
Assessment of Oil Palm Pollinating Weevil (Elaeidobius kamerunicus) Population Density in Biparental dura × pisifera Hybrids on Deep Peat-Soil in Perak State, Malaysia

 ,
,  ,
,  , ,
, ,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Planting Materials
2.2. Experimental Design and Research Location
2.3. Procedure and Data Collection on the Elaeidobius kamerunicus Population Density in Oil Palm Hybrids
2.4. Data Analysis
3. Results
3.1. ANOVA and Variance Component of the Elaeidobius kamerunicus Population on Each Day of Anthesis
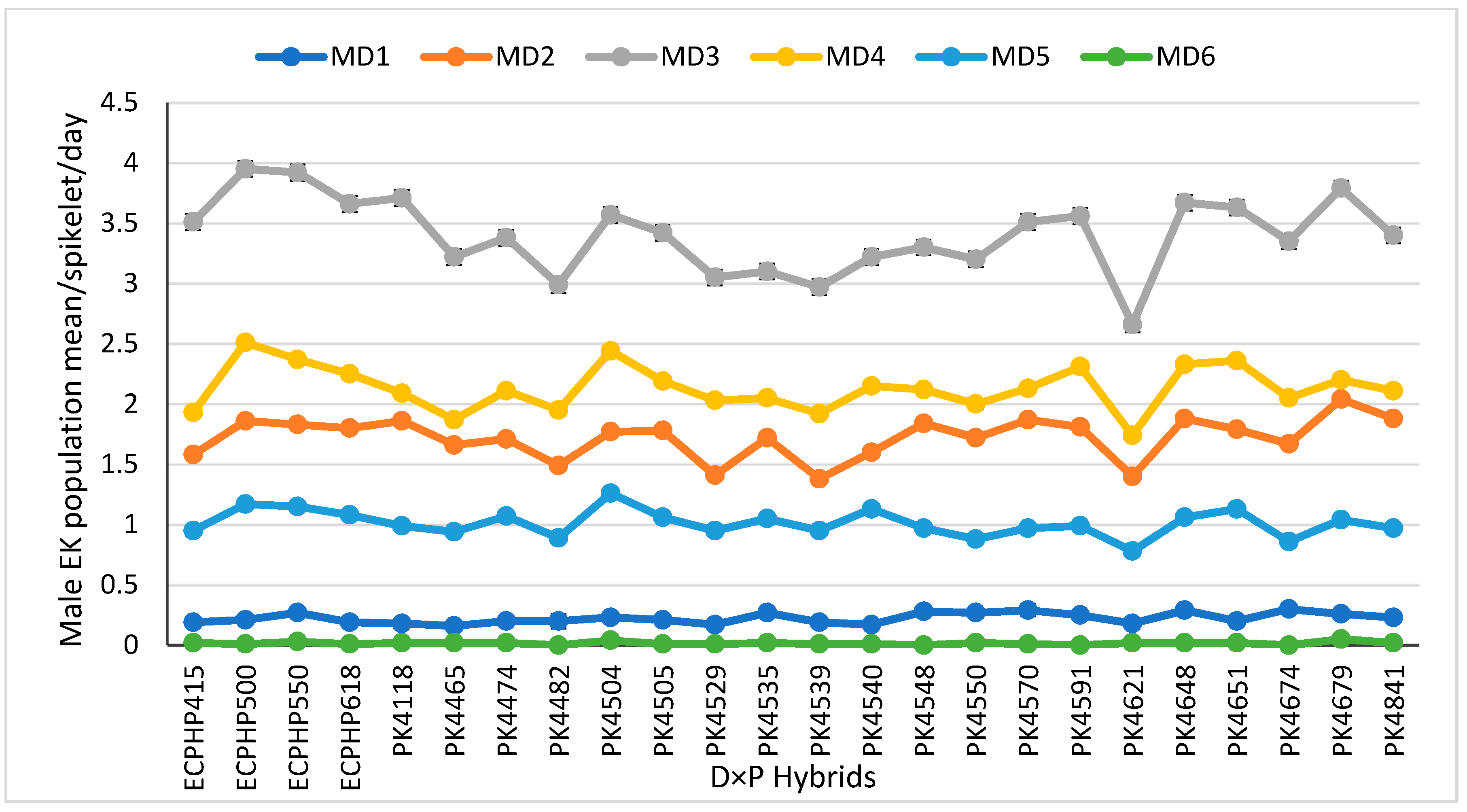
3.1.1. Daily Population Mean ± Standard Error of Male Elaeidobius kamerunicus per Male Inflorescence Spikelet at Each Day of Anthesis
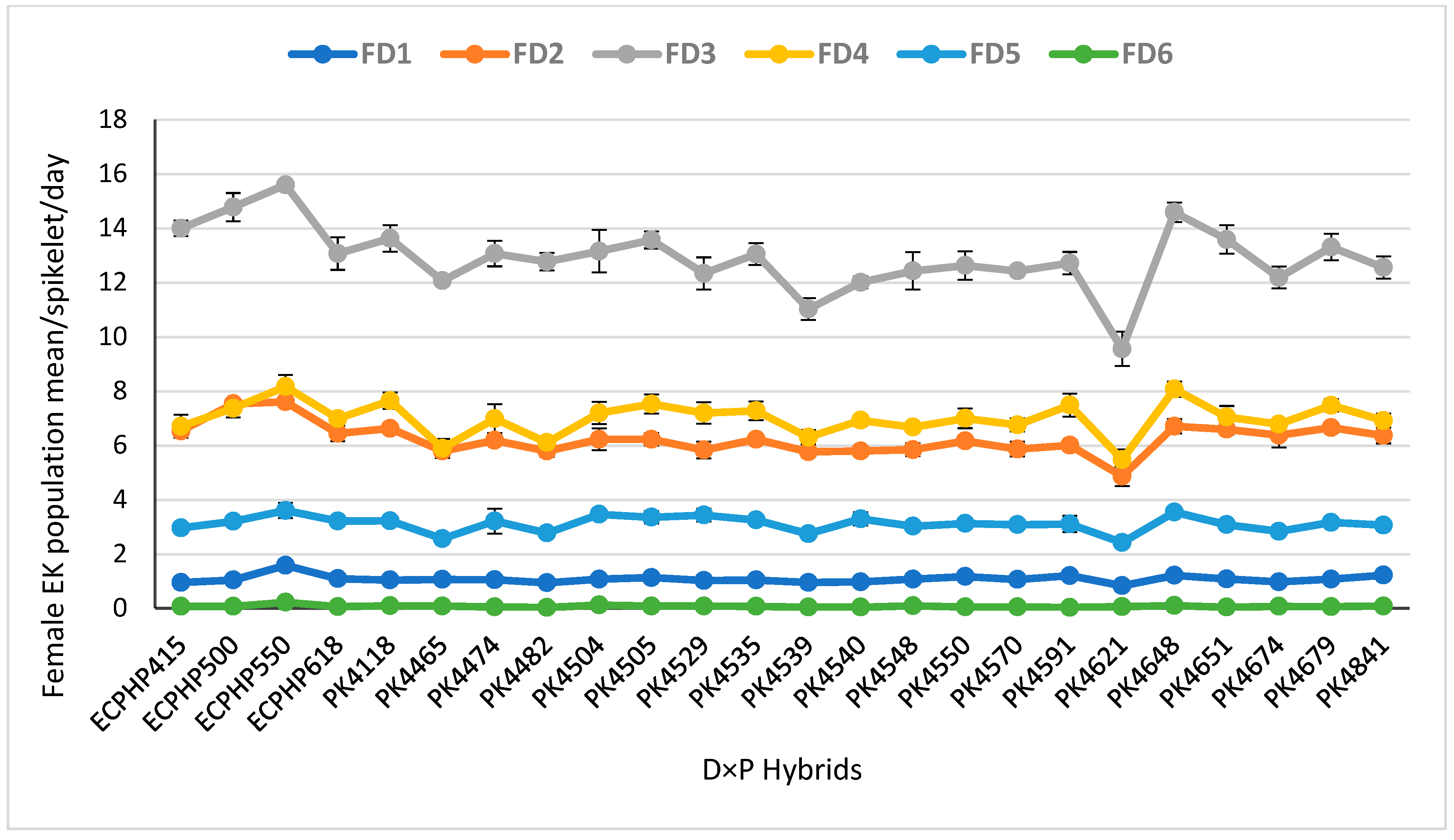
3.1.2. Daily Population Mean ± Standard Error of Female E. kamerunicus per Spikelet
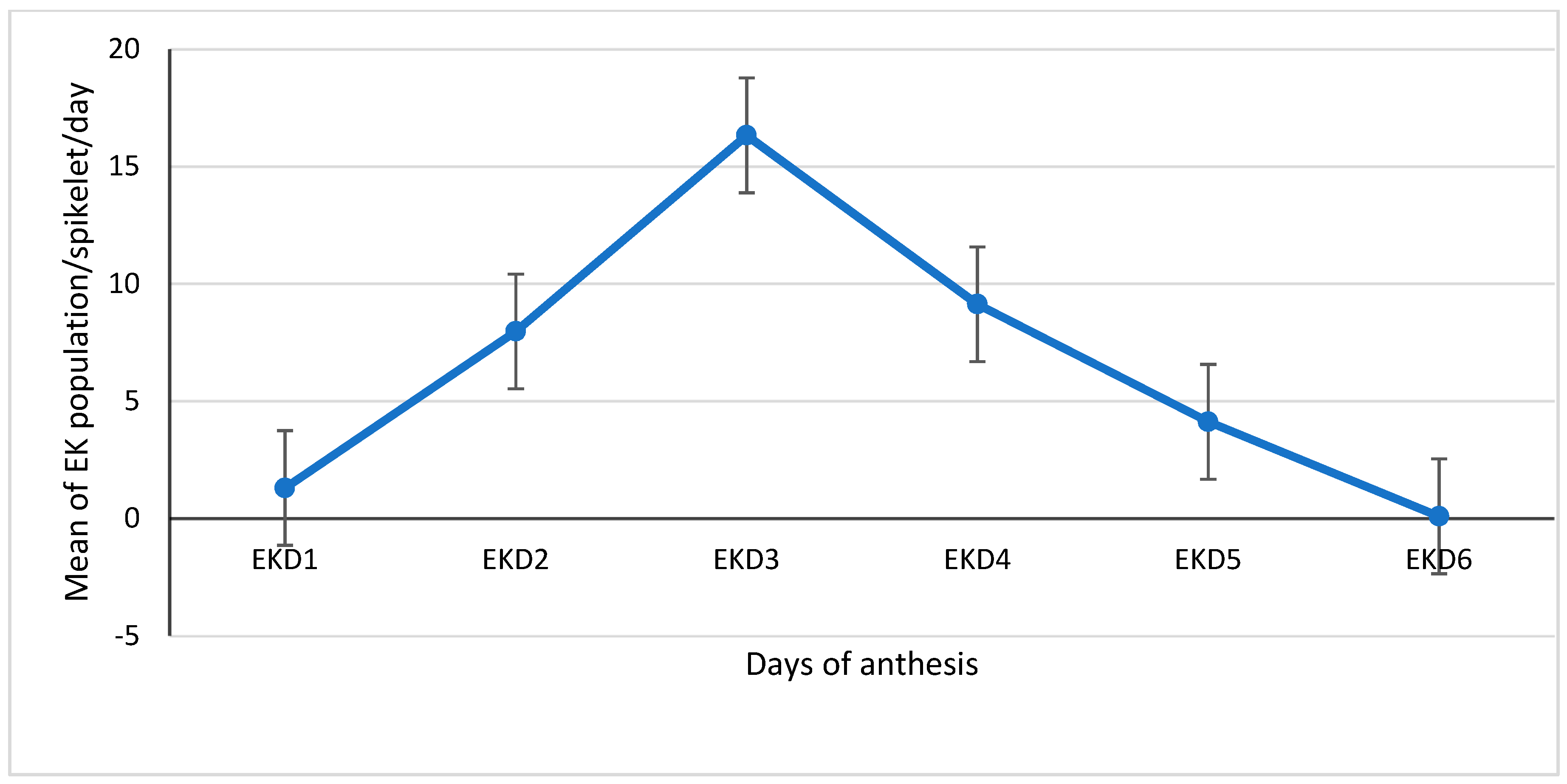
3.1.3. Daily Population Means of E. kamerunicus per Spikelet of Each Anthesis Days (1–6)
3.2. Monthly Population Mean of E. kamerunicus per Spikelet and its Population Density per Hybrid
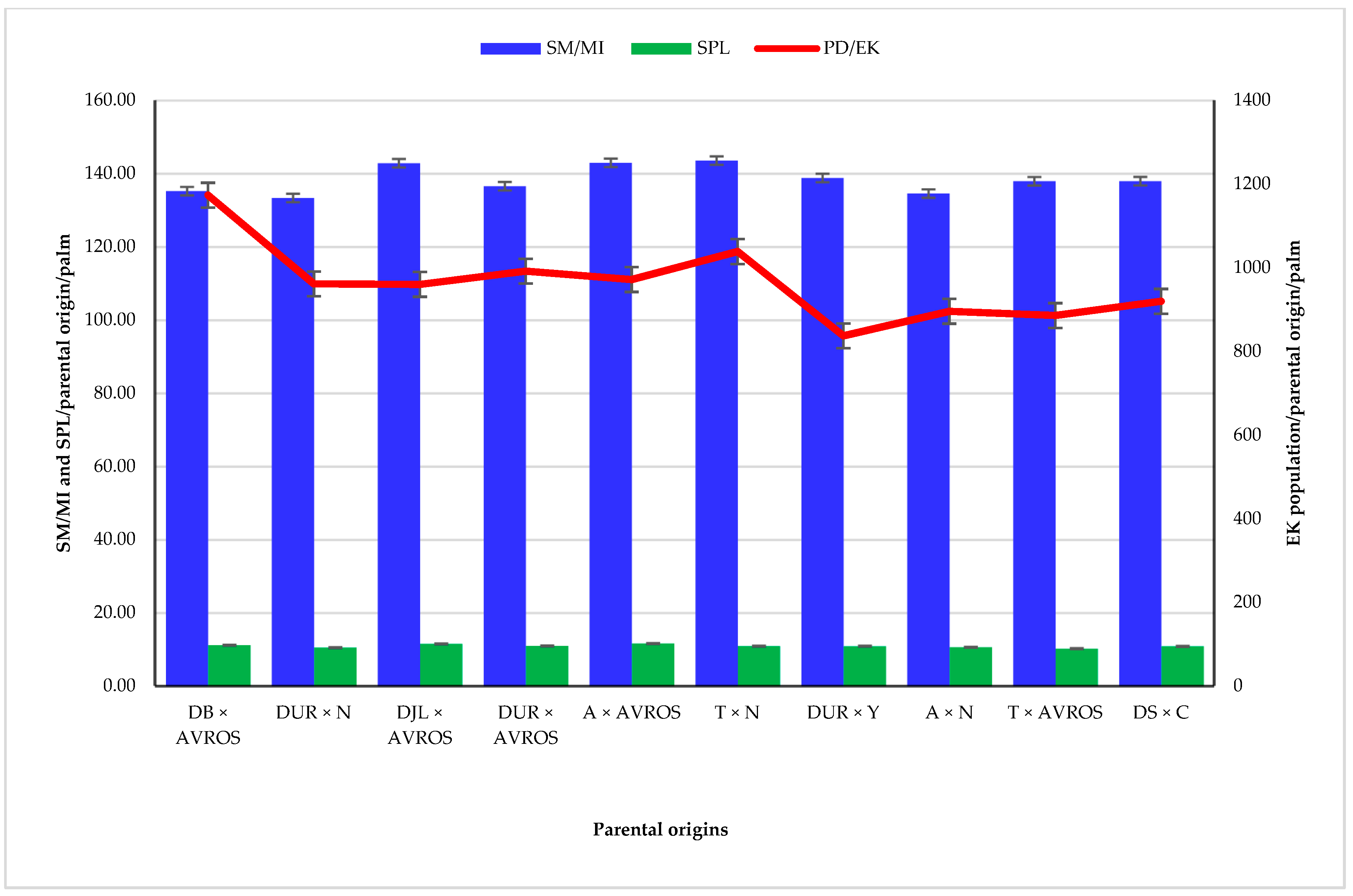
3.3. Mean Number of Spikelets per Male Inflorescence, Spikelet Length, and E. kamerunicus Population Mean among Parental Origins
3.4. Phenotypic Correlation Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Myint, K.A.; Amiruddin, M.D.; Rafii, M.Y.; Samad, M.Y.A.; Ramlee, S.I.; Yaakub, Z.; Oladosu, Y. Genetic diversity and selection criteria of MPOB-Senegal oil palm (Elaeis guineensis Jacq.) germplasm by quantitative traits. Ind. Crops Prod. 2019, 139, 111558. [Google Scholar] [CrossRef]
- Jamian, S.; Norhisham, A.; Ghazali, A.; Zakaria, A.; Azhar, B. Impacts of 2 species of predatory Reduviidae on bagworms in oil palm plantations. Insect Sci. 2017, 24, 285–294. [Google Scholar] [CrossRef] [PubMed]
- Economic Co-operation Development (OECD); Food and Agriculture Organization (FAO). Agricultural Outlook 2017–2026; OECD Publishing: Paris, France, 2017. [Google Scholar] [CrossRef]
- Woittiez, L.S.; van Wijk, M.T.; Slingerland, M.; van Noordwijk, M.; Giller, K.E. Yield gaps in oil palm: A quantitative review of contributing factors. Eur. J. Agron. 2017, 83, 57–77. [Google Scholar] [CrossRef]
- Yusop, M.R.; Sukaimi, J.; Amiruddin, M.D.; Jalloh, M.; Swaray, S.; Yusuff, O.; Chukwu, S.C. Genetic Improvement of Oil Palm Through Recurrent Selection. In The Oil Palm Genome; Springer: Cham, Switzerland, 2020; pp. 35–46. [Google Scholar] [CrossRef]
- Soh, A.C.; Mayes, S.; Roberts, J.A. Oil Palm Breeding: Genetics and Genomics; CRC Press: Boca Raton, FL, USA, 2017; pp. 25–26. [Google Scholar]
- Chong, K.L.; Kanniah, K.D.; Pohl, C.; Tan, K.P. A review of remote sensing applications for oil palm studies. Geo Spat. Inf. Sci. 2017, 20, 184–200. [Google Scholar] [CrossRef] [Green Version]
- Zulkifli, Y.; Norziha, A.; Naqiuddin, M.H.; Fadila, A.M.; Nor Azwani, A.B.; Suzana, M.; Samsul, K.R.; Ong-Abdullah, M.; Singh, R.; Ghulam Kadir, A.P.; et al. Designing the oil palm of the future. J. Oil Palm Res. 2017, 29, 440–455. [Google Scholar]
- Yousefi, M.; Rafie, A.S.M.; Abd Aziz, S.; Azrad, S. Introduction of current pollination techniques and factors affecting pollination effectiveness by Elaeidobius kamerunicus in oil palm plantations on regional and global scale: A review. S. Afr. J. Bot. 2020, 132, 171–179. [Google Scholar] [CrossRef]
- Norman, K.; Ramle, M.; Saharul, A.M.; Mohd, R.S. Fruit set and weevil pollination issues in oil palm. Planter 2018, 94, 565–578. [Google Scholar]
- Haran, J.M.; Beaudoin-Ollivier, L.; Benoit, L.; Kuschel, G. Revision of the palm-pollinating weevil genus Elaeidobius Kuschel, 1952 (Curculionidae, Curculioninae, Derelomini) with descriptions of two new species. Eur. J. Taxon. 2020, 684, 1–32. [Google Scholar] [CrossRef]
- Wahid, M.B.; Masijan, Z.; Hassan, A.H.; Dolmat, M.T. The population census and the pollination efficiency of the weevil Elaeidobius kamerunicus in Malaysia-A status report 1983–1986. In Proceedings of the International Oil Palm/Palm Oil Conferences-Progress and Prospects 1987-Conference 1: 1988. Agriculture, Kuala Lumpur, Malaysia, 23–26 June 1988; IPMKSM: Selangor, Malaysia, 1988. [Google Scholar]
- Dhileepan, K. Variation in populations of the introduced pollinating weevil (Elaeidobius kamerunicus) (Coleoptera: Curculionidae) and its impact on fruit set of oil palm (Elaeis guineensis) in India. Bull. Entomol. Res. 1994, 84, 477–485. [Google Scholar] [CrossRef]
- Hussein, M.Y.; Lajis, N.H.; Ali, J.H. August. Biological and chemical factors associated with the successful introduction of Elaeidobius kamerunicus Faust, the oil palm pollinator in Malaysia. VI Int. Symp. Pollinat. 1990, 288, 81–87. [Google Scholar]
- Syed, R.A.; Law, I.H.; Corley, R.H.V. Insect pollination of oil palm: Introduction, establishment and pollinating efficiency of Elaeidobidus kamerunicus in Malaysia. Planter 1982, 58, 547–561. [Google Scholar]
- Appiah, S.O.; Agyei-Dwarko, D. Studies on Entomophil Pollination towards Sustainable Production and Increased Profitability in the Oil PALM: A Review. Elixir Agric. 2013, 55, 12878–12883. [Google Scholar]
- Frimpong, E.; Adjaloo, M.K. Inadequate Pollinators Blamed for Low Cocoa and Oil Palm Yields. GhanaWeb. 2012. Available online: https://www.ghanaweb.com/GhanaHomePage/business/Inadequate-pollinators-blamed-for-low-cocoa-and-oil-palm-yields-236163 (accessed on 17 May 2019).
- Kouakou, M.; Tuo, Y.; Hala, A.K.; Douan, B.G.; Dagnogo, M.; Koua, H.K. Influence of the number of inflorescences and some climatic factors on the abundance of Elaeidobius kamerunicus (Coleoptera: Curculionidae), main pollinator of the oil palm in Ivory Coast. Int. J. Biol. Chem. Sci. 2018, 12, 1571–1582. [Google Scholar] [CrossRef]
- Muhamad Fahmi, M.H.; Ahmad Bukhary, A.K.; Norma, H.; Idris, A.B. Analysis of volatile organic compound from Elaeis guineensis inflorescences planted on different soil types in Malaysia. In AIP Conference Proceedings; AIP Publishing: Melville, NY, USA, 2016; Volume 1784, p. 060020. [Google Scholar]
- Mohamad, S.A.; Syarif, M.N.Y.; Ahmad, S.N.; Masri, M.M.M.; Hung, K.J.; Kamarudin, N. Population density of elaeidobius kamerunicus faust in different spikelet position at anthesising male inflorescence of elaeis guineensis Jacq in sabah and sarawak, Malaysia. J. Oil Palm. 2020, 1–16. [Google Scholar] [CrossRef]
- O’Brien, C.W.; Woodruff, R.E. First records in the United States and South America of the African oil palm weevils Elaeidobius subvittatus Faust and Elaeidobius kamerunicus Faust (Coleoptera: Curculionidae). Fla. Dep. Agric. Consum. Serv. 1986, 330, 1–2. [Google Scholar]
- Nasir, D.M.; Mamat, N.S.; Muneim, N.A.A.; Ong-Abdullah, M.; Abd Latip, N.F.B.; Su, S.; Hazmi, I.R.B. Morphometric Analysis of the Oil Palm Pollinating Weevil, Elaeidobius kamerunicus (Faust, 1878) (Coleoptera: Curculionidae) from Oil Palm Plantations in Malaysia. J. Entomol. Res. Soc. 2020, 22, 275–291. [Google Scholar]
- Ayuningsih, M. Frekuensi Kunjungan Elaeidobius Kamerunicus Faust Pada Bunga Betina Dan Efektivitasnya Terhadap Pembentukan Buah Kelapa Sawit. PhD. Thesis, Institut Pertanian Bogor, Bogor, Java, Indonesia, 2013. [Google Scholar]
- Meliala, R.A.S. Studi Biologi Serangga Penyerbuk Kelapa Sawit Elaeidobius Kamerunicus Faust (Coleoptera: Curculionidae) Elaeis Guineensis Jacq. di Laboratium. Master’s Thesis, Universitas Sumatera Utara, Medan, Indonesia, 2008. [Google Scholar]
- Tuo, Y.; Koua, H.K.; Hala, N. Biology of Elaeidobius kamerunicus and Elaeidobius plagiatus (Coleoptera: Curculionidae), main pollinators of oil palm in West Africa. Eur. J. Sci. Res. 2011, 49, 426–432. [Google Scholar]
- Soetopo, D. Population of oil palm pollinator insect (Elaeidobius kamerunicus faust.) at PTP Nusantara VIII Cisalak Baru, Rangkasbitung-Banten. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2020; Volume 418, p. 012045. [Google Scholar]
- Syed, R.A. Los insectos polinizadores de la palma africana. Rev. Palmas 1984, 5, 19–64. [Google Scholar]
- Corley, R.H.V.; Tinker, P.B. The Oil Palm, 5th ed.; John Wiley AND Sons: West Sussex, UK, 2015; pp. 37–42. [Google Scholar]
- Prasetyo, A.E.; Purba, W.O.; Susanto, A. Elaeidobius kamerunicus: Application of hatch and carry technique for increasing oil palm fruit set. J. Oil Palm Res. 2014, 26, 195–202. [Google Scholar]
- Basri, M.W.; Norman, K.H. Role and effectiveness of Elaeidobius kamerunicus, Thrips hawaiiensis and Pyroderces sp. in pollination of mature oil palm in Peninsular Malaysia. Elaeis 1997, 9, 1–16. [Google Scholar]
- Kamaruddin, N.; Moslim, R.; Mohamad, S.A.; Sulaiman, M.R. Environmental effects on insects and their population dynamic. J. Entomol. Zool. Stud. 2018, 2, 1–7. [Google Scholar]
- Sisye, S.E. Diversity and Abundance of Oil Palm Insect Pollinators in Selected Agro-Ecological Zones in Uganda. Bachelor’s Thesis, (Biological) of Makerere University, Kampala, Uganda, 2018; pp. 11–21. [Google Scholar]
- Swaray, S.; Din Amiruddin, M.; Rafii, M.Y.; Jamian, S.; Ismail, M.F.; Jalloh, M.; Marjuni, M.; Mustakim, M.M.; Yusuff, O. Influence of Parental dura and pisifera genetic origins on oil palm fruit set ratio and yield components in their D×P progenies. Agronomy 2020, 10, 1793. [Google Scholar] [CrossRef]
- De Almeida Rios, S.; da Cunha, R.N.V.; Lopes, R.; Barcelos, E.; da Rocha, R.N.C.; de Lima, W.A.A. Correlation and Path analysis for yield components in Dura oil palm germplasm. Ind. Crop. Prod. 2018, 112, 724–733. [Google Scholar] [CrossRef]
- Chiu, S.B.; Khoo, K.C.; Hussein, M.Y. A method of estimating the natural population of the pollinating weevil Elaeidobius kamerunicus Faust, of oil palm [Elaeis guineensis]. In Proceedings of the 1. Regional Symposium on Biological Control, Serdang, Selangor, Malaysia, 4–6 September 1985; Penerbit Universiti Pertanian Malaysia: Selangor, Malaysia, 1986. [Google Scholar]
- Janick, J.; Paull, R.E. The Encyclopedia of Fruit and Nuts; CABI Publishing: Oxfordshire, UK, 2008; p. 954. [Google Scholar]
- Abdullah, N.; Yusop, M.R.; Ithnin, M.; Saleh, G.; Latif, M.A. Genetic variability of oil palm parental genotypes and performance of its’ progenies as revealed by molecular markers and quantitative traits. Comptes Rendus Biol. 2011, 334, 290–299. [Google Scholar] [CrossRef]
- Barcelos, E.; Rios, S.D.A.; Cunha, R.N.; Lopes, R.; Motoike, S.Y.; Babiychuk, E.; Skirycz, A.; Kushnir, S. Oil palm natural diversity and the potential for yield improvement. Front. Plant Sci. 2015, 6, 190. [Google Scholar] [CrossRef]
- Abd Latip, N.F.B.; Abidin, C.M.R.B.Z.; Abd Ghani, I.B.; Fahmi, M.; Al–Talafha, H. Effect of oil palm planting materials, rainfall, number of male inflorescence and spikelet on the population abundance of oil palm pollinator, Elaeidobius kamerunicus Faust (Coleoptera: Curculionidae). Serangga 2018, 23, 35–45. [Google Scholar]
- Setyamidjaja, D. Kelapa Sawit Teknik Budaya, Panen, Dan Pengelolaan; Kanisius: Jakarta, Indonesia, 2006; 127p. [Google Scholar]
- Yue, J.; Yan, Z.; Bai, C.; Chen, Z.; Lin, W.; Jiao, F. Pollination activity of Elaeidobius kamerunicus (Coleoptera: Curculionoidea) on oil palm on Hainan island. Fla. Entomol. 2015, 98, 499–505. [Google Scholar] [CrossRef]
- Fatihah, N.A.; Fahmi, M.M.; Luqman, H.A.; Nadiah, S.S.; Teo, T.M.; Riza, I.H.; Idris, A.B. Effects of rainfall, number of male inflorescences and spikelets on the population abundance of Elaeidobius Kamerunicus (Coleoptera: Curculionidae). Sains Malays. 2019, 48, 15–21. [Google Scholar]
- Hoedemaekers, K.; Derksen, J.; Hoogstrate, S.W.; Wolters-Arts, M.; Oh, S.A.; Twell, D.; Mariani, C.; Rieu, I. Bursting pollen is required to organize the pollen germination plaque and pollen tube tip in Arabidopsis thaliana. New Phytol. 2015, 206, 255–267. [Google Scholar] [CrossRef] [Green Version]
- Vianna, M.R.; Junior, P.D.M.; de Oliveira, L.A. Manejo de Polinizadores e o incremento da produtividade agrícola: Uma abordagem sustentável dos serviços do ecossistema. Cad. Agroecol. 2007, 2, 44–447. [Google Scholar]
- Moura, J.I.L.; Cividanes, F.J.; Pires, J.L.; Santos, L.P.; Santos, E.A.; Valle, R.R.; Delabie, J.H.C. Behavior of Curculionidae pollinators on oil palm inflorescences in the State of Bahia, Brazil. Agrotrópica 2010, 22, 45–50. [Google Scholar]
- Harun, M.H.; Noor, M.R.M. Fruit set and oil palm bunch components. J. Oil Palm Res. 2002, 14, 24–33. [Google Scholar]
- Luqman, H.A.; Noor Nasuha, A.A.; Dzulhelmi, M.N.; Nurul Fatihah, A.L.; Muhamad Fahmi, M.H.; Teo, T.M.; Idris, A.B.; Izfa Riza, H. Diversity and composition of beetles (Order: Coleoptera) in three different ages of oil palms in Lekir oil palm plantation, Perak, Malaysia. Serangga 2018, 23, 58–71. [Google Scholar]
- Rizuan, Z.A.; Hisham, N.H.; Shamsudin, A. Role of pollinating weevil (Elaeidobius kamerunicus), seasonal effect and its relation to fruit set in oil palm area of FELDA. In Proceedings of the PIPOC 2013 International Palm Oil Congress, Kuala Lumpur Malaysia, 19–21 November 2013; pp. 19–21. [Google Scholar]
- Amit, B.; Tuen, A.A.; Haron, K.; Harun, M.H.; Kamarudin, N. The diet of Yellow-vented Bulbul (Pycnonotus goiavier) in oil palm agroecosystems. J. Oil Palm Res. 2015, 27, 417–424. [Google Scholar]
- HA, M.L.; Dzulhelmi, M.N. The potential natural predators of Elaeidobius kamerunicus Faust, 1878 (Coleoptera: Curculionidae) in Malaysia. Serangga 2018, 22, 240–252. [Google Scholar]
- Sugih, W.; Setyobudi, H.; Achmad, F.; Thiagarajan. Influence of rainfall, palm age and assisted pollination on oil palm fruit set in Riau, Indonesia. In Proceedings of the International Palm Oil Congress: Competitiveness for the 21st Century, Agriculture Conference, Kuala Lumpur, Malaysia, 23–28 September 1996; IPMKSM: Selangor, Malaysia, 1996; pp. 207–220. [Google Scholar]
- MDIC. Malaysia Department of Information on Climate. 2016. Available online: https://www.malaysia.gov.my/portal/content/144 (accessed on 31 December 2020).
- Yusdayati, R.R.; Hamid, N.H. Effect of Several Insecticide against Oil Palm Pollinator’s Weevil, Elaeidobius Kamerunicus (Coleoptera: Curculionidae). Serangga 2015, 20, 27–35. [Google Scholar]
- Arolu, I.W.; Rafii, M.Y.; Marjuni, M.; Hanafi, M.M.; Sulaiman, Z.; Rahim, H.A.; Abidin, M.I.Z.; Amiruddin, M.D.; Din, A.K.; Nookiah, R. Breeding of high yielding and dwarf oil palm planting materials using Deli dura × Nigerian pisifera population. Euphytica 2017, 213, 154. [Google Scholar] [CrossRef]
- Meléndez, M.R.; Ponce, W.P. Pollination in the oil palms Elaeis guineensis, E. oleifera and their hybrids (O×G), in tropical America. Pesq. Agropec. Trop. 2016, 46, 102–110. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S/N | Crossing Materials | Pedigree No. ♂ Palm × ♀ Palm | Code |
|---|---|---|---|
| 1 | AVROS × Deli Banting | 0.394/456 × 0.279/24 | ECPHP415 |
| 2 | Nigeria × Deli Ulu Remis | 0.337/552 × 0.338/361 | ECPHP500 |
| 3 | AVROS × Deli Banting | 0.394/234 × 0.279/24 | ECPHP550 |
| 4 | AVROS × Deli Johor Labis | 0.394/234 × 0.281/44 | ECPHP618 |
| 5 | AVROS × Deli Ulu Remis | 0.174/480 × 0.254/191 | PK4118 |
| 6 | AVROS × Angola | 0.174/480 × 0.311/405 | PK4465 |
| 7 | Nigeria × Tanzania | 0.337/1092 × 0.256/2058 | PK4474 |
| 8 | AVROS × Angola | 0.394/24 × 0.311/405 | PK4482 |
| 9 | AVROS × Angola | 0.174/247 × 0.312/99 | PK4504 |
| 10 | Avros × Angola | 0.174/211 × 0.311/269 | PK4505 |
| 11 | Yangambi × Deli Ulu Remis | 0.395/204 × 0.332/451 | PK4529 |
| 12 | AVROS × Deli Ulu Remis | 0.394/24 × 0.332/100 | PK4535 |
| 13 | Nigeria, × Angola | 0.337/1092 × 0.312/682 | PK4539 |
| 14 | Nigeria × Deli Ulu Remis | 0.337/1092 × 0.332/218 | PK4540 |
| 15 | Yangambi × Deli Ulu Remis | 0.395/204 × 0.332/45 | PK4548 |
| 16 | AVROS × Deli Ulu Remis | 0.395/419 × 0.332/278 | PK4550 |
| 17 | AVROS × Tanzania | 0.394/24 × 0.256/2313 | PK4570 |
| 18 | AVROS × Deli Ulu Remis | 0.395/419 × 0.332/340 | PK4591 |
| 19 | Nigeria × Deli Ulu Remis | 0.337/554 × 0.332/220 | PK4621 |
| 20 | Nigeria × Deli Ulu Remis | 0.337/1091 × 0.332/116 | PK4648 |
| 21 | Nigeria × Tanzania | 0.337/1092 × 0.256/2425 | PK4651 |
| 22 | AVROS × Deli Ulu Remis | 0.395/372 × 0.332/116 | PK4674 |
| 23 | Nigeria × Angola | 0.337/291 × 0.312/1241 | PK4679 |
| 24 | Cameroon × Deli Serdang | 0.219/1371 × 0.212/6 | PK4841 |
| S/V | DF | MD1 | FD1 | EKD1 | MD2 | FD2 | EKD2 | MD3 | FD3 | EKD3 |
|---|---|---|---|---|---|---|---|---|---|---|
| Replications (R) | 3 | ˂0.01 ns | 0.01 ns | 0.01 ns | 0.02 ns | 0.10 ns | 0.11 ns | 0.13 ns | 2.16 ns | 1.98 ns |
| Hybrids (G) | 23 | 0.01 ns | 0.08 ** | 0.10 ** | 0.11 ** | 1.29 ** | 1.87 ** | 0.38 ** | 5.92 ** | 8.87 ** |
| Error (e) | 60 | ˂0.01 | 0.01 | 0.02 | 0.02 | 12.17 | 0.28 | 4.33 | 0.76 | 1.10 |
| Variance component | ||||||||||
| σ2g | ˂0.01 (13.33) + | ˂0.01 (55.24) | 0.03 (60.00) | 0.02 (50.00) | 0.28 (58.33) | 0.42 (60.87) | 0.08 (53.33) | 1.33 (63.64) | 2.03 (65.48) | |
| σ2e | ˂0.01 (86.67) | ˂0.01 (44.76) | 0.02 (40.00) | 0.02 (50.00) | 0.2 (41.66) | 0.27 (39.13) | 0.07 (46.67) | 0.76 (36.36) | 1.07 (34.52) | |
| σ2ph | ˂0.01 | ˂0.01 | 0.05 | 0.05 | 0.48 | 0.69 | 0.15 | 2.09 | 3.10 | |
| Mean | 0.22 | 1.08 | 1.3 | 1.73 | 6.27 | 7.99 | 3.42 | 12.94 | 16.36 | |
| Std Error | 0.01 | 0.02 | 0.02 | 0.02 | 0.07 | 0.09 | 0.04 | 0.16 | 0.19 | |
| S/V | DF | MD4 | FD4 | EKD4 | MD5 | FD5 | EKD5 | MD6 | FD6 | EKD6 |
| Replications (R) | 3 | 0.10 ns | 1.57 * | 1.75 * | 0.01 ns | 0.18 ns | 0.22 ns | ˂0.01 ns | ˂0.01 ns | ˂0.01 ns |
| Hybrids (G) | 23 | 0.13 ** | 1.48 ** | 2.27 ** | 0.04 ns | 0.31 ** | 0.52 ** | ˂0.01 ns | 0.01 ** | 0.01 ** |
| Error (e) | 60 | 0.05 | 0.37 | 0.59 | 0.03 | 0.10 | 0.17 | ˂0.01 | ˂0.01 | ˂0.01 |
| Variance component | ||||||||||
| σ2g | 0.02 (28.57) | 0.30 (44.78) | 0.46 (43.81) | ˂0.01 (11.77) | 0.06 (35.29) | 0.10 (37.04) | ˂0.01 (7.50) | ˂0.01 (40.00) | ˂0.01 (47.50) | |
| σ2e | 0.05 (71.43) | 0.37 (55.22) | 0.59 (56.19) | 0.03 (88.23) | 0.11 (64.71) | 0.17 (62.96) | ˂0.01 (92.50) | ˂0.01 (60.00) | ˂0.01 (52.50) | |
| σ2ph | 0.07 | 0.67 | 1.05 | 0.034 | 0.17 | 0.27 | ˂0.01 | ˂0.01 | ˂0.01 | |
| Mean | 2.14 | 7.02 | 9.16 | 1.02 | 3.13 | 4.15 | 0.02 | 0.07 | 0.09 | |
| Stderr | 0.03 | 0.09 | 0.11 | 0.02 | 0.04 | 0.06 | ˂0.01 | 0.01 | 0.01 |
| S/V | DF | MEK/S | PD/EK |
|---|---|---|---|
| Replications (R) | 3 | 3.41 ns | 203,442.10 ns |
| Months (M) | 11 | 684.25 ** | 18,934,909.40 ** |
| Hybrids (G) | 23 | 27.07 ** | 863,523.40 ** |
| M*G | 253 | 3.63 ** | 140,595.90 * |
| Error (e) | 753 | 1.91 | 117,313.40 |
| Mean | 8.55 | 973.68 | |
| Stderr | 0.10 | 18.31 | |
| No. | Month | ||
| 1 | February 2019 | 10.14 c ± 0.15 | 1158.57 d ± 41.28 |
| 2 | March 2019 | 11.41 b ± 0.27 | 1526.78 b ± 69.00 |
| 3 | April 2019 | 7.57 d ± 0.14 | 709.34 ef ± 24.45 |
| 4 | May 2019 | 5.92 e ± 0.22 | 548.02 f ± 31.91 |
| 5 | June 2019 | 7.70 d ± 0.17 | 1216.00 cd ± 46.79 |
| 6 | July 2019 | 8.22 d ± 0.17 | 653.38 ef ± 25.38 |
| 7 | August 2019 | 3.96 f ± 0.12 | 360.63 g ± 14.24 |
| 8 | September 2019 | 4.36 f ± 0.17 | 359.57 g ± 16.71 |
| 9 | October 2019 | 10.62 c ± 0.16 | 1218.66 cd ±34.92 |
| 10 | November 2019 | 8.10 d ±0.16 | 729.16 e ± 23.24 |
| 11 | December 2019 | 11.79 b ±0.18 | 1357.58 bc ± 49.55 |
| 12 | January 2020 | 12.81 a ± 0.23 | 1846.49 a ± 60.69 |
| Hybrid | |||
| 1 | ECPHP415 | 8.91 b–e ± 0.13 | 1104.63 a–e ± 99.34 |
| 2 | ECPHP500 | 9.80 ab ± 0.26 | 1226.82 ab ± 61.21 |
| 3 | ECPHP550 | 10.25 a ± 0.11 | 1241.39 a ± 73.74 |
| 4 | ECPHP618 | 8.74 b–e ± 0.32 | 959.91 b–f ± 72.82 |
| 5 | PK4118 | 9.01 b–e ± 0.22 | 1149.72 a–c ± 62.19 |
| 6 | PK4465 | 7.99 d–f ± 0.05 | 975.24 a–f ± 40.75 |
| 7 | PK4474 | 8.49 c–f ± 0.2 | 1053.40 a–f ± 6.06 |
| 8 | PK4482 | 8.06 d–f ± 0.21 | 941.35 c–f ± 74.68 |
| 9 | PK4504 | 8.67 c–e ± 0.5 | 980.68 a–f ± 60.58 |
| 10 | PK4505 | 8.77 b–e ± 0.19 | 988.46 a–f ± 27.02 |
| 11 | PK4529 | 7.94 d–f ± 0.38 | 840.10 d–g ± 37.38 |
| 12 | PK4535 | 8.47 c–f ± 0.19 | 899.19 c–g ± 49.06 |
| 13 | PK4539 | 7.42 fg ± 0.14 | 813.47 fg ± 24.54 |
| 14 | PK4540 | 7.91 ef ± 0.15 | 880.07 c–g ± 19.5 |
| 15 | PK4548 | 8.25 d–f ± 0.39 | 833.60 e–g ± 22.37 |
| 16 | PK4550 | 8.38 c–f ± 0.21 | 986.65 a–f ± 58.97 |
| 17 | PK4570 | 8.33 c–f ± 0.16 | 885.42 c–g ± 30.67 |
| 18 | PK4591 | 8.51 c–f ± 0.23 | 928.37 c–f ± 39.54 |
| 19 | PK4621 | 6.49 g ± 0.33 | 629.90 g ± 44.09 |
| 20 | PK4648 | 9.43 a–c ± 0.2 | 1108.05 a–d ± 94.8 |
| 21 | PK4651 | 8.96 b–e ± 0.26 | 1023.26 a–f ± 46.12 |
| 22 | PK4674 | 8.33 c–f ± 0.41 | 992.81 a–f ± 47.68 |
| 23 | PK4679 | 9.04 b–d ± 0.28 | 977.63 a–f ± 28.8 |
| 24 | PK4841 | 8.53 c–e ± 0.27 | 919.31 c–f ± 66.73 |
| Mean ± Stderr | 8.55 ± 0.10 | 973.68 ± 18.31 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Swaray, S.; Y. Rafii, M.; Din Amiruddin, M.; Firdaus Ismail, M.; Jamian, S.; Jalloh, M.; Oladosu, Y.; Mustakim Mohamad, M.; Marjuni, M.; Kolapo, O.K.; et al. Assessment of Oil Palm Pollinating Weevil (Elaeidobius kamerunicus) Population Density in Biparental dura × pisifera Hybrids on Deep Peat-Soil in Perak State, Malaysia. Insects 2021, 12, 221. https://doi.org/10.3390/insects12030221
Swaray S, Y. Rafii M, Din Amiruddin M, Firdaus Ismail M, Jamian S, Jalloh M, Oladosu Y, Mustakim Mohamad M, Marjuni M, Kolapo OK, et al. Assessment of Oil Palm Pollinating Weevil (Elaeidobius kamerunicus) Population Density in Biparental dura × pisifera Hybrids on Deep Peat-Soil in Perak State, Malaysia. Insects. 2021; 12(3):221. https://doi.org/10.3390/insects12030221
Chicago/Turabian StyleSwaray, Senesie, Mohd Y. Rafii, Mohd Din Amiruddin, Mohd Firdaus Ismail, Syari Jamian, Momodu Jalloh, Yusuff Oladosu, Mohd Mustakim Mohamad, Marhalil Marjuni, Olalekan Kazeem Kolapo, and et al. 2021. "Assessment of Oil Palm Pollinating Weevil (Elaeidobius kamerunicus) Population Density in Biparental dura × pisifera Hybrids on Deep Peat-Soil in Perak State, Malaysia" Insects 12, no. 3: 221. https://doi.org/10.3390/insects12030221
APA StyleSwaray, S., Y. Rafii, M., Din Amiruddin, M., Firdaus Ismail, M., Jamian, S., Jalloh, M., Oladosu, Y., Mustakim Mohamad, M., Marjuni, M., Kolapo, O. K., & Chukwu, S. C. (2021). Assessment of Oil Palm Pollinating Weevil (Elaeidobius kamerunicus) Population Density in Biparental dura × pisifera Hybrids on Deep Peat-Soil in Perak State, Malaysia. Insects, 12(3), 221. https://doi.org/10.3390/insects12030221