
Assessing Diversity, Plasmodium Infection and Blood Meal Sources in Mosquitoes (Diptera: Culicidae) from a Brazilian Zoological Park with Avian Malaria Transmission

 ,
, 
 , , , ,
, , , ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
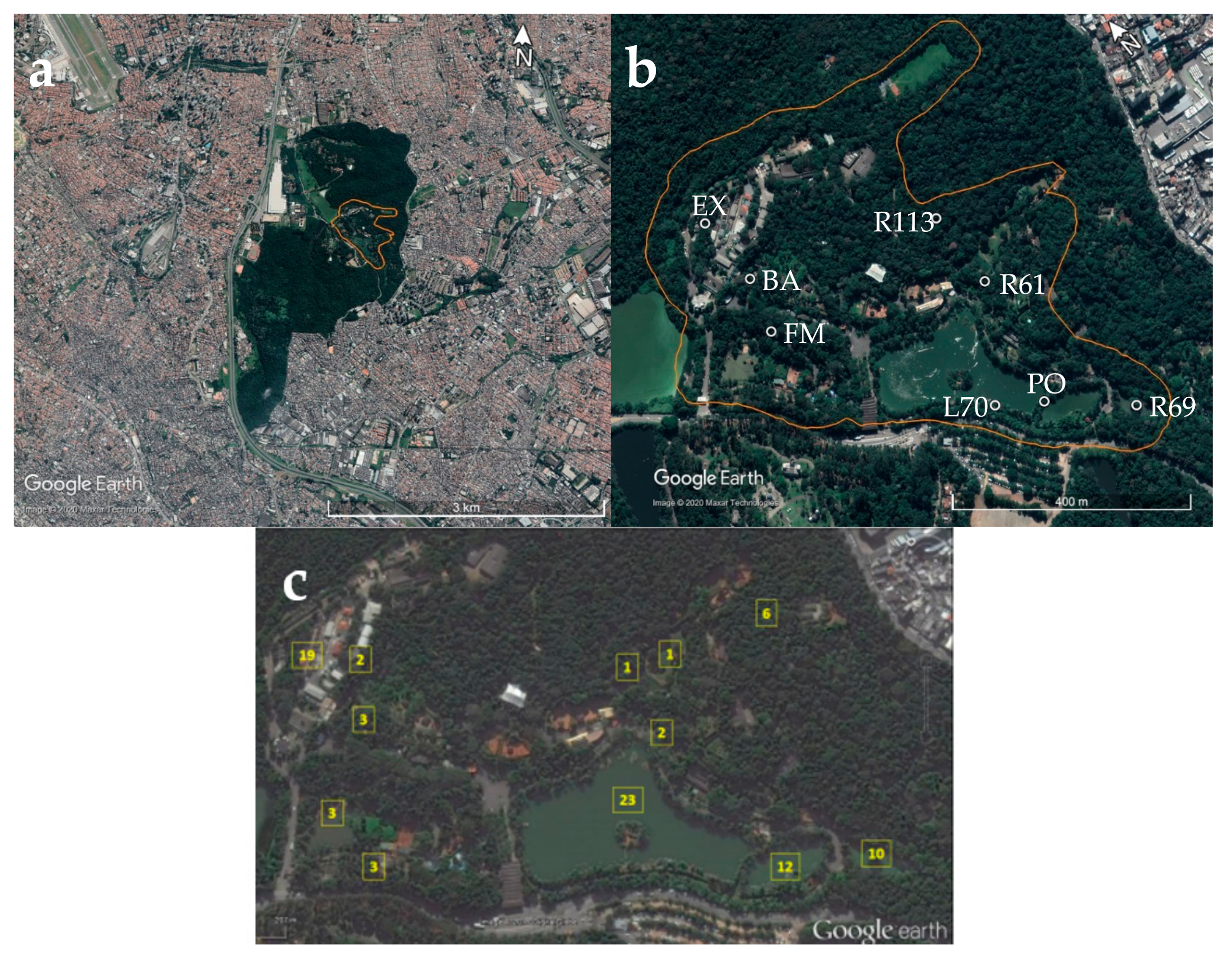
2.1. Study Area
2.2. Mosquito Collection and Identification
2.3. Plasmodium/Haemoproteus Detection
2.4. Mosquito DNA Barcoding
2.5. Blood Meal Source Identification
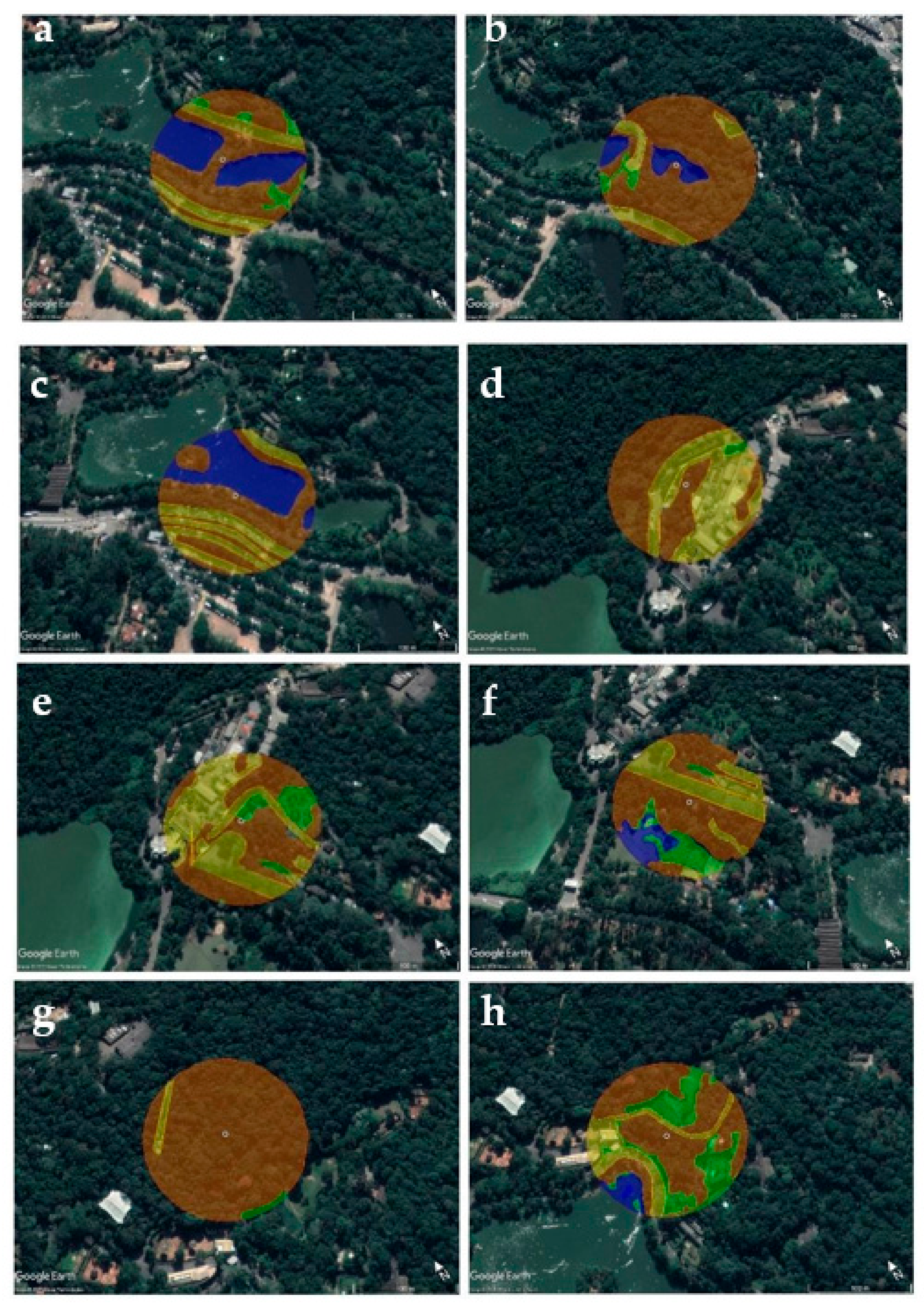
2.6. Landscape Data Analysis
2.7. Data Analysis
3. Results
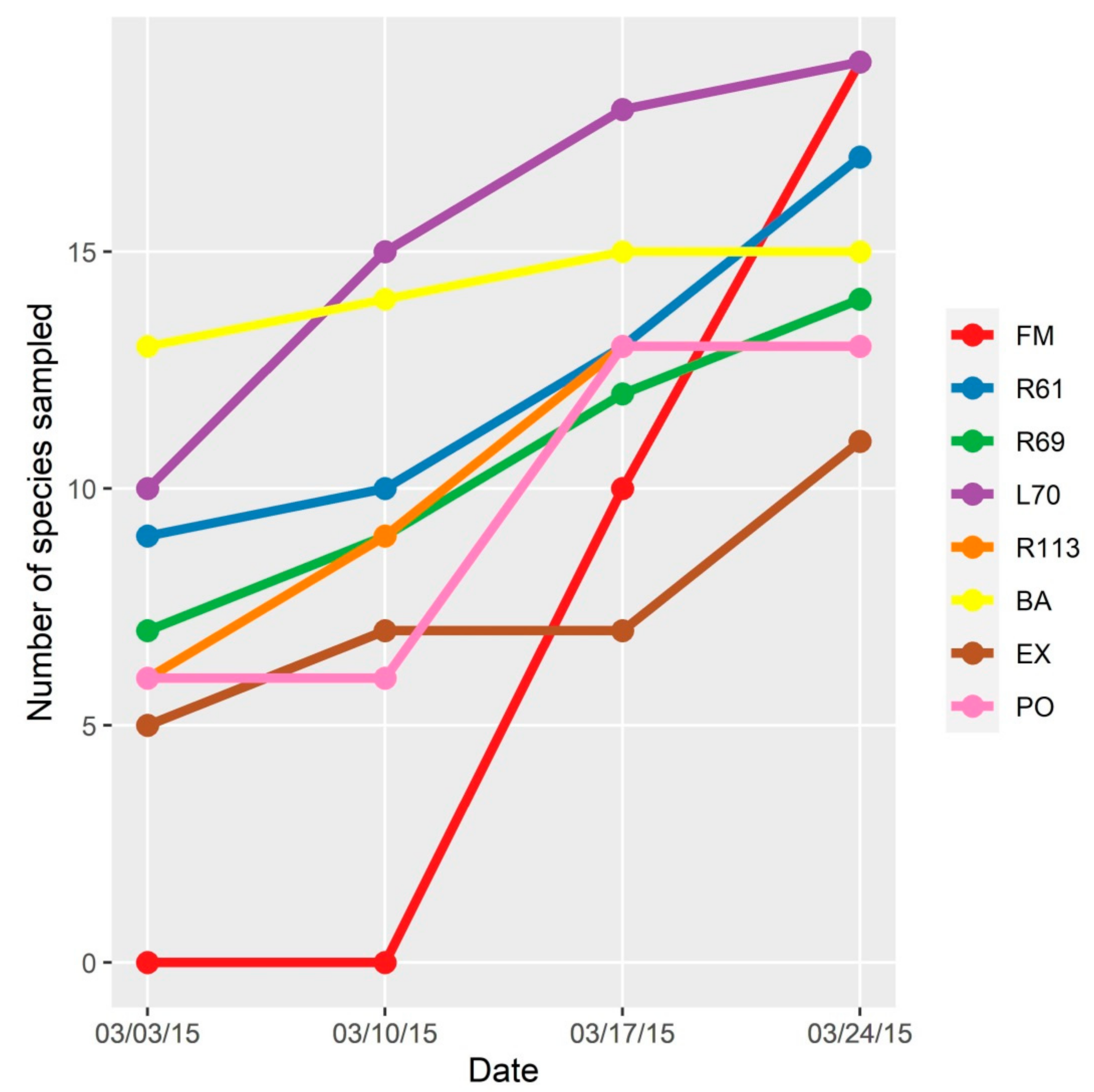
3.1. Diversity of Culicidae Species
3.2. Haemosporidian Diversity and Prevalence in Mosquitoes
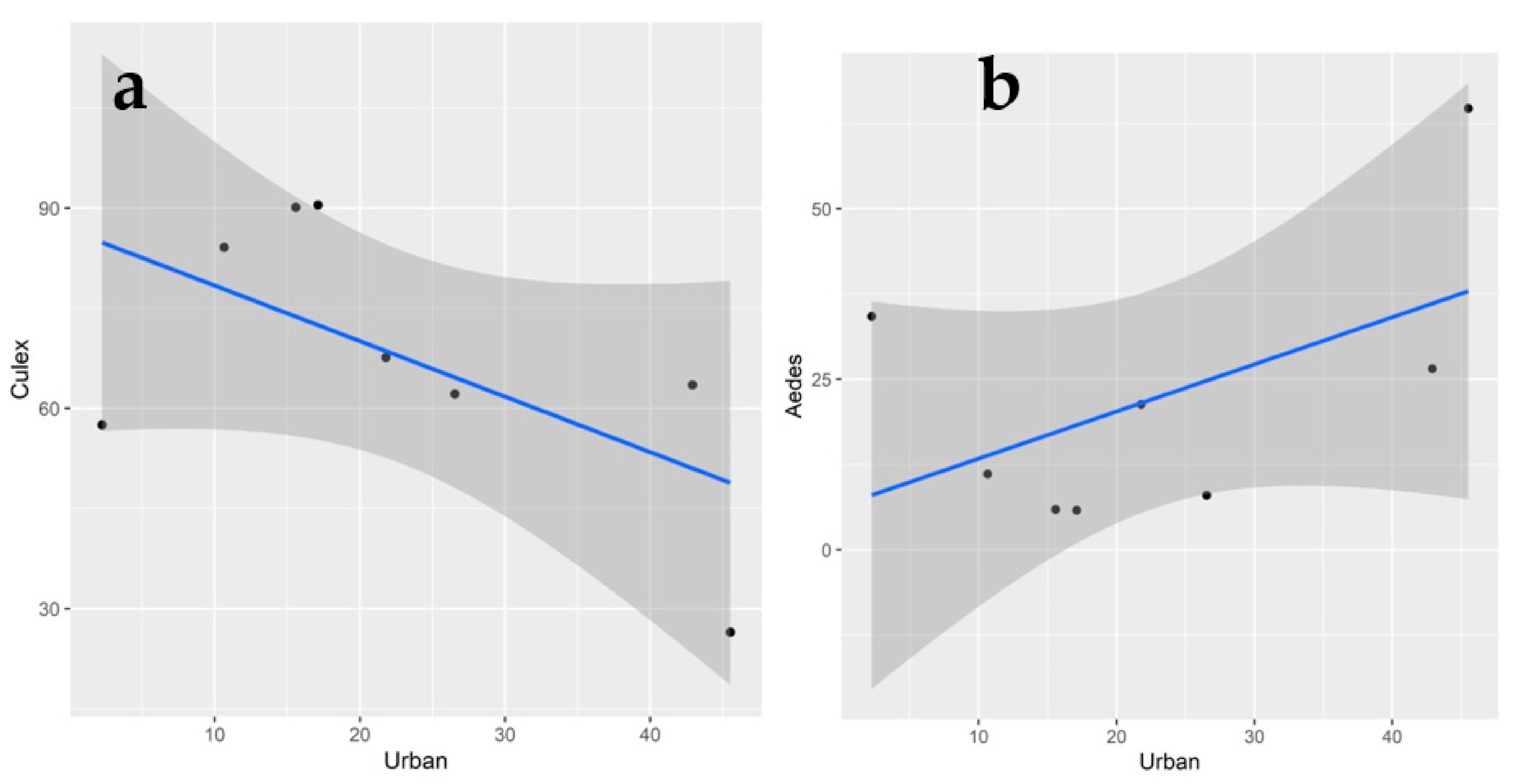
3.3. Landscape Characteristics and Mosquito Diversity
3.4. Blood Meal Source
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 03/03/2015 | 10/03/2015 | 17/03/2015 | 24/03/2015 | |
|---|---|---|---|---|
| Minimum temperature (°C) | 17.2 | 19.6 | 17.8 | 15.8 |
| Maximum temperature (°C) | 29.1 | 27.4 | 27.0 | 25.4 |
| Average temperature (°C) | 22.1 | 22.4 | 21.0 | 19.7 |
| Rainfall (mm) | 0 | 0.016 (1.6 min *) | 0.012 (0.8 min *) | 0 |
| Collection Site ID | Mosquito Species | Host Species | Number Bloodmeals Identified |
|---|---|---|---|
| BA | Aedes (Och.) scapularis | Homo sapiens | 1 |
| Culex (Cux.) chidesteri | Homo sapiens | 1 | |
| Culex (Cux.) coronator complex | Cygnus atratus | 1 | |
| Culex (Cux.) coronator complex | Homo sapiens | 1 | |
| Culex (Cux.) coronator complex | Pavo muticus | 1 | |
| Culex (Cux.) declarator | Didelphis aurita | 1 | |
| Culex (Cux.) declarator | Homo sapiens | 2 | |
| Culex (Cux.) declarator | Nycticorax nycticorax | 1 | |
| Culex (Cux.) declarator | Spizaetus ornatos | 1 | |
| EX | Mansonia (Man.) titillans | Homo sapiens | 1 |
| FM | Culex (Cux.) declarator | Canis lupus familiaris | 1 |
| Culex (Cux.) declarator | Cathartes melambrotus | 1 | |
| Culex (Cux.) declarator | Homo sapiens | 1 | |
| Culex (Cux.) declarator | Pipile jacutinga | 1 | |
| Uranotaenia (Ura.) pulcherrima | Homo sapiens | 1 | |
| Culex (Cux.) declarator | Balearica regulorum | 1 | |
| L70 | Culex (Cux.) habilitator | Anser anser | 1 |
| Culex (Mel.) Melanoconion Section | Ardea herodias | 1 | |
| Culex (Mel.) Melanoconion Section | Nycticorax nycticorax | 1 | |
| Culex (Cux.) sp. | Homo sapiens | 2 | |
| PO | Culex (Cux.) sp. | Cygnus atratus | 1 |
| Culex (Cux.) sp. | Nycticorax nycticorax | 1 | |
| Culex (Mel.) Melanoconion Section | Homo sapiens | 1 | |
| R61 | Culex (Cux.) coronator complex | Homo sapiens | 1 |
| R69 | Anopheles (Nys.) evansae | Homo sapiens | 1 |
| Culex (Cux.) ameliae | Cygnus atratus | 1 | |
| Culex (Cux.) coronator complex | Turdus rufiventris | 1 | |
| Culex (Cux.) sp. | Homo sapiens | 1 | |
| Culex (Mel.) Melanoconion Section | Rhea americana | 1 | |
| R113 | Culex (Cux.) scimitar | Homo sapiens | 1 |
References
- Valkiūnas, G. Avian Malaria Parasites and Other Haemosporidia; CRC: Boca Raton, FL, USA, 2005. [Google Scholar]
- Fecchio, A.; Chagas, C.R.F.; Bell, J.A.; Kirchgatter, K. Evolutionary ecology, taxonomy, and systematics of avian malaria and related parasites. Acta Trop. 2020, 204, 105364. [Google Scholar] [CrossRef] [PubMed]
- LaPointe, D.A.; Goff, M.L.; Atkinson, C.T. Comparative susceptibility of introduced forest-dwelling mosquitoes in Hawai’i to avian malaria, Plasmodium relictum. J Parasitol. 2005, 91, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Ishtiaq, F.; Guillaumot, L.; Clegg, S.M.; Phillimore, A.B.; Black, R.A.; Owens, I.P.F.; Mundy, N.I.; Sheldon, B.C. Avian haematozoan parasites and their associations with mosquitoes across Southwest Pacific Islands. Mol. Ecol. 2008, 17, 4545–4555. [Google Scholar] [CrossRef] [PubMed]
- Njabo, K.Y.; Cornel, A.J.; Sehgal, R.N.; Loiseau, C.; Buermann, W.; Harrigan, R.J.; Pollinger, J.; Valkiūnas, G.; Smith, T.B. Coquillettidia (Culicidae, Diptera) mosquitoes are natural vectors of avian malaria in Africa. Malar. J. 2009, 8, 193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santiago-Alarcon, D.; Palinauskas, V.; Schaefer, H.M. Diptera vectors of avian Haemosporidian parasites: Untangling parasite life cycles and their taxonomy. Biol. Rev. Camb. Philos. Soc. 2012, 87, 928–964. [Google Scholar] [CrossRef] [PubMed]
- Clark, N.J.; Clegg, S.M.; Lima, M.R. A review of global diversity in avian haemosporidians (Plasmodium and Haemoproteus: Haemosporida): New insights from molecular data. Int. J. Parasitol. 2014, 44, 329–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atkinson, C.T.; Thomas, N.J.; Hunter, D.B. Parasitic Diseases of Wild Birds; Wiley-Blackwell: Oxford, UK, 2008. [Google Scholar]
- Chagas, C.R.F.; Valkiūnas, G.; Nery, C.V.C.; Henrique, P.C.; Gonzalez, I.H.L.; Monteiro, E.F.; Guimarães, L.O.; Romanod, C.M.; Kirchgatter, K. Plasmodium (Novyella) nucleophilum from an Egyptian Goose in São Paulo Zoo, Brazil: Microscopic confirmation and molecular characterization. Int. J. Parasitol. Parasites Wildl. 2013, 2, 286–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panayotova-Pencheva, M.S. Parasites in captive animals: A review of studies in some European zoos. Zool. Garten. 2013, 82, 60–71. [Google Scholar] [CrossRef]
- Bueno, M.G.; Lopez, R.P.; de Menezes, R.M.; Costa-Nascimento, M.J.; Lima, G.F.M.C.; Araújo, R.A.S.; Guida, F.J.V.; Kirchgatter, K. Identification of Plasmodium relictum causing mortality in penguins (Spheniscus magellanicus) from São Paulo Zoo, Brazil. Vet. Parasitol. 2010, 173, 123–127. [Google Scholar] [CrossRef]
- Chagas, C.R.; Guimarães, L.d.O.; Monteiro, E.F.; Valkiūnas, G.; Katayama, M.V.; Santos, S.V.; Guida, F.J.; Simões, R.F.; Kirchgatter, K. Hemosporidian parasites of free-living birds in the São Paulo Zoo, Brazil. Parasitol. Res. 2016, 115, 1443–1452. [Google Scholar] [CrossRef] [PubMed]
- Chagas, C.R.; Valkiūnas, G.; de Oliveira Guimarães, L.; Monteiro, E.F.; Guida, F.J.; Simões, R.F.; Rodrigues, P.T.; de Albuquerque Luna, E.J.; Kirchgatter, K. Diversity and distribution of avian malaria and related haemosporidian parasites in captive birds from a Brazilian megalopolis. Malar. J. 2017, 17, 83. [Google Scholar] [CrossRef]
- Nasci, R.S. A lightweight battery-powered aspirator for collecting resting mosquitoes in the field. Mosq. News 1981, 41, 808–811. [Google Scholar]
- Sudia, W.D.; Chamberlain, R.W. Battery-operated light trap, an improved model. Mosq. News 1962, 22, 126–129. [Google Scholar]
- Lane, J. Neotropical Culicidae; University of São Paulo: São Paulo, Brazil, 1953; Volume 2, 1112p. [Google Scholar]
- Forattini, O.P. Breve nota sobre variações em espécies de Anopheles (Diptera: Culicidae). Rev. Bras. Ent. 1964, 11, 139–140. [Google Scholar]
- Forattini, O.P. Culicidologia Médica. Identificação, Biologia, Epidemiologia; Edusp: São Paulo, Brazil, 2002; Volume 2, 860p. [Google Scholar]
- Hellgren, O.; Waldenström, J.; Bensch, S. A new PCR assay for simultaneous studies of Leucocytozoon, Plasmodium, and Haemoproteus from avian blood. J. Parasitol. 2004, 90, 797–802. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Bensch, S.; Hellgren, O.; Pérez-Tris, J. MalAvi: A public database of malaria parasites and related haemosporidians in avian hosts based on mitochondrial cytochrome b lineages. Mol. Ecol. Resour. 2009, 9, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Ruiz, F.; Linton, Y.M.; Ponsonby, D.J.; Conn, J.E.; Herrera, M.; Quiñones, M.L.; Vélez, I.D.; Wilkerson, R.C. Molecular comparison of topotypic specimens confirms Anopheles (Nyssorhynchus) dunhami Causey (Diptera: Culicidae) in the Colombian Amazon. Mem. Inst. Oswaldo Cruz 2010, 105, 899–903. [Google Scholar] [CrossRef] [PubMed]
- Meier, R.; Shiyang, K.; Vaidya, G.; Ng, P.K.L. DNA barcoding and taxonomy in Diptera: A tale of high intraspecific variability and low identification success. Syst. Biol. 2006, 55, 715–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kocher, T.D.; Thomas, W.K.; Meyer, A.; Edwards, S.V.; Pääbo, S.; Villablanca, F.X.; Wilson, A.C. Dynamics of mitochondrial DNA evolution in animals: Amplification and sequencing with conserved primers. Proc. Natl. Acad. Sci. USA 1989, 86, 6196–6200. [Google Scholar] [CrossRef] [Green Version]
- Kirchgatter, K.; Tubaki, R.M.; Malafronte Rdos, S.; Alves, I.C.; Lima, G.F.M.C.; Guimarães, L.O.; Zampaulo, R.A.; Wunderlich, G. Anopheles (Kerteszia) cruzii (Diptera: Culicidae) in peridomiciliary area during asymptomatic malaria transmission in the Atlantic Forest: Molecular identification of blood-meal sources indicates humans as primary intermediate hosts. Rev. Inst. Med. Trop. São Paulo 2014, 56, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Magurran, A.E. Measuring Biological Diversity; Blackwell: Oxford, UK, 2004. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria. Available online: http://www.R-project.org/ (accessed on 19 October 2020).
- Baliton, R.S.; Wulandari, C.; Landicho, L.; Cabahug, R.E.D.; Paelmo, R.F.; Comia, R.A.; Visco, R.G.; Budiono, P.; Herwanti, S.; Rusita, R.; et al. Ecological services of agroforestry landscapes in selected watershed areas in the Philippines and Indonesia. Biotropia 2017, 24, 71–84. [Google Scholar]
- Abella-Medrano, C.A.; Ibáñez-Bernal, S.; Mac Gregor-Fors, I.; Santiago-Alarcon, D. Spatiotemporal variation of mosquito diversity (Diptera: Culicidae) at places with different land-use types within a neotropical montane cloud forest matrix. Parasit Vectors 2015, 24, 487. [Google Scholar] [CrossRef] [Green Version]
- Loaiza, J.R.; Rovira, J.R.; Sanjur, O.I.; Zepeda, J.A.; Pecor, J.E.; Foley, D.H.; Dutari, L.; Radtke, M.; Pongsiri, M.J.; Molinar, O.S.; et al. Forest disturbance and vector transmitted diseases in the lowland tropical rainforest of central Panama. Trop. Med. Int. Health 2019, 24, 849–861. [Google Scholar] [CrossRef] [PubMed]
- Ferraguti, M.; Martínez de la Puente, J.; Muñoz, J.; Roiz, D.; Ruiz, S.; Soriguer, R.; Figuerola, J. Avian Plasmodium in Culex and Ochlerotatus Mosquitoes from Southern Spain: Effects of Season and Host-Feeding Source on Parasite Dynamics. PLoS ONE 2013, 18, e66237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takken, W.; Verhulst, N.O. Host preferences of blood-feeding mosquitoes. Annu. Rev. Entomol. 2013, 58, 433–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez de la Puente, J.; Ferraguti, M.; Ruiz, S.; Roiz, D.; Soriguer, R.C.; Figuerola, J. Culex pipiens forms and urbanization: Effects on blood feeding sources and transmission of avian Plasmodium. Malar. J. 2016, 15, 589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Žiegytė, R.; Valkiūnas, G. Recent advances in vector studies of avian haemosporidian parasites. Ekologija 2014, 60, 73–83. [Google Scholar]
- Martínez de la Puente, J.; Muñoz, J.; Capelli, G.; Montarsi, F.; Soriguer, R.; Arnoldi, D.; Rizzoli, A.; Figuerola, J. Avian malaria parasites in the last supper: Identifying encounters between parasites and the invasive Asian mosquito tiger and native mosquito species in Italy. Malar. J. 2015, 28, 14–32. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, F.C., Jr.; Rodrigues, R.A.; Sato, Y.; Borges, M.A.; Braga, É.M. Searching for putative avian malaria vectors in a Seasonally Dry Tropical Forest in Brazil. Parasit Vectors 2016, 9, 587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bukauskaitė, D.; Iezhova, T.A.; Ilgūnas, M.; Valkiūnas, G. High susceptibility of the laboratory-reared biting midges Culicoides nubeculosus to Haemoproteus infections, with review on Culicoides species that transmit avian haemoproteids. Parasitology 2019, 146, 333–341. [Google Scholar] [CrossRef]
- Njabo, K.Y.; Cornel, A.J.; Bonneaud, C.; Toffelmier, E.; Sehgal, R.N.M.; Valkiūnas, G.; Russell, A.F.; Smith, T.B. Nonspecific patterns of vector, host and avian malaria parasite associations in a central African rainforest. Mol. Ecol. 2011, 20, 1049–1061. [Google Scholar] [CrossRef] [Green Version]
- Valkiūnas, G.; Kazlauskienė, R.; Bernotienė, R.; Palinauskas, V.; Iezhova, T.A. Abortive long-lasting sporogony of two Haemoproteus species (Haemosporida, Haemoproteidae) in the mosquito Ochlerotatus cantans, with perspectives on haemosporidian vector research. Parasitol. Res. 2013, 112, 2159–2169. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez-López, R.; Martínez de la Puente, J.; Gangoso, L.; Yan, J.; Soriguer, R.C.; Figuerola, J. Do mosquitoes transmit the avian malaria-like parasite Haemoproteus? An experimental test of vector competence using mosquito saliva. Parasit Vectors 2016, 28, 609. [Google Scholar] [CrossRef] [Green Version]
- Valkiūnas, G. Haemosporidian vector research: Marriage of molecular and microscopical approaches is essential. Mol. Ecol. 2011, 20, 3084–3086. [Google Scholar] [CrossRef] [PubMed]
- Perrella, D.F.; Ferrari, D.S.; Katayama, M.V.; Paiva, R.V.; Guida, F.J. A Avifauna do Parque Estadual das Fontes do Ipiranga, um remanescente de Mata Atlântica imerso na área urbana de São Paulo, SP. Ornithologia 2018, 10, 4–16. [Google Scholar]
- Anjos, C.C.; Chagas, C.R.F.; Fecchio, A.; Schunck, F.; Costa-Nascimento, M.J.; Monteiro, E.F.; Mathias, B.S.; Bell, J.A.; Guimarães, L.O.; Comiche, K.J.M.; et al. Avian malaria and related parasites from resident and migratory birds in the Brazilian Atlantic Forest, with description of a new Haemoproteus species. Pathogens 2021, 10, 103. [Google Scholar] [CrossRef] [PubMed]
- Mantilla, J.S.; González, A.D.; Valkiūnas, G.; Moncada, L.I.; Matta, N.E. Description and molecular characterization of Plasmodium (Novyella) unalis sp. nov. from the Great Thrush (Turdus fuscater) in highland of Colombia. Parasitol. Res. 2013, 112, 4193–4204. [Google Scholar] [CrossRef] [PubMed]
- Fecchio, A.; Ellis, V.A.; Bell, J.A.; Andretti, C.B.; D’Horta, F.M.; Silva, A.M.; Tkach, V.V.; Weckstein, J.D. Avian malaria, ecological host traits and mosquito abundance in southeastern Amazonia. Parasitology 2017, 144, 1117–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fecchio, A.; Wells, K.; Bell, J.A.; Tkach, V.V.; Lutz, H.L.; Weckstein, J.D.; Clegg, S.M.; Clark, N.J. Climate variation influences host specificity in avian malaria parasites. Ecol. Lett. 2019, 22, 547–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- International Union for Conservation of Nature and National Resources. The IUCN Red List of Threatened Species. Version 2020-2. Available online: https://www.iucnredlist.org./ (accessed on 19 October 2020).




| Mosquito Species (or Species Groups) | Total Collected | Total Analyzed | Individual Analyses | Positive Samples | Pools | Total Samples in Pools | Positive Pools |
|---|---|---|---|---|---|---|---|
| Aedes (Och.) scapularis | 294 | 294 | 160 | 5 (1.7) | 16 | 134 | - |
| Aedes (Stg.) albopictus | 15 | 15 | 6 | 1 (6.6) | 2 | 9 | - |
| Anopheles (Nys.) evansae | 33 | 33 | 8 | - | 3 | 25 | - |
| Anopheles (Nys.) galvaoi | 3 | 3 | 3 | - | - | - | - |
| Anopheles (Nys.) spp. | 5 | 5 | 5 | - | - | - | - |
| Anopheles (Nys.) strodei | 22 | 22 | 6 | - | 2 | 16 | - |
| Coquilettidia (Rhy.) nigricans | 1 | 1 | 1 | - | - | - | - |
| Coquillettidia (Rhy.) hermanoi | 2 | 2 | - | - | 1 | 2 | - |
| Coquillettidia (Rhy.) sp. | 1 | 1 | 1 | - | - | - | - |
| Culex (Cux.) acharistus | 8 | 8 | 5 | 1 (12.5) | 1 | 3 | 1 (12.5) |
| Culex (Cux.) ameliae | 65 | 65 | 10 | - | 7 | 55 | - |
| Culex (Cux.) bidens | 209 | 206 | 21 | - | 20 | 185 | - |
| Culex (Cux.) chidesteri | 36 | 35 | 6 | - | 4 | 29 | - |
| Culex (Cux.) coronator complex | 225 | 221 | 56 | - | 19 | 165 | 2 (0.90) |
| Culex (Cux.) declarator | 136 | 135 | 41 | - | 11 | 94 | - |
| Culex (Cux.) dolosus | 5 | 5 | 5 | 1 (20.0) | - | - | - |
| Culex (Cux.) habilitator | 66 | 64 | 49 | - | 2 | 15 | - |
| Culex (Cux.) nigripalpus | 10 | 10 | 5 | - | 2 | 5 | - |
| Culex (Cux.) quinquefasciatus | 1 | 1 | 1 | - | - | - | - |
| Culex (Cux.) saltanensis | 2 | 2 | - | - | 1 | 2 | - |
| Culex (Cux.) scimitar | 28 | 28 | - | - | 3 | 28 | - |
| Culex (Cux.) spp. | 529 | 9 | 9 | - | - | - | - |
| Culex (Mcx.) imitator | 5 | 5 | - | - | 1 | 5 | - |
| Culex (Mel.) Melanoconion Section | 220 | 219 | 61 | - | 19 | 158 | - |
| Limatus durhamii | 1 | 1 | 1 | - | - | - | - |
| Mansonia (Man.) indubitans | 65 | 65 | 21 | 1 (1.55) | 6 | 44 | 1 (1.55) |
| Mansonia (Man.) pseudotitillans | 18 | 17 | 5 | - | 2 | 12 | - |
| Mansonia (Man.) spp. | 5 | 5 | 5 | - | - | - | - |
| Mansonia (Man.) titillans | 25 | 25 | 14 | - | 2 | 11 | - |
| Uranotaenia (Ura.) pulcherrima | 4 | 4 | - | - | 1 | 4 | - |
| Total | 2039 | 1506 | 505 | 9 (0.60) | 125 | 1001 | 4 (0.26) |
| ID Positive Sample | ID Mosquito | Species (According Table 1) | Species (According DNA Barcode) # | Collection Day/Month | Collection Site | Parasite/ Lineage/GenBank Accession |
|---|---|---|---|---|---|---|
| 12 | 12 | Aedes (Stg.) albopictus | Aedes (Stg.) albopictus | 03/03 | EX | P. nucleophilum/ pDENPET03/MW492356 |
| 16 | 16 | Culex (Cux.) acharistus | Culex (Cux.) coronator | 03/03 | EX | P. nucleophilum/ pDENPET03/MW492357 |
| 44 | 44 | Mansonia (Man.) indubitans | Mansonia (Man.) sp. † | 03/03 | R113 | P. nucleophilum/ pDENPET03/MW492358 |
| 60 | 60 | Aedes (Och.) scapularis | Aedes (Och.) scapularis | 03/03 | BA | P. nucleophilum/ pDENPET03/MW492359 |
| 67 | 67 | Aedes (Och.) scapularis | Aedes (Och.) scapularis | 03/03 | BA | P. nucleophilum/ pDENPET03/MW492360 |
| 76 | 76 | Aedes (Och.) scapularis | Aedes (Och.) scapularis | 03/03 | BA | P. nucleophilum/ pDENPET03/MW492361 |
| 87 | 87 | Aedes (Och.) scapularis | Aedes (Och.) scapularis | 03/03 | BA | P. nucleophilum/ pDENPET03/MW492362 |
| 121 | 121 | Culex (Cux.) dolosus aff. | Culex (Cux.) maxi | 10/03 | EX | Plasmodium sp./ pCULMAX01/MW492363 |
| 884 * | 884 * | Aedes (Och.) scapularis | Aedes (Och.) scapularis | 30/06 | BA | Haemoproteus (Parahaemoproteus) sp./ hAEDSCA01/MW492364 |
| Pool 36 | 887–889 | Culex (Cux.) acharistus | Culex (Cux.) nigripalpus | 17/03 | R61 | Plasmodium sp./ pVIOLI03/MW492365 |
| Pool 103 | 634 | Mansonia (Man.) indubitans | Mansonia (Man.) spp. † | 17/03 | R61 | Haemoproteus (Parahaemoproteus) sp./ hMYISWA01/MW492366 |
| 726 | 24/03 | R113 | ||||
| 798–800 | 10/03 | L70 | ||||
| Pool 119 | 947–954 | Culex (Cux.) coronator complex | Culex (Cux.) spp. † | 03/03 | BA | Plasmodium sp./ pCULEX05/MW492367 |
| Pool 135 | 461 | Culex (Cux.) coronator complex | Culex (Cux.) spp. † | 17/03 | R61 | Plasmodium sp./ pCULEX06/MW492368 |
| 463 | ||||||
| 630–633 |
| Site | Forest Area Edge Length (m) | Forest Area (High Trees) (m2) | Lake (m2) | Edification (m2) | Paving (m2) | Bush/Sand/Grass (m2) | Unclassified (m2) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PO | 1647 | 12,556 | (49.3) | 7034 | (27.6) | 0 | (0) | 4355 | (17.1) | 1277 | (5.0) | 224 | (0.9) |
| R69 | 1340 | 19,495 | (76.6) | 2295 | (9.0) | 0 | (0) | 2709 | (10.6) | 633 | (2.5) | 314 | (1.2) |
| L70 | 1792 | 10,928 | (42.9) | 8247 | (32.4) | 0 | (0) | 5541 | (21.8) | 0 | (0) | 730 | (2.9) |
| EX | 1093 | 13,382 | (52.6) | 0 | (0) | 7438 | (29.2) | 4142 | (16.3) | 285 | (1.1) | 199 | (0.8) |
| BA | 1832 | 10,972 | (43.1) | 0 | (0) | 5348 | (21.0) | 5565 | (21.9) | 2302 | (9.0) | 1256 | (4.9) |
| FM | 1305 | 12,323 | (48.4) | 2006 | (7.9) | 220 | (0.9) | 6531 | (25.7) | 2370 | (9.3) | 1996 | (7.8) |
| R113 | 181 | 24,530 | (96.4) | 0 | (0) | 0 | (0) | 569 | (2.2) | 328 | (1.3) | 19 | (0.1) |
| R61 | 1602 | 14,144 | (55.6) | 963 | (3.8) | 1533 | (6.0) | 2432 | (9.6) | 6124 | (24.1) | 249 | (1.0) |
| Vertebrate Hosts | Ae. scapularis | An. evansae | Cx. ameliae | Cx. chidesteri | Cx. coronator complex | Cx. declarator | Cx. habilitator | Cx. scimitar | Cx. (Mel.) Melanoconion Section | Cx. (Cux.) spp. | Ma. titillans | Ur. pulcherrima | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Family | Host Species | ||||||||||||
| Birds | 100% | 60% | 50% | 100% | 75% | 40% | |||||||
| Accipitridae | Spizaetus ornatos | 1 (BA) | |||||||||||
| Anatidae | Anser anser | 1 (L70) | |||||||||||
| Cygnus atratus | 1 (R69) | 1 (BA) | 1 (PO) | ||||||||||
| Ardeidae | Ardea herodias | 1 (L70) | |||||||||||
| Nycticorax nycticorax | 1 (BA) | 1 (L70) | 1 (PO) | ||||||||||
| Cathartidae | Cathartes melambrotus | 1 (FM) | |||||||||||
| Cracidae | Pipile jacutinga | 1 (FM) | |||||||||||
| Gruidae | Balearica regulorum | 1(FM) | |||||||||||
| Phasianidae | Pavo muticus | 1 (BA) | |||||||||||
| Rheidae | Rhea americana | 1 (R69) | |||||||||||
| Turdidae | Turdus rufiventris | 1 (R69) | |||||||||||
| Mammals | 100% | 100% | 100% | 40% | 50% | 100% | 25% | 60% | 100% | 100% | |||
| Canidae | Canis lupus familiaris | 1 (FM) | |||||||||||
| Didelphidae | Didelphis aurita | 1 (BA) | |||||||||||
| Hominídea | Homo sapiens | 1 (BA) | 1 (R69) | 1 (BA) | 1(BA) 1(R61) | 2 (BA) 1 (FM) | 1 (R113) | 1 (PO) | 2 (L70) 1 (R69) | 1 (EX) | 1 (FM) | ||
| Total | 1 | 1 | 1 | 1 | 5 | 10 | 1 | 1 | 4 | 5 | 1 | 1 | |
| BA, PO, R69, L70, R61, R113, EX, and FM are sampling sites as described in the Materials and Methods section. | |||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guimarães, L.d.O.; Simões, R.F.; Chagas, C.R.F.; Menezes, R.M.T.d.; Silva, F.S.; Monteiro, E.F.; Holcman, M.M.; Bajay, M.M.; Pinter, A.; Camargo-Neves, V.L.F.d.; et al. Assessing Diversity, Plasmodium Infection and Blood Meal Sources in Mosquitoes (Diptera: Culicidae) from a Brazilian Zoological Park with Avian Malaria Transmission. Insects 2021, 12, 215. https://doi.org/10.3390/insects12030215
Guimarães LdO, Simões RF, Chagas CRF, Menezes RMTd, Silva FS, Monteiro EF, Holcman MM, Bajay MM, Pinter A, Camargo-Neves VLFd, et al. Assessing Diversity, Plasmodium Infection and Blood Meal Sources in Mosquitoes (Diptera: Culicidae) from a Brazilian Zoological Park with Avian Malaria Transmission. Insects. 2021; 12(3):215. https://doi.org/10.3390/insects12030215
Chicago/Turabian StyleGuimarães, Lilian de Oliveira, Roseli França Simões, Carolina Romeiro Fernandes Chagas, Regiane Maria Tironi de Menezes, Fabiana Santos Silva, Eliana Ferreira Monteiro, Marcia Moreira Holcman, Miklos Maximiliano Bajay, Adriano Pinter, Vera Lucia Fonseca de Camargo-Neves, and et al. 2021. "Assessing Diversity, Plasmodium Infection and Blood Meal Sources in Mosquitoes (Diptera: Culicidae) from a Brazilian Zoological Park with Avian Malaria Transmission" Insects 12, no. 3: 215. https://doi.org/10.3390/insects12030215
APA StyleGuimarães, L. d. O., Simões, R. F., Chagas, C. R. F., Menezes, R. M. T. d., Silva, F. S., Monteiro, E. F., Holcman, M. M., Bajay, M. M., Pinter, A., Camargo-Neves, V. L. F. d., & Kirchgatter, K. (2021). Assessing Diversity, Plasmodium Infection and Blood Meal Sources in Mosquitoes (Diptera: Culicidae) from a Brazilian Zoological Park with Avian Malaria Transmission. Insects, 12(3), 215. https://doi.org/10.3390/insects12030215