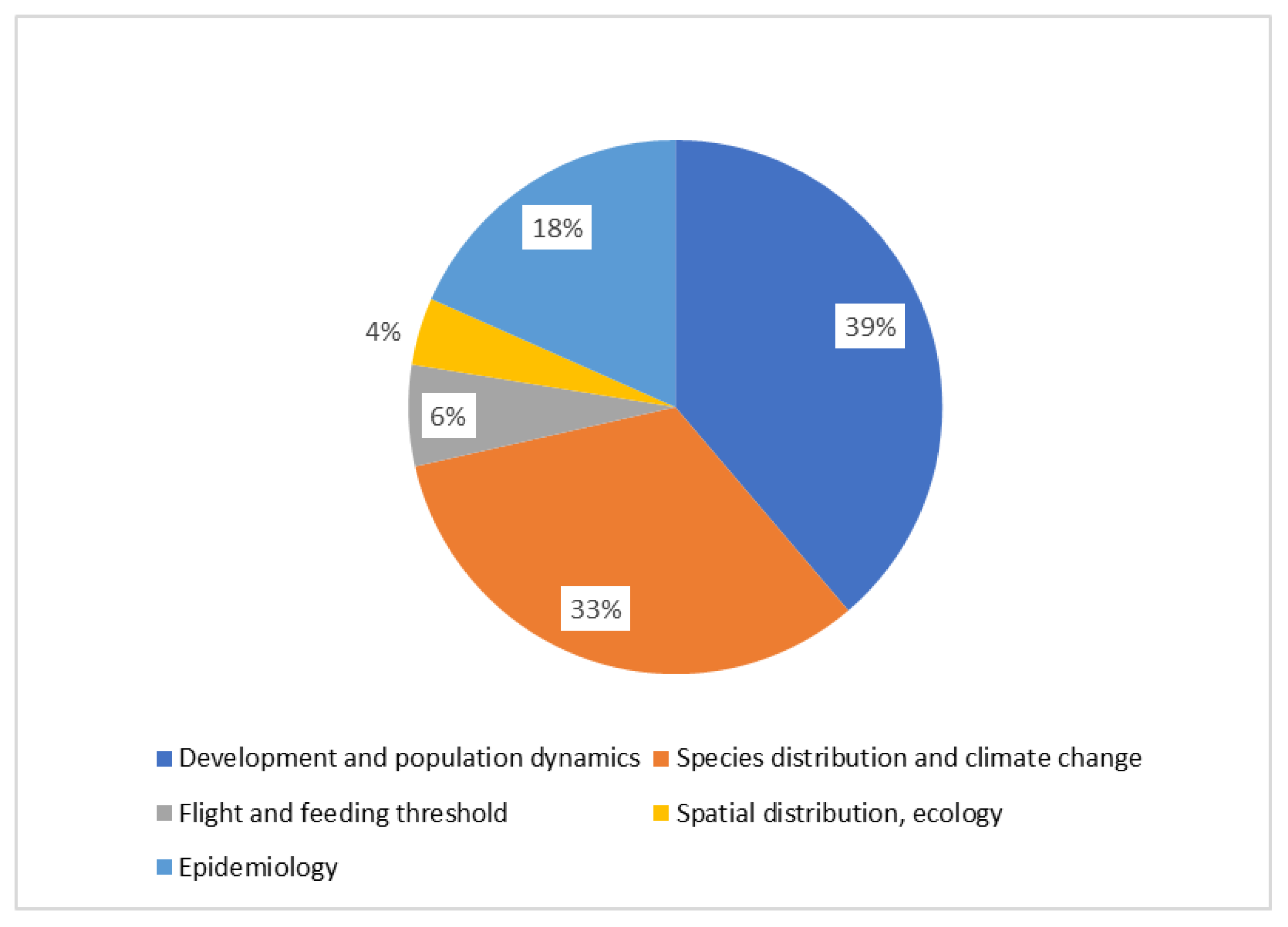
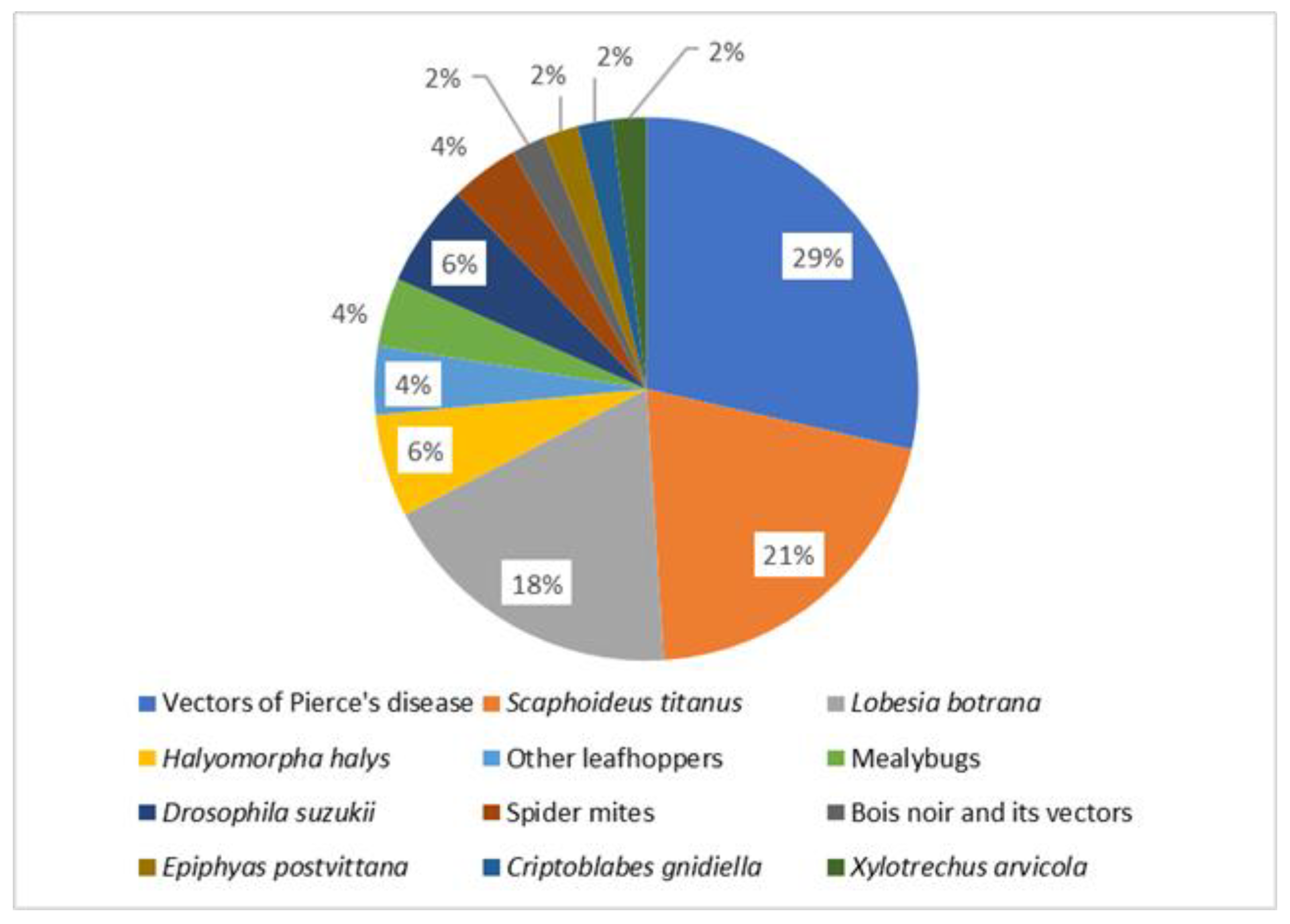
4. Modelling Grapevine Pests
4.1. Case Study 1: Grapevine Yellows and Their Vectors
4.2. Case Study 2: Grape Berry Moths
4.3. Case Study 3: Pierce’s Disease and Its Vectors
4.4. Other Species
5. Decision Support Systems: Present and Future
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Pertot, I.; Caffi, T.; Rossi, V.; Mugnai, L.; Hoffmann, C.; Grando, M.S.; Gary, C.; Lafond, D.; Duso, C.; Thiery, D.; et al. A critical review of plant protection tools for reducing pesticide use on grapevine and new perspectives for the implementation of IPM in viticulture. Crop Prot. 2017, 97, 70–84. [Google Scholar] [CrossRef]
- Rossi, V.; Salinari, F.; Poni, S.; Caffi, T.; Bettati, T. Addressing the implementation problem in agricultural decision support systems: The example of vite.net (R). Comput. Electron. Agric. 2014, 100, 88–99. [Google Scholar] [CrossRef]
- Rossi, V.; Caffi, T.; Salinari, F. Helping farmers face the increasing complexity of decision-making for crop protection. Phytopathol. Mediterr. 2012, 51, 457–479. [Google Scholar]
- Gilioli, G.; Pasquali, S.; Marchesini, E. A modelling framework for pest population dynamics and management: An application to the grape berry moth. Ecol. Model. 2016, 320, 348–357. [Google Scholar] [CrossRef]
- Venette, R.C.; Kriticos, D.J.; Magarey, R.D.; Koch, F.H.; Baker, R.H.A.; Worner, S.P.; Raboteaux, N.N.G.; McKenney, D.W.; Dobesberger, E.J.; Yemshanov, D.; et al. Pest risk maps for invasive alien species: A roadmap for improvement. Bioscience 2010, 60, 349–362. [Google Scholar] [CrossRef]
- Tonnang, H.E.Z.; Herve, B.D.B.; Biber-Freudenberger, L.; Salifu, D.; Subramanian, S.; Ngowi, V.B.; Guimapi, R.Y.A.; Anani, B.; Kakmeni, F.M.M.; Affognong, H.; et al. Advances in crop insect modelling methods-Towards a whole system approach. Ecol. Model. 2017, 354, 88–103. [Google Scholar] [CrossRef] [Green Version]
- Abramowitz, M.; Stegun, I.A.; Romer, R.H. Handbook of mathematical functions with formulas, graphs, and mathematical tables. Am. J. Phys. 1988, 56, 958. [Google Scholar] [CrossRef] [Green Version]
- Ningthoujam, S.; Talukdar, A.D.; Sarker, S.D.; Nahar, L.; Choudhury, M. Prediction of medicinal properties using mathematical models and computation, and selection of plant materials. Comput. Phytochem. 2018, 43–73. [Google Scholar] [CrossRef]
- Peck, S.L. A tutorial for understanding ecological modeling papers for the nonmodeler. Am. Entomol. 2000, 46, 40–49. [Google Scholar] [CrossRef] [Green Version]
- Briére, J.F.; Pracros, P.; Le Roux, A.Y.; Pierre, J.S. A novel rate model of temperature-dependent development for arthropods. Environ. Entomol. 1999, 28, 22–29. [Google Scholar] [CrossRef]
- Milonas, P.G.; Savopoulou-Soultani, M.; Stavridis, D.G. Day-degree models for predicting the generation time and flight activity of local populations of Lobesia botrana (Den. & Schiff.) (Lep., Tortricidae) in Greece. J. Appl. Entomol. 2001, 125, 515–518. [Google Scholar] [CrossRef]
- Gallardo, A.; Ocete, R.; Lopez, M.A.; Maistrello, L.; Ortega, F.; Semedo, A.; Soria, F.J. Forecasting the flight activity of Lobesia botrana (Denis & Schiffermuller) (Lepidoptera, Tortricidae) in Southwestern Spain. J. Appl. Entomol. 2009, 133, 626–632. [Google Scholar] [CrossRef]
- Logan, J.A.; Wollkind, D.J.; Hoyt, S.C.; Tanigoshi, L.K. An analytic model for description of temperature dependent rate phenomena in arthropods. Environ. Entomol. 1976, 5, 1133–1140. [Google Scholar] [CrossRef]
- Falzoi, S.; Lessio, F.; Spanna, F.; Alma, A. Influence of temperature on the embryonic and post-embryonic development of Scaphoideus titanus (Hemiptera: Cicadellidae), vector of grapevine Flavescence doree. Int. J. Pest Manag. 2014, 60, 246–257. [Google Scholar] [CrossRef]
- Briére, J.F.; Pracros, P. Comparison of temperature-dependent growth models with the development of Lobesia botrana (Lepidoptera: Tortricidae). Environ. Entomol. 1998, 27, 94–101. [Google Scholar] [CrossRef]
- Manetsch, T.J. Time-varying distributed delays and their use in aggregative models of large systems. IEEE Trans. Syst. Man Cybern. 1976, 8, 547–553. [Google Scholar] [CrossRef]
- Gilbert, E.; Powell, J.A.; Logan, J.A.; Bentz, B.J. Comparison of three models predicting developmental milestones given environmental and individual variation. Bull. Math. Biol. 2004, 66, 1821–1850. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.Q.; Chen, L.S.; Agarwal, R. Recent progress on stage-structured population dynamics. Math. Comput. Model. 2002, 36, 1319–1360. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, X.N.; Wei, Y.J. Dynamics of a stage-structured single population model with state-dependent delay. Adv. Differ. Equ. 2018, 2018, 364. [Google Scholar] [CrossRef]
- Maggi, F.; Marzachi, C.; Bosco, D. A stage-structured model of Scaphoideus titanus in vineyards. Environ. Entomol. 2013, 42, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Alma, A.; Lessio, F.; Nickel, H. Insects as phytoplasma vectors: Ecological and epidemiological aspects. In Phytoplasmas: Plant Pathogenic Bacteria-II. Transmission and Management of Phytoplasma-Associated Diseases; Bertaccini, A., Weintraub, P.G., Rao, G.P., Mori, N., Eds.; Springer: Singapore, 2019; pp. 1–25. [Google Scholar]
- Rigamonti, I.E.; Jermini, M.; Fuog, D.; Baumgaertner, J. Towards an improved understanding of the dynamics of vineyard-infesting Scaphoideus titanus leafhopper populations for better timing of management activities. Pest Manag. Sci. 2011, 67, 1222–1229. [Google Scholar] [CrossRef]
- Liebhold, A.M.; Rossi, R.E.; Kemp, W.P. Geostatistics and geographic information systems in applied insect ecology. Annu. Rev. Entomol. 1993, 38, 303–327. [Google Scholar] [CrossRef]
- Reineke, A.; Thiery, D. Grapevine insect pests and their natural enemies in the age of global warming. J. Pest Sci. 2016, 89, 313–328. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, A.P.; Ponti, L.; Hoddle, M.; Almeida, R.P.P.; Irvin, N.A. Geographic distribution and relative abundance of the invasive glassy-winged sharpshooter: Effects of temperature and egg parasitoids. Environ. Entomol. 2011, 40, 755–769. [Google Scholar] [CrossRef] [Green Version]
- Anderson, R.P.; Lew, D.; Peterson, A.T. Evaluating predictive models of species’ distributions: Criteria for selecting optimal models. Ecol. Model. 2003, 162, 211–232. [Google Scholar] [CrossRef]
- Warren, D.L.; Seifert, S.N. Ecological niche modeling in Maxent: The importance of model complexity and the performance of model selection criteria. Ecol. Appl. 2011, 21, 335–342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutherst, R.W.; Maywald, G.F.; Kriticos, D. CLIMEX Version 3. User’s Guide; Hearne Scientific Software Pty Ltd.: Melbourne, Australia, 2007; p. 131. [Google Scholar]
- Sutherst, R.W.; Maywald, G.F. CLIMEX—A biogeographical approach to entomology. In Pest Management: Future Challenges, Proceedings of the 6th Australasian Applied Entomological Research Conference, Brisbane, Australia, 29 September–2 October 1998; Zalucki, M.P., Drew, R., White, G.G., Eds.; University of Qld Press: Brisbane, Australia, 1998; Volumes 1–2, pp. 344–345. [Google Scholar]
- Baek, S.; Hwang, A.; Kim, H.; Lee, H.; Lee, J.H. Temperature-dependent development and oviposition models of Halyomorpha halys (Hemiptera: Pentatomidae). J. Asia Pac. Entomol. 2017, 20, 367–375. [Google Scholar] [CrossRef]
- Fraser, D.; Kumar, S.; Aguilar, G. Mapping the potential global range of the brown marmorated stink bug, Halyomorpha halys, with particular reference to New Zealand. Climate 2017, 5, 75. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.P.; Bu, W.J.; Gao, Y.B.; Liu, G.Q. Potential geographic distribution of brown marmorated stink bug invasion (Halyomorpha halys). PLoS ONE 2012, 7, e31246. [Google Scholar] [CrossRef]
- Yoon, J.M.; Hrynkiv, V.; Morano, L.; Nguyen, A.T.; Wilder, S.; Mitchell, F. Mathematical modeling of glassy-winged sharpshooter population. Math. Biosci. Eng. 2014, 11, 667–677. [Google Scholar] [CrossRef]
- Bostanian, N.J.; Bourgeois, G.; Vincent, C.; Plouffe, D.; Trudeau, M.; Lasnier, J. Modeling leafhopper nymphs in temperate vineyards for optimal sampling. Environ. Entomol. 2006, 35, 1477–1482. [Google Scholar] [CrossRef]
- Banks, H.T.; Banks, J.E.; Cody, N.G.; Hoddle, M.S.; Meade, A.E. Population model for the decline of Homalodisca vitripennis (Hemiptera: Cicadellidae) over a ten-year period. J. Biol. Dyn. 2019, 13, 422–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilkington, L.J.; Lewis, M.; Jeske, D.; Hoddle, M.S. Calculation and thematic mapping of demographic parameters for Homalodisca vitripennis (Hemiptera: Cicadellidae) in California. Ann. Entomol. Soc. Am. 2014, 107, 424–434. [Google Scholar] [CrossRef] [Green Version]
- Castle, S.J.; Naranjo, S.E.; Bi, J.L.; Byrne, F.J.; Toscano, N.C. Phenology and demography of Homalodisca coagulata (Hemiptera: Cicadellidae) in Southern California citrus and implications for management. Bull. Entomol. Res. 2005, 95, 621–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godefroid, M.; Cruaud, A.; Streito, J.C.; Rasplus, J.Y.; Rossi, J.P. Xylella fastidiosa: Climate suitability of European continent. Sci. Rep. 2019, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, R.; Mizell, R.F.; Andersen, P.C.; Brodbeck, B.V. Overwintering biology, food supplementation and parasitism of eggs of Homalodisca coagulata (Say) (Homoptera: Cicadellidae) by Gonatocerus ashmeadi Girault and Gonatocerus morrilli (Howard) (Hymenoptera: Mymaridae). J. Entomol. Sci. 2004, 39, 214–222. [Google Scholar] [CrossRef]
- Son, Y.; Groves, R.L.; Daane, K.M.; Morgan, D.J.W.; Krugner, R.; Johnson, M.W. Estimation of feeding threshold for Homalodisca vitripennis (Hemiptera: Cicadellidae) and its application to prediction of overwintering mortality. Environ. Entomol. 2010, 39, 1264–1275. [Google Scholar] [CrossRef]
- Mizell, R.F.; Tipping, C.; Andersen, P.C.; Brodbeck, B.V.; Hunter, W.B.; Northfield, T. Behavioral model for Homalodisca vitripennis (Hemiptera: Cicadellidae): Optimization of host plant utilization and management implications. Environ. Entomol. 2008, 37, 1049–1062. [Google Scholar] [CrossRef] [Green Version]
- Kyrkou, I.; Pusa, T.; Ellegaard-Jensen, L.; Sagot, M.F.; Hansen, L.H. Pierce’s disease of grapevines: A review of control strategies and an outline of an epidemiological model. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Daugherty, M.P.; Almeida, R.P.P. Understanding how an invasive vector drives pierce’s disease epidemics: Seasonality and vine-to-vine spread. Phytopathology 2019, 109, 277–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickman, A.G.; Dickman, R. Computational model of a vector-mediated epidemic. Am. J. Phys. 2015, 83, 468–474. [Google Scholar] [CrossRef] [PubMed]
- Lieth, J.H.; Meyer, M.M.; Yeo, K.H.; Kirkpatrick, B.C. Modeling cold curing of pierce’s disease in Vitis vinifera ‘Pinot Noir’ and ‘Cabernet Sauvignon’ grapevines in California. Phytopathology 2011, 101, 1492–1500. [Google Scholar] [CrossRef] [Green Version]
- Cerutti, F.; Baumgartner, J.; Delucchi, V. The dynamics of grape leafhopper Empoasca vitis Gothe populations in Switzerland and the implications for habitat management. Biocontrol Sci. Technol. 1991, 1, 177–194. [Google Scholar] [CrossRef]
- Kim, M.J.; Baek, S.; Lee, J.H. Egg hatching and first instar falling models of Metcalfa pruinosa (Hemiptera: Flatidae). Insects 2020, 11, 345. [Google Scholar] [CrossRef]
- Rotter, A.; Nikolic, P.; Turnsek, N.; Kogovsek, P.; Blejec, A.; Gruden, K.; Dermastia, M. Statistical modeling of long-term grapevine response to ‘Candidatus Phytoplasma solani’ infection in the field. Eur. J. Plant Pathol. 2018, 150, 653–668. [Google Scholar] [CrossRef] [Green Version]
- Ge, Q.; Wen, F. Predicting the potential geographical distribution of flavescence doree and its vector Scaphoideus titanus Ball in China using DIVA-GIS. Acta Phytophylacica Sin. 2006, 33, 51–58. [Google Scholar]
- Rigamonti, I.E.; Mariani, L.; Cola, G.; Jermini, M.; Baumgartner, J. Abrupt and gradual temperature changes influence on the climatic suitability of Northwestern Alpine grapevine-growing regions for the invasive grape leafhopper Scaphoideus titanus Ball (Hemiptera, Cicadellidae). Acta Oecologica Int. J. Ecol. 2018, 91, 22–29. [Google Scholar] [CrossRef]
- Quiroga, N.; Ivulic, D.; Saaverda, M.; Sandoval-Rodriguez, A.; Infante, R.; Morales, L.; Fiore, N. Risk analysis of the establishment of Scaphoideus titanus, vector of “flavescence dorée” phytoplasma in grapevine, under current and estimated climate change in Chle. Phytopathogenic Mollicutes 2017, 7, 39–44. [Google Scholar] [CrossRef]
- Steffek, R.; Reisenzein, H.; Strauss, G.; Leichtfried, T.; Hofrichter, J.; Kopacka, I.; Schwarz, M.; Pusterhofer, J.; Biedermann, R.; Renner, W.; et al. VitisCLIM, a project modelling epidemiology and economic impact of grapevine ‘flavescence doree’ phytoplasma in Austrian viticulture under a climate change scenario. Bull. Insectol. 2011, 64, S191–S192. [Google Scholar]
- Lessio, F.; Mondino, E.B.; Alma, A. Spatial patterns of Scaphoideus titanus (Hemiptera: Cicadellidae): A geostatistical and neural network approach. Int. J. Pest Manag. 2011, 57, 205–216. [Google Scholar] [CrossRef]
- Lessio, F.; Portaluri, A.; Paparella, F.; Alma, A. A mathematical model of flavescence doree epidemiology. Ecol. Model. 2015, 312, 41–53. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, A.P.; Daane, K.M.; Ponti, L.; Walton, V.M.; Ellis, C.K. Prospective evaluation of the biological control of vine mealybug: Refuge effects and climate. J. Appl. Ecol. 2008, 45, 524–536. [Google Scholar] [CrossRef]
- Atallah, S.S.; Gomez, M.I.; Conrad, J.M.; Nyrop, J.P. A plant-level, spatial, bioeconomic model of plant disease diffusion and control: Grapevine leafroll disease. Am. J. Agric. Econ. 2015, 97, 199–218. [Google Scholar] [CrossRef]
- Castex, V.; de Cortazar-Atauri, I.G.; Calanca, P.; Beniston, M.; Moreau, J. Assembling and testing a generic phenological model to predict Lobesia botrana voltinism for impact studies. Ecol. Model. 2020, 420. [Google Scholar] [CrossRef]
- Gutierrez, A.P.; Ponti, L.; Gilioli, G.; Baumgartner, J. Climate warming effects on grape and grapevine moth (Lobesia botrana) in the Palearctic region. Agric. For. Entomol. 2018, 20, 255–271. [Google Scholar] [CrossRef] [Green Version]
- Lv, W.; Li, Z.; Wu, X.; Ni, W.; Qv, W. Maximum Entropy Niche-Based Modeling (Maxent) of potential geographical distributions of Lobesia botrana (Lepidoptera: Tortricidae) in China. In Computer and Computing Technologies in Agriculture, Proceedings of the 5th International Conference in Computer and Computing Technologies in Agriculture, Beijing, China, 29–31 October 2011; Li, D., Chen, Y., Eds.; Springer: Berlin/Heidelberg, Germany, 2012. Part III. pp. 239–246. [Google Scholar]
- Perez-De la O, B.; Lopez-Martinez, V.; Jimenez-Garcia, D.; Campos-Figueroa, M. Model simulation of potential distribution of lobesia botrana (Denis & Schiffermuller) in grape, Vitis vinifera (L.) vineyard regions of Mexico. Southwest. Entomol. 2016, 41, 693–703. [Google Scholar]
- Gutierrez, A.P.; Ponti, L.; Cooper, M.L.; Gilioli, G.; Baumgartner, J.; Duso, C. Prospective analysis of the invasive potential of the European grapevine moth Lobesia botrana (Den. & Schiff.) in California. Agric. For. Entomol. 2012, 14, 225–238. [Google Scholar] [CrossRef]
- Amo-Salas, M.; Ortega-Lopez, V.; Harman, R.; Alonso-Gonzalez, A. A new model for predicting the flight activity of Lobesia botrana (Lepidoptera: Tortricidae). Crop Prot. 2011, 30, 1586–1593. [Google Scholar] [CrossRef]
- He, S.Q.; Worner, S.P.; Ikeda, T. Modeling the potential global distribution of light brown apple moth Epiphyas postvittana (Lepidoptera: Tortricidae) using CLIMEX. J. Asia Pac. Entomol. 2012, 15, 479–485. [Google Scholar] [CrossRef]
- Vidart, M.V.; Mujica, M.V.; Calvo, M.V.; Duarte, F.; Bentancourt, C.M.; Franco, J.; Scatoni, I.B. Relationship between male moths of Cryptoblabes gnidiella (Milliere) (Lepidoptera: Pyralidae) caught in sex pheromone traps and cumulative degree-days in vineyards in southern Uruguay. SpringerPlus 2013, 2. [Google Scholar] [CrossRef] [Green Version]
- Soria, F.J.; Lopez, M.A.; Perez, M.A.; Maistrello, L.; Armendariz, I.; Ocete, R. Predictive model for the emergence of Xylotrechus arvicola (Coleoptera: Cerambycidae) in La Rioja vineyards (Spain). Vitis 2013, 52, 91–96. [Google Scholar]
- Leach, H.; van Timmeren, S.; Wetzel, W.; Isaacs, R. Predicting within- and between-year variation in activity of the invasive spotted wing Drosophila (Diptera: Drosophilidae) in a temperate region. Environ. Entomol. 2019, 48, 1223–1233. [Google Scholar] [CrossRef]
- Rossini, L.; Contarini, M.; Giarruzzo, F.; Assennato, M.; Speranza, S. Modelling Drosophila suzukii adult male populations: A physiologically based approach with validation. Insects 2020, 11, 751. [Google Scholar] [CrossRef]
- de la Vega, G.J.; Corley, J.C. Drosophila suzukii (Diptera: Drosophilidae) distribution modelling improves our understanding of pest range limits. Int. J. Pest Manag. 2019, 65, 217–227. [Google Scholar] [CrossRef]
- Raworth, D.A. Initiation of oviposition after winter diapause in the spider mite Tetranychus urticae (Acari: Tetranychidae): Prediction and historical patterns. Popul. Ecol. 2007, 49, 201–210. [Google Scholar] [CrossRef]
- Dolatyar, S.; Jafari, S.; Pakyari, H. Effect of temperature on the development of Eotetranychus hirsti (Tetranychidae) on fig leaves. Acarologia 2015, 55, 247–254. [Google Scholar] [CrossRef]
- Rigamonti, I.E.; Trivellone, V.; Jermini, M.; Fuog, D.; Baumgaertner, J. Multiannual infestation patterns of grapevine plant inhabiting Scaphoideus titanus (Hemiptera: Cicadellidae) leafhoppers. Can. Entomol. 2014, 146, 67–79. [Google Scholar] [CrossRef] [Green Version]
- Lessio, F.; Tota, F.; Alma, A. Tracking the dispersion of Scaphoideus titanus Ball (Hemiptera: Cicadellidae) from wild to cultivated grapevine: Use of a novel mark-capture technique. Bull. Entomol. Res. 2014, 104, 432–443. [Google Scholar] [CrossRef] [PubMed]
- Riolo, P.; Minuz, R.L.; Landi, L.; Nardi, S.; Ricci, E.; Righi, M.; Isidoro, N. Population dynamics and dispersal of Scaphoideus titanus from recently recorded infested areas in central-eastern Italy. Bull. Insectol. 2014, 67, 99–107. [Google Scholar]
- Maggi, F.; Bosco, D.; Marzachì, C. Dynamics of acquisition and transmission of “flavescence dorée” phytoplasma in grapevine. Phytopathogenic Mollicutes 2014, 4, 59–71. [Google Scholar] [CrossRef]
- Jovic, J.; Riedle-Bauer, M.; Chuche, J. Vector role of cixiids and other planthopper species. In Transmission and Management of Phytoplasma-Associated Diseases; Bertaccini, A., Weintraub, P.G., Rao, G.P., Mori, N., Eds.; Springer: Singapore, 2019; Volume 2, pp. 79–114. [Google Scholar]
- Ay, J.S.; Gozlan, E. Disease dispersion as a spatial interaction: The case of Flavescence Doree. Nat. Resour. Model. 2020, 33. [Google Scholar] [CrossRef]
- Pavan, F.; Mori, N.; Bressan, S.; Mutton, P. Control strategies for grapevine phytoplasma diseases: Factors influencing the profitability of replacing symptomatic plants. Phytopathol. Mediterr. 2012, 51, 11–22. [Google Scholar]
- Rossini, L.; Severini, M.; Contarini, M.; Speranza, S. EntoSim, a ROOT-based simulator to forecast insects’ life cycle: Description and application in the case of Lobesia botrana. Crop Prot. 2020, 129. [Google Scholar] [CrossRef]
- Baumgartner, J.; Gutierrez, A.P.; Pesolillo, S.; Severini, M. A model for overwintering process of European grapevine moth Lobesia botrana (Denis & Schiffermuller) (Lepidoptera, Tortricidae) populations. J. Entomol. Acarol. Res. 2012, 44, 8–16. [Google Scholar]
- Ainseba, B.; Picart, D.; Thiery, D. An innovative multistage, physiologically structured, population model to understand the European grapevine moth dynamics. J. Math. Anal. Appl. 2011, 382, 34–46. [Google Scholar] [CrossRef]
- Almeida, R.P.P.; Nascimento, F.E.; Chau, J.; Prado, S.S.; Tsai, C.W.; Lopes, S.A.; Lopes, J.R.S. Genetic structure and biology of Xylella fastidiosa strains causing disease in citrus and coffee in Brazil. Appl. Environ. Microbiol. 2008, 74, 3690–3701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saponari, M.; Boscia, D.; Nigro, F.; Martelli, G.P. Identification of DNA sequences related to Xylella fastidiosa in oleander, almond and olive trees exhibiting leaf scorch symptoms in Apulia (Southern Italy). J. Plant Pathol. 2013, 95, 668. [Google Scholar]
- Redak, R.A.; Purcell, A.H.; Lopes, J.R.S.; Blua, M.J.; Mizell, R.F.; Andersen, P.C. The biology of xylem fluid-feeding insect vectors of Xylella fastidiosa and their relation to disease epidemiology. Annu. Rev. Entomol. 2004, 49, 243–270. [Google Scholar] [CrossRef]
- Saponari, M.; Loconsole, G.; Cornara, D.; Yokomi, R.K.; de Stradis, A.; Boscia, D.; Bosco, D.; Martelli, G.P.; Krugner, R.; Porcelli, F. Infectivity and transmission of Xylella fastidiosa by Philaenus spumarius (Hemiptera: Aphrophoridae) in Apulia, Italy. J. Econ. Entomol. 2014, 107, 1316–1319. [Google Scholar] [CrossRef] [Green Version]
- Al-Wahaibi, A.K.; Morse, J.G. Homalodisca coagulata (Hemiptera: Cicadellidae) embryonic development at constant temperatures. Fla. Entomol. 2003, 86, 477–478. [Google Scholar] [CrossRef]
- Son, Y.; Nadel, H.; Baek, S.; Johnson, M.W.; Morgan, D.J.W. Estimation of developmental parameters for adult emergence of Gonatocerus morgani, a novel egg parasitoid of the glassy-winged sharpshooter, and development of a degree-day model. Biol. Control 2012, 60, 233–240. [Google Scholar] [CrossRef]
- Mazzetto, F.; Lessio, F.; Giacosa, S.; Rolle, L.; Alma, A. Relationships between Drosophila suzukii and grapevine in North-western Italy: Seasonal presence and cultivar susceptibility. Bull. Insectol. 2020, 73, 29–38. [Google Scholar]
- de Oliveira, A.C.S.; Martins, S.G.F.; Zacarias, M.S. An individual-based model for the interaction of the mite Tetranychus urticae (Koch, 1836) with its predator Neoseiulus californicus (McGregor, 1954) (Acari: Tetranychidae, Phytoseiidae). Ecol. Model. 2013, 255, 11–20. [Google Scholar] [CrossRef]
- Nachman, G. The effects of prey patchiness, predator aggregation, and mutual interference on the functional response of Phytoseiulus persimilis feeding on Tetranychus urticae (Acari: Phytoseiidae, Tetranychidae). Exp. Appl. Acarol. 2006, 38, 87–111. [Google Scholar] [CrossRef]
- Clements, D.R.; Harmsen, R.; Clements, P.J. A mechanistic simulation to complement an empirical transition matrix model of acarine population-dynamics. Ecol. Model. 1991, 59, 257–277. [Google Scholar] [CrossRef]
- Baumgärtner, J.; Baronio, P. Phenological model of the flight of Lobesia botrana Den. & Schiif. (Lep. Tortricidae) in relation to the environmental conditions of Emilia-Romagna. Boll. Ist. Ent “G. Grandi” Univ. Bologna. 1989, 43, 157–170. [Google Scholar]
- Lessio, F.; Alma, A. Spatial distribution of nymphs of Scaphoideus titanus (Homoptera: Cicadellidae) in grapes, and evaluation of sequential sampling plans. J. Econ. Entomol. 2006, 99, 578–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ifoulis, A.A.; Savopoulou-Soultani, M. Probability distribution, sampling unit, data transformations and sequential sampling of European vine moth, Lobesia botrana (Lepidoptera: Tortricidae) larval counts from Northern Greece vineyards. Eur. J. Entomol. 2007, 104, 753–761. [Google Scholar] [CrossRef] [Green Version]
- Duchene, E.; Huard, F.; Dumas, V.; Schneider, C.; Merdinoglu, D. The challenge of adapting grapevine varieties to climate change. Clim. Res. 2010, 41, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Mosedale, J.R.; Abernethy, K.E.; Smart, R.E.; Wilson, R.J.; Maclean, I.M.D. Climate change impacts and adaptive strategies: Lessons from the grapevine. Glob. Chang. Biol. 2016, 22, 3814–3828. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Order | Species | Development and Population Dynamics | Species Distribution and Climate Change | Flight and Feeding Threshold | Spatial Distribution, Ecology | Epidemiology |
|---|---|---|---|---|---|---|
| Hemiptera | Halyomorpha halys | [31] | [32,33] | |||
| Vectors of Xylella fastidiosa | [34,35,36,37,38] | [26,39,40] | [41] | [42] | [43,44,45,46] | |
| Other leafhoppers/planthoppers | [47,48] | |||||
| Bois noir and its vectors | [49] | |||||
| Scaphoideus titanus | [14,22] | [50,51,52,53] | [54] | [20,53,55] | ||
| Mealybugs | [56] | [57] | ||||
| Lepidoptera | Lobesia botrana | [11,15,58] | [59,60,61,62] | [12,63] | ||
| Epiphyas postvittana | [64] | |||||
| Criptoblabes gnidiella | [65] | |||||
| Coleoptera | Xylotrechus arvicola | [66] | ||||
| Diptera | Drosophila suzukii | [67,68] | [69] | |||
| Acari | Spider mites | [70,71] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lessio, F.; Alma, A. Models Applied to Grapevine Pests: A Review. Insects 2021, 12, 169. https://doi.org/10.3390/insects12020169
Lessio F, Alma A. Models Applied to Grapevine Pests: A Review. Insects. 2021; 12(2):169. https://doi.org/10.3390/insects12020169
Chicago/Turabian StyleLessio, Federico, and Alberto Alma. 2021. "Models Applied to Grapevine Pests: A Review" Insects 12, no. 2: 169. https://doi.org/10.3390/insects12020169
APA StyleLessio, F., & Alma, A. (2021). Models Applied to Grapevine Pests: A Review. Insects, 12(2), 169. https://doi.org/10.3390/insects12020169