
Organic Farming and Cover-Crop Management Reduce Pest Predation in Austrian Vineyards

 ,
,  , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Predation Rate Assessment
2.3. Landscape Analysis
2.4. Data Analysis
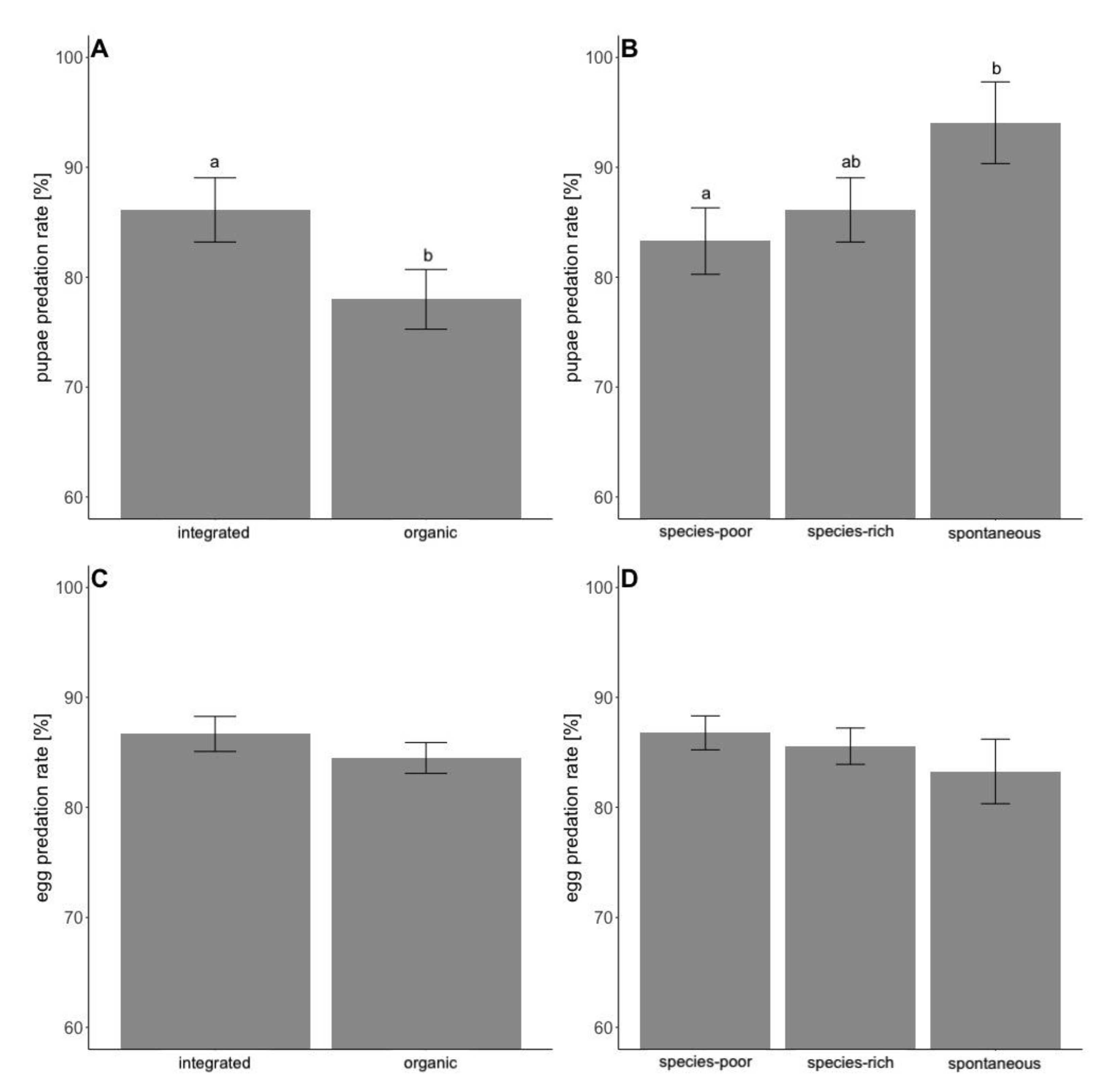
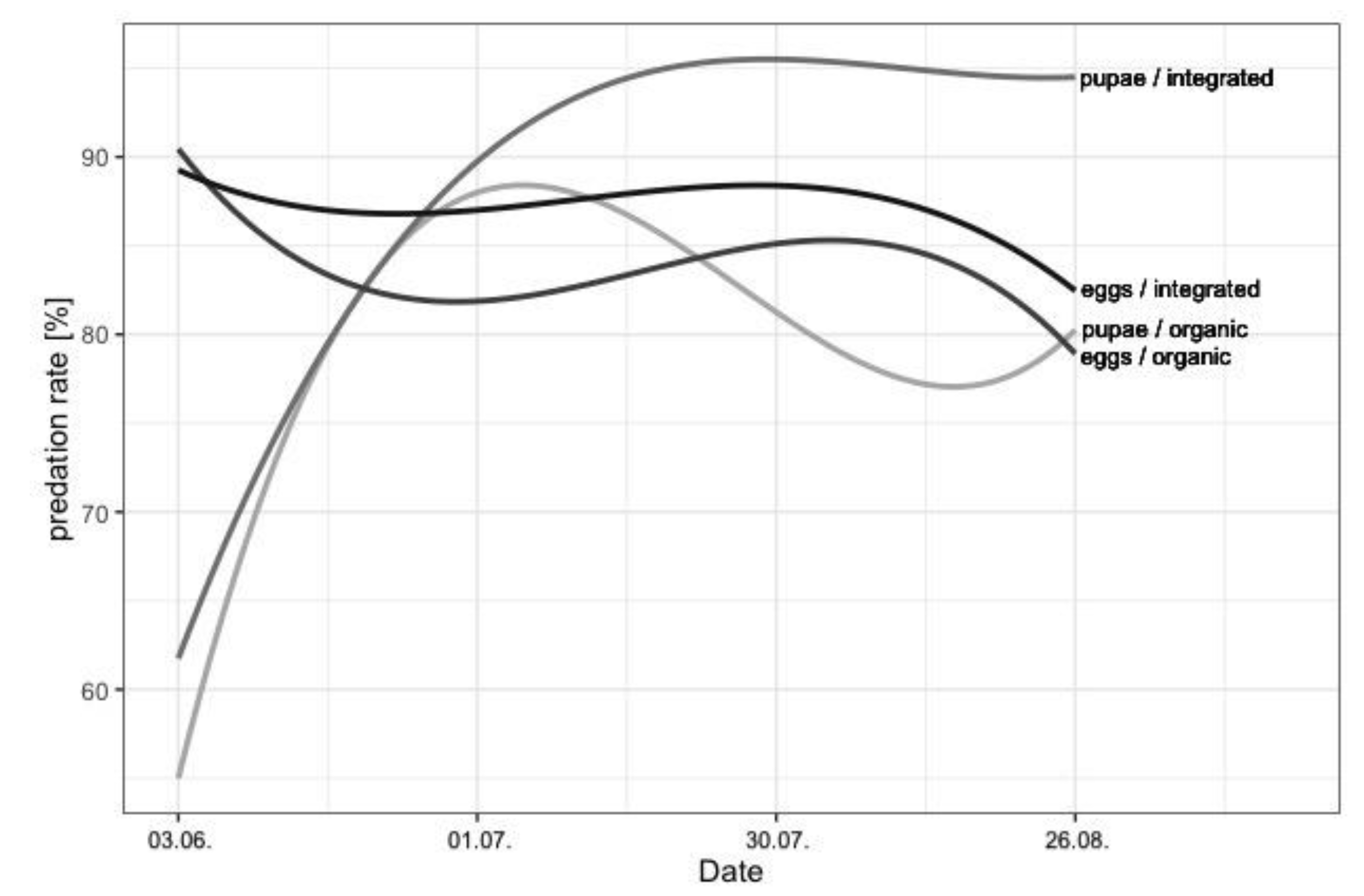
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hallmann, C.A.; Sorg, M.; Jongejans, E.; Siepel, H.; Hofland, N.; Schwan, H.; Stenmans, W.; Müller, A.; Sumser, H.; Hörren, T.; et al. More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS ONE 2017, 12, e0185809. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Seibold, S.; Gossner, M.M.; Simons, N.K.; Blüthgen, N.; Müller, J.; Ambarlı, D.; Ammer, C.; Bauhus, J.; Fischer, M.; Habel, J.C.; et al. Arthropod decline in grasslands and forests is associated with landscape-level drivers. Nature 2019, 574, 671–674. [Google Scholar] [CrossRef]
- Bianchi, F.J.J.A.; Booij, C.J.H.; Tscharntke, T. Sustainable pest regulation in agricultural landscapes: A review on landscape composition, biodiversity and natural pest control. Proc. R. Soc. B Biol. Sci. 2006, 273, 1715–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurr, G.M.; Wratten, S.D.; Luna, J.M. Multi-function agricultural biodiversity: Pest management and other benefits. Basic Appl. Ecol. 2003, 4, 107–116. [Google Scholar] [CrossRef]
- Bengtsson, J.; Ahnström, J.; Weibull, A.-C. The effects of organic agriculture on biodiversity and abundance: A meta-analysis: Organic agriculture, biodiversity and abundance. J. Appl. Ecol. 2005, 42, 261–269. [Google Scholar] [CrossRef]
- Puig-Montserrat, X.; Stefanescu, C.; Torre, I.; Palet, J.; Fabregas, E.; Dantart, J.; Arrizabalaga, A.; Flaquer, C. Effects of organic and conventional crop management on vineyard biodiversity. AEE 2017, 243, 19–26. [Google Scholar] [CrossRef]
- Muneret, L.; Mitchell, M.; Seufert, V.; Aviron, S.; Djoudi, E.A.; Pétillon, J.; Plantegenest, M.; Thiéry, D.; Rusch, A. Evidence that organic farming promotes pest control. Nat. Sustain. 2018, 1, 361. [Google Scholar] [CrossRef]
- Bruggisser, O.T.; Schmidt-Entling, M.H.; Bacher, S. Effects of vineyard management on biodiversity at three trophic levels. Biol. Conserv. 2010, 143, 1521–1528. [Google Scholar] [CrossRef] [Green Version]
- Reiff, J.M.; Ehringer, M.; Hoffmann, C.; Entling, M.H. Fungicide reduction favors the control of phytophagous mites under both organic and conventional viticulture. AEE 2021, 305, 107172. [Google Scholar] [CrossRef]
- Muneret, L.; Auriol, A.; Thiéry, D.; Rusch, A. Organic farming at local and landscape scales fosters biological pest control in vineyards. Ecol. Appl. 2018, 29, e01818. [Google Scholar] [CrossRef] [Green Version]
- Veres, A.; Petit, S.; Conord, C.; Lavigne, C. Does landscape composition affect pest abundance and their control by natural enemies? A review. AEE 2013, 166, 110–117. [Google Scholar] [CrossRef]
- Paredes, D.; Rosenheim, J.A.; Chaplin-Kramer, R.; Winter, S.; Karp, D.S. Landscape simplification increases vineyard pest outbreaks and insecticide use. Ecol. Lett. 2021, 24, 73–83. [Google Scholar] [CrossRef]
- Serée, L.; Rouzes, R.; Thiéry, D.; Rusch, A. Temporal variation of the effects of landscape composition on lacewings (Chrysopidae: Neuroptera) in vineyards. Agric. For. Entomol. 2020, 22, 274–283. [Google Scholar] [CrossRef]
- Thomson, L.J.; Hoffmann, A.A. Vegetation increases the abundance of natural enemies in vineyards. Biol. Control. 2009, 49, 259–269. [Google Scholar] [CrossRef]
- Gonçalves, F.; Carlos, C.; Aranha, J.; Torres, L. Does habitat heterogeneity affect the diversity of epigaeic arthropods in vineyards? Agric. For. Entomol. 2018, 20, 366–379. [Google Scholar] [CrossRef]
- Judt, C.; Guzmán, G.; Gómez, J.A.; Cabezas, J.M.; Entrenas, J.A.; Winter, S.; Zaller, J.G.; Paredes, D. Diverging Effects of Landscape Factors and Inter-Row Management on the Abundance of Beneficial and Herbivorous Arthropods in Andalusian Vineyards (Spain). Insects 2019, 10, 320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karp, D.S.; Chaplin-Kramer, R.; Meehan, T.D.; Martin, E.A.; DeClerck, F.; Grab, H.; Gratton, C.; Hunt, L.; Larsen, A.E.; Martínez-Salinas, A.; et al. Crop pests and predators exhibit inconsistent responses to surrounding landscape composition. Proc. Natl. Acad. Sci. USA 2018, 115, e7863–e7870. [Google Scholar] [CrossRef] [Green Version]
- Muneret, L.; Thiéry, D.; Joubard, B.; Rusch, A. Deployment of organic farming at a landscape scale maintains low pest infestation and high crop productivity levels in vineyards. J. Appl. Ecol. 2018, 55, 1516–1525. [Google Scholar] [CrossRef]
- Wilson, H.; Miles, A.F.; Daane, K.M.; Altieri, M.A. Landscape diversity and crop vigor outweigh influence of local diversification on biological control of a vineyard pest. Ecosphere 2017, 8, e01736. [Google Scholar] [CrossRef]
- Tscharntke, T.; Karp, D.S.; Chaplin-Kramer, R.; Batáry, P.; DeClerck, F.; Gratton, C.; Hunt, L.; Ives, A.; Jonsson, M.; Larsen, A.; et al. When natural habitat fails to enhance biological pest control—Five hypotheses. Biol. Conserv. 2016, 204, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Rusch, A.; Binet, D.; Delbac, L.; Thiéry, D. Local and landscape effects of agricultural intensification on Carabid community structure and weed seed predation in a perennial cropping system. Landsc. Ecol. 2016, 31, 2163–2174. [Google Scholar] [CrossRef]
- Burgio, G.; Marchesini, E.; Reggiani, N.; Montepaone, G.; Schiatti, P.; Sommaggio, D. Habitat managementof organic vineyard in Northern Italy: The role of cover plants management on arhtropod functional biodiversity. Bull. Entomol. Res. 2016, 106, 759–768. [Google Scholar] [CrossRef]
- Sáenz-Romo, M.G.; Veas-Bernal, A.; Martínez-García, H.; Ibáñez-Pascual, S.; Martínez-Villar, E.; Campos-Herrera, R.; Marco-Mancebón, V.S.; Pérez-Moreno, I. Effects of Ground Cover Management on Insect Predators and Pests in a Mediterranean Vineyard. Insects 2019, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, M.; Kleijn, D.; Williams, N.M.; Tschumi, M.; Blaauw, B.R.; Bommarco, R.; Campbell, A.J.; Dainese, M.; Drummond, F.A.; Entling, M.H.; et al. The effectiveness of flower strips and hedgerows on pest control, pollination services and crop yield: A quantitative synthesis. Ecol. Lett. 2020. [CrossRef]
- Eckert, M.; Mathulwe, L.L.; Gaigher, R.; Joubert van der Merwe, L.; Pryke, J.S. Native cover crops enhance arthropod diversity in vineyards of the Cape Floristic Region. J. Insect Conserv. 2020, 24, 133–149. [Google Scholar] [CrossRef]
- Thiéry, D.; Moreau, J. Relative performance of European grapevine moth (Lobesia botrana) on grapes and other hosts. Oecologia 2005, 143, 548–557. [Google Scholar] [CrossRef]
- Svobodová, E.; Trnka, M.; Žalud, Z.; Semerádová, D.; Dubrovský, M.; Eitzinger, J.; Štěpánek, P.; Brázdil, R. Climate variability and potential distribution of selected pest species in south Moravia and north-east Austria in the past 200 years—Lessons for the future. J. Agric. Sci. 2014, 152, 225–237. [Google Scholar] [CrossRef]
- Pennington, T.; Reiff, J.M.; Theiss, K.; Entling, M.H.; Hoffmann, C. Reduced fungicide applications improve insect pest control in grapevine. BioControl 2018, 63, 687–695. [Google Scholar] [CrossRef]
- Marchesini, E.; Monta, L.D. Observations on natural enemies of Lobesia botrana (Den. & Schiff.)(Lepidoptera, Tortricidae) in Venetian vineyards. Boll. di Zool Agrar e di Bachic 1994, 26, 201–230. [Google Scholar]
- Papura, D.; Roux, P.; Joubard, B.; Razafimbola, L.; Fabreguettes, O.; Delbac, L.; Rusch, A. Predation of grape berry moths by harvestmen depends on landscape composition. Biol. Control. 2020, 150, 104358. [Google Scholar] [CrossRef]
- Bagnoli, B.; Lucchi, A. Parasitoids of Lobesia botrana (Den. & Schiff.) in Tuscany. IOBC/WPRS Bull. 2006, 29, 139–142. [Google Scholar]
- Hoffmann, C.; Köckerling, J.; Biancu, S.; Gramm, T.; Michl, G.; Entling, M.H. Can Flowering Greencover Crops Promote Biological Control in German Vineyards? Insects 2017, 8, 121. [Google Scholar] [CrossRef] [Green Version]
- Xuéreb, A.; Thiéry, D. Does natural larval parasitism of Lobesia botrana (Lepidoptera: Tortricidae) vary between years, generation, density of the host and vine cultivar? Bull. Entomol. Res. 2006, 96, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Thomson, L.J.; Glenn, D.C.; Hoffmann, A.A. Effects of sulfur on Trichogramma egg parasitoids in vineyards: Measuring toxic effects and establishing release windows. Aust. J. Exp. Agric. 2000, 40, 1165–1171. [Google Scholar] [CrossRef]
- Thomson, L.J.; Hoffmann, A.A. Field validation of laboratory-derived IOBC toxicity ratings for natural enemies in commercial vineyards. Biol. Control. 2006, 39, 507–515. [Google Scholar] [CrossRef]
- G#xFC;ven, B.; Göven, M.A. Side effects of pesticides used in cotton and vineyard areas of Aegean Region on the green lacewing, Chrysoperla carnea (Steph.) (Neuroptera: Chrysopidae), in the laboratory. IOBC/WPRS Bull. 2003, 26, 21–24. [Google Scholar]
- Nash, M.A.; Hoffmann, A.A.; Thomson, L.J. Identifying signature of chemical applications on indigenous and invasive nontarget arthropod communities in vineyards. Ecol. Appl. 2010, 20, 1693–1703. [Google Scholar] [CrossRef]
- Battaglin, W.A.; Sandstrom, M.W.; Kuivila, K.M.; Kolpin, D.W.; Meyer, M.T. Occurrence of azoxystrobin, propiconazole, and selected other fungicides in US streams, 2005–2006. Water Air Soil Pollut. 2011, 218, 307–322. [Google Scholar] [CrossRef]
- Viret, O.; Siegfried, W.; Dubuis, P.H.; Gindro, K. Falscher Rebenmehltau; Eidgenössisches Departement für Wirtschaft, Bildung und Forschung WBF, Agroscope, AMTRA: Lausanne, Switzerland, 2014; pp. 1–2. [Google Scholar]
- Verzeichnis Zugelassener Pflanzenschutzmittel—Weinbau. Deutsches Pflanzenschutzmittelregister. Available online: https://apps2.bvl.bund.de/psm/jsp/ListeMain.jsp?page=1&ts=1610276481000 (accessed on 10 January 2021).
- Rubel, F.; Brugger, K.; Haslinger, K.; Auer, I. The climate of the European Alps: Shift of very high resolution Köppen-Geiger climate zones 1800–2100. Meteorol. Z. 2017, 26, 115–125. [Google Scholar] [CrossRef]
- Wetterstatistik. BOKU Institut für Pflanzenschutz. Available online: http://rebschutz.boku.ac.at (accessed on 20 March 2020).
- Holmes, C.W. The Massey grass meter. Dairy Farming Annu. 1974, 26–30. [Google Scholar]
- Möth, S.; Walzer, A.; Redl, M.; Petrović, B.; Hoffmann, C.; Winter, S. Unexpected effects of local management and landscape composition on predatory mites and their food resources in vineyards. Insects 2021, 12, 180. [Google Scholar] [CrossRef]
- Markheiser, A.; Rid, M.; Biancu, S.; Gross, J.; Hoffmann, C. Physical factors influencing the oviposition behaviour of European grapevine moths Lobesia botrana and Eupoecilia ambiguella. J. Appl. Entomol. 2018, 142, 201–210. [Google Scholar] [CrossRef] [Green Version]
- EUNIS Habitat Type. European Environment Agency (EEA). Available online: https://eunis.eea.europa.eu/habitats.jsp/ (accessed on 29 September 2016).
- ArcGIS Desktop. Environmental System Research Institute. Available online: http://desktop.arcgis.com/de/ (accessed on 23 October 2019).
- Hesselbarth, M.H.K.; Sciaini, M.; With, K.A.; Wiegand, K.; Nowosad, J. Landscapemetrics: An open-source R tool to calculate landscape metrics. Ecography 2019, 42, 1648–1657. [Google Scholar] [CrossRef] [Green Version]
- The R Development Core Team. R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; SAGE Publications: Thousand Oaks, CA, USA, 2019. [Google Scholar]
- Wei, T.; Simko, V.R. Package “Corrplot”: Visualization of a Correlation Matrix. R Package Version 0.84, 2017. Available online: https://github.com/taiyun/corrplot (accessed on 13 April 2020).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P.; Heiberger, R.M.; Schuetzenmeister, A.; Scheibe, S.; Hothorn, M.T. Package ‘Multcomp’. Simultaneous Inference in General Parametric Models; Project for Statistical Computing: Vienna, Austria, 2016. [Google Scholar]
- Zuur, A.; Ieno, E.N.; Walker, N.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R. In Statistics for Biology and Health; Springer: New York, NY, USA, 2009. [Google Scholar]
- Langellotto, G.A.; Denno, R.F. Responses of invertebrate natural enemies to complex-structured habitats: A meta-analytical synthesis. Oecologia 2004, 139, 1–10. [Google Scholar] [CrossRef]
- Pennington, T.; Kraus, C.; Alakina, E.; Entling, M.H.; Hoffmann, C. Minimal pruning and reduced plant protection promote predatory mites in grapevine. Insects 2017, 8, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drieu, R.; Rusch, A. Conserving species-rich predator assemblages strengthens natural pest control in a climate warming context. Agric. For. Entomol. 2016, 19, 52–59. [Google Scholar] [CrossRef] [Green Version]
- Reinecke, A.; Thiéry, D. Grapevine insect pests and their natural enemies in the age of global warming. J. Pest. Sci. 2016, 89, 313–328. [Google Scholar] [CrossRef]
- Östman, Ö.; Ekbom, B.; Bengtsson, J. Landscape heterogeneity and farming practice influence biological control. Basic Appl. Ecol. 2001, 2, 365–371. [Google Scholar] [CrossRef]
- Kragten, S.; de Snoo, G.R. Bio-Support: Modelling the impact of landscape elements for pest control. Proc. Neth. Entomol. Soc. Meet. 2004, 15, 93–97. [Google Scholar]
- Bereswill, R.; Golla, B.; Streloke, M.; Schulz, R. Entry and toxicity of organic pesticides and copper in vineyard streams: Erosion rills jeopardise the efficiency of riparian buffer strips. AEE 2012, 146, 81–92. [Google Scholar] [CrossRef]
- Danne, A.; Thomson, L.J.; Sharley, D.J.; Penfold, C.M.; Hoffmann, A.A. Effects of Native Grass Cover Crops on Beneficial and Pest Invertebrates in Australian Vineyards. Environ. Entomol. 2010, 39, 970–978. [Google Scholar] [CrossRef] [PubMed]
- Muneret, L.; Auriol, A.; Bonnard, O.; Richart-Cervera, S.; Thiéry, D.; Rusch, A. Organic farming expansion drives natural enemy abundance but not diversity in vineyard-dominated landscapes. Ecol. Evol. 2019, 9, 13532–13542. [Google Scholar] [CrossRef] [PubMed]
- Serra, G.; Lentini, A.; Verdinelli, M.; Delrio, G. Effects of cover crop management on grape pests in a Mediterranean environment. IOBC-WPR Bull. 2006, 29, 209–214. [Google Scholar]
- Gonçalves, F.; Zina, V.; Crespo, L.; Oliveira, I.; Torres, L. Ants (Hymenoptera: Formicidae) and Spiders (Araneae) Co-occurring in the Ground of Vineyards from Douro Demarcated Region. Sociobiology 2017, 64, 404–416. [Google Scholar] [CrossRef] [Green Version]
- Happe, A.K.; Roquer-Beni, L.; Bosch, J.; Alins, G.; Mody, K. Earwigs and woolly apple aphids in integrated and organic apple orchards: Responses of a generalist predator and a pest prey to local and landscape factors. AEE 2018, 268, 44–51. [Google Scholar] [CrossRef]
- Ješovnik, A.; Blažević, I.; Lemić, D.; Pajač Živković, I. Ant fauna of annual and perennial crops. Appl. Ecol. Environ. Res. 2019, 17, 12709–12722. [Google Scholar] [CrossRef]
- Schmidt, M.H.; Tscharntke, T. The role of perennial habitats for Central European farmland spiders. AEE 2005, 105, 235–242. [Google Scholar] [CrossRef]
- Batáry, P.; Báldi, A.; Kleijn, D.; Tscharntke, T. Landscape-moderated biodiversity effects of agri-environmental management: A meta-analysis. Proc. R. Soc. Lond. B Biol. Sci. 2011, 278, 1894–1902. [Google Scholar] [CrossRef] [PubMed]
- Rusch, A.; Delbac, L.; Thiéry, D. Grape moth density in Bordeaux vineyards depends on local habitat management despite effects of landscape heterogeneity on their biological control. J. Appl. Ecol. 2017, 54, 1794–1803. [Google Scholar] [CrossRef] [Green Version]
- Frank, S.D.; Wratten, S.D.; Sandhu, H.S.; Shrewsbury, P.M. Video analysis to determine how habitat strata affects predator diversity and predation of Epiphyas postvittana (Lepidoptera: Tortricidae) in a vineyard. Biol. Control. 2007, 41, 230–236. [Google Scholar] [CrossRef]
- Buchholz, U.; Schruft, G. Räuberische Arthropoden auf Blüten und Früchten der Weinrebe (Vitis vinifera L.) als Antagonisten des Einbindigen Traubenwicklers (Eupoecilia ambiguella Hbn.) (Lep., Cochylidae). J. Appl. Entomol. 1994, 118, 31–37. [Google Scholar] [CrossRef]
- Huth, C.; Schirra, K.J.; Seitz, A.; Louis, F. A beneficial species becomes a pest—The common earwig Forficula auricularia (Linnaeus 1758). IOBC-WPRS Bull. 2011, 67, 249–256. [Google Scholar]
- Kehrli, P.; Karp, J.; Burdet, J.P.; Deneulin, P.; Danthe, E.; Lorenzini, F.; Linder, C. Impact of processed earwigs and their faeces on the aroma and taste of ‘Chasselas’ and ‘Pinot Noir’ wines. Vitis 2012, 51, 87–93. [Google Scholar]
- Bucher, R.; Hermann, J.D.; Schüepp, C.; Herzog, F.; Entling, M.H. Arthropod Colonisation of Trees in Fragmented Landscapes Depends on Species Traits. Open Ecol. J. 2010, 3, 111–117. [Google Scholar] [CrossRef] [Green Version]
- Nicholas, A.H.; Spooner-Hart, R.N.; Vickers, R.A. Abundance and natural control of the woolly aphid Eriosoma lanigerum in an Australian apple orchard. BioControl 2005, 50, 271–291. [Google Scholar] [CrossRef]
- Wilson, H.; Daane, K.M. Review of Ecologically-Based Pest Management in California Vineyards. Insects 2017, 8, 108. [Google Scholar] [CrossRef] [Green Version]
- Chong, C.S.; D’Alberto, C.F.; Thomson, L.J.; Hoffmann, A.A. Influence of native ants on arthropod communities in a vineyard. Agric. For. Entomol. 2010, 12, 223–232. [Google Scholar] [CrossRef]
- Grieshop, M.J.; Werling, B.; Buehrer, K.; Perrone, J.; Isaacs, R.; Landis, D. Big Brother is Watching: Studying Insect Predation in the Age of Digital Surveillance. Am. Entomol. 2012, 58, 172–182. [Google Scholar] [CrossRef] [Green Version]
- Stutz, S.; Entling, M.H. Effects of the landscape context on aphid-ant-predator interactions on cherry trees. Biol. Control. 2011, 57, 37–43. [Google Scholar] [CrossRef]
- Masoni, A.; Frizzi, F.; Brühl, C.; Zocchi, N.; Palchetti, E.; Chelazzi, G.; Santini, G. Management matters: A comparison of ant assemblages in organic and conventional vineyards. AEE 2017, 246, 175–183. [Google Scholar] [CrossRef]
- Thomson, L.J.; Sharley, D.J.; Hofmann, A.A. Beneficial organisms as bioindicators for environmental sustainability in the grape industry in Australia. Aust. J. Exp. Agric. 2007, 47, 404–411. [Google Scholar] [CrossRef]
- Varela, L.G.; Elkins, R.B.; Van Steenwyk, R.A. Evolution of Secondary Pests in California Pear Orchards under Mating Disruption for Codling Moth Control. 11th Int. Pear Symp. Acta Hort. 2011, 909, 531–542. [Google Scholar] [CrossRef]
- Weinberg, J.L.; Bunin, L.J.; Das, R. Application of the Industrial Hygiene Hierarchy of Controls to Prioritize and Promote Safer Methods of Pest Control: A Case Study. Public Health Rep. 2009, 124, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Maas, S.; Detzel, P.; Staudt, A. Gefährdungsanalyse der Heuschrecken Deutschlands. In Verbreitungsatlas, Gefährdungseinstufung und Schutzkonzepte; Bundesamt für Naturschutz: Bonn, Germany, 2002; 401p. [Google Scholar]
- Grabenweger, G.; Kehrli, P.; Schlick-Steiner, B.; Steiner, F.; Stolz, M.; Bacher, S. Predator complex of the horse chestnut leafminer Cameraria ohridella: Identification and impact assessment. J. Appl. Entomol. 2005, 129, 353–362. [Google Scholar] [CrossRef]
- Toepfer, S.; Kuhlmann, U. Survey for natural enemies of the invasive alien chrysomelid, Diabrotica virgifera virgifera, in Central Europe. BioControl 2004, 49, 385–395. [Google Scholar] [CrossRef]
- Vidano, C.; Arzone, A.; Anò, C. Researches on natural enemies of viticolous Auchenorrhyncha. In Meeting on Integrated Pest Control in Viticulture; A. A. Balkema: Rotterdam, The Netherlands, 1987; pp. 97–101. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organic | Integrated | |
|---|---|---|
| aAPTLc | 19.95 ± 6.88 | 10.61 ± 14.93 |
| Total number of pesticide applications | 9.69 ± 1.74 | 6.31 ± 2.30 |
| Copper applications | 8.88 ± 1.89 | 1.25 ± 0.86 |
| Sulfur applications | 9.19 ± 1.64 | 4.13± 1.89 |
| Applications with synthetic fungicides | – | 5.75 ± 2.67 |
| Insecticide applications | – | 0.25 ± 0.45 |
| Acaricide applications | 0.25 ± 0.45 | 0.38 ± 0.50 |
| Herbicide applications | – | 0.31 ± 0.60 |
| Compost fertilizer (m3/ha) | 3.28 ± 5.06 | 0.63 ± 2.50 |
| Artificial fertilizer (kg/ha) | – | 31.25 ± 68.01 |
| Tillage frequency in inter-rows | 1.00 ± 0.97 | 1.19 ± 1.47 |
| Tillage frequency in rows | 2.81 ± 0.83 | 2.50 ± 1.51 |
| Years without tillage in inter-rows | 2.63 ± 1.40 | 2.31 ± 1.99 |
| Mulching frequency in inter-rows | 2.00 ± 1.32 | 2.13 ± 0.96 |
| Rolling frequency in inter-rows | 0.44 ± 0.81 | 0.81 ± 1.33 |
| Inter-row vegetation height in spring (cm) | 11.45 ± 9.90 | 10.70 ± 8.97 |
| Row vegetation height in spring (cm) | 3.83 ± 2.90 | 5.93 ± 4.23 |
| Inter-row vegetation height in summer (cm) | 10.88 ± 7.43 | 10.13 ± 8.55 |
| Row vegetation height in summer (cm) | 8.90 ± 12.30 | 6.33 ± 4.84 |
| Vegetation cover in spring (%) in inter-rows | 82.38 ± 19.44 | 80.84 ± 20.03 |
| Vegetation cover in summer (%) in inter-rows | 61.70 ± 22.85 | 40.36 ± 25.43 |
| Bare ground cover in spring (%) in inter-rows | 12.59 ± 19.59 | 11.09 ± 14.74 |
| Bare ground cover in summer (%) in inter-rows | 11.75 ± 10.31 | 20.23 ± 11.14 |
| Species-Poor | Species-Rich | Spontaneous | |
|---|---|---|---|
| Tillage frequency in inter-rows | 1.00 ± 1.21 | 1.28 ± 1.38 | 0.83 ± 0.98 |
| Tillage frequency in rows | 2.33 ± 0.89 | 3.29 ± 1.2 | 1.83 ± 1.17 |
| Years without tillage in inter-rows | 1.42 ± 1.38 | 2.29 ± 1.14 | 1.33 ± 1.37 |
| Mulching frequency in inter-rows | 2.58 ± 1.24 | 1.71 ± 1.07 | 1.83 ± 0.75 |
| Rolling frequency in inter-rows | 0.42 ± 0.79 | 1.07 ± 1.38 | – |
| Inter-row vegetation height in spring (cm) | 16.17 ± 13.62 | 7.89 ± 3.09 | 6.63 ± 2.05 |
| Row vegetation height in spring (cm) | 4.45 ± 2.98 | 3.58 ± 3.07 | 4.72 ± 2.32 |
| Inter-row vegetation height in summer (cm) | 10.14 ± 8.54 | 11.18 ± 8.27 | 8.98 ± 4.33 |
| Row vegetation height in summer (cm) | 5.31 ± 3.04 | 9.38 ± 14.19 | 9.19 ± 5.99 |
| Vegetation cover in spring in inter-rows (%) | 75.79 ± 25.19 | 86.56 ± 8.76 | 75.87 ± 28.24 |
| Vegetation cover in summer in inter-rows (%) | 44.48 ± 24.88 | 51.98 ± 29.07 | 49.14 ± 22.15 |
| Bare ground cover in spring in inter-rows (%) | 13.75 ± 21.40 | 8.31 ± 7.64 | 13.06 ± 17.74 |
| Bare ground cover in summer in inter-rows (%) | 14.44 ± 9.79 | 16.23 ± 12.83 | 18.80 ± 10.13 |
| Herbicide applications in rows | 0.42 ± 0.67 | – | – |
| Management (Integrated/Organic) | Implementation of Cover-Crops (Species-Poor/Species-Rich/Spontaneous) | Proportion of Woody Seminatural Habitats | ||||
|---|---|---|---|---|---|---|
| χ² | p | χ² | p | χ² | p | |
| L. botrana eggs | 1.69 | 0.193 | 1.28 | 0.528 | 0.01 | 0.918 |
| L. botrana pupae | 11.76 | <0.001 | 6.46 | 0.040 | 0.52 | 0.469 |
| Individuals Captured on Photos | Individuals Involved in Predation or Parasitism | Predated L. botrana Pupae | Predation or Parasitism Incidents on L. botrana Eggs | |||||
|---|---|---|---|---|---|---|---|---|
| Organic | Integrated | Organic | Integrated | Organic | Integrated | Organic | Integrated | |
| Acari | ||||||||
| Anystidae | – | 2 (0) | – | 2 (0) | – | – | – | 2 (0) |
| Trombidiidae | – | 2 (1) | – | 2 (1) | – | – | – | 2 (1) |
| Araneae | ||||||||
| cf. Cheiracanthium sp. | 7 (7) | 9 (9) | 2 (2) | 3 (3) | – | – | 2 (2) | 3 (3) |
| cf. Drassodes sp. | – | 1 (1) | – | – | – | – | – | – |
| cf. Ebrechtella tricuspidata | 2 (0) | – | 1 (0) | – | – | – | 1 (0) | – |
| cf. Marpissa muscosa | – | 3 (0) | – | – | – | – | – | – |
| cf. Philodromus sp. | 1 (1) | – | 1 (1) | – | – | – | 1 (1) | – |
| cf. Pseudicius encarpatus | – | 2 (0) | – | – | – | – | – | – |
| cf. Salticidae | 1 | 6 (0) | – | – | – | – | – | – |
| Cheiracanthium sp. | 6 (6) | – | 1 (1) | – | 1 (1) | – | – | – |
| Salticidae | 1 (0) | – | 1 (0) | – | – | – | 1 (0) | – |
| Salticus sp. | 1 (0) | – | – | – | – | – | – | – |
| Coleoptera | ||||||||
| Carabidae | 6 (6) | 2 (2) | 2 (2) | 2 (2) | 7 (7) | 4 (4) | – | – |
| Coccinellidae | – | 1 (0) | – | – | – | – | – | – |
| Curculionidae | 1 (0) | – | – | – | – | – | – | – |
| sp. | 1 (0) | 1 (0) | – | – | – | – | – | – |
| Dermaptera | ||||||||
| Forficula auricularia | 38 (36) | 57 (51) | 16 (15) | 34 (30) | 16 (12) | 52 (46) | 9 (7) | 8 (6) |
| Diptera | ||||||||
| sp. | 13 (1) | 6 (1) | – | – | – | – | – | – |
| Hymenoptera | ||||||||
| Formicidae | 95 (24) | 118 (22) | 46 (20) | 13 (2) | 6 (1) | – | 27 (9) | 13 (2) |
| parasitic wasp | 17 (16) | 21 (15) | 16 (16) | 15 (11) | – | – | 16 (16) | 15 (11) |
| Lepidoptera | ||||||||
| sp. | – | 1 (0) | – | – | – | – | – | – |
| Neuroptera | ||||||||
| Chrysopidae (adult) | – | 1 (1) | – | 1 (1) | – | – | – | 1 (1) |
| Chrysopidae (larvae) | 2 (2) | – | 2 (2) | – | – | – | 2 (2) | – |
| Opiliones | ||||||||
| sp. | 1 (1) | – | – | – | – | – | – | – |
| Orthoptera | ||||||||
| Leptophytes albovittata | – | 1 (1) | – | – | – | – | – | – |
| Meconema meridionale | – | 2 (2) | – | 2 (2) | – | 9 (9) | – | – |
| Meconema thalassinum | – | 2 (2) | – | 2 (2) | – | 3 (3) | – | – |
| Oecanthus pellucens | 1 (1) | – | – | – | – | – | – | – |
| Phaneroptera falcata | 7 (6) | – | 5 (4) | – | 15 (13) | – | – | – |
| Phaneroptera nana | 4 (4) | – | 3 (3) | – | 13 (13) | – | – | – |
| Tettigonia viridissima | 1 (1) | 4 (4) | 1 (1) | 3 (3) | 5 (5) | 10 (10) | – | – |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reiff, J.M.; Kolb, S.; Entling, M.H.; Herndl, T.; Möth, S.; Walzer, A.; Kropf, M.; Hoffmann, C.; Winter, S. Organic Farming and Cover-Crop Management Reduce Pest Predation in Austrian Vineyards. Insects 2021, 12, 220. https://doi.org/10.3390/insects12030220
Reiff JM, Kolb S, Entling MH, Herndl T, Möth S, Walzer A, Kropf M, Hoffmann C, Winter S. Organic Farming and Cover-Crop Management Reduce Pest Predation in Austrian Vineyards. Insects. 2021; 12(3):220. https://doi.org/10.3390/insects12030220
Chicago/Turabian StyleReiff, Jo Marie, Sebastian Kolb, Martin H. Entling, Thomas Herndl, Stefan Möth, Andreas Walzer, Matthias Kropf, Christoph Hoffmann, and Silvia Winter. 2021. "Organic Farming and Cover-Crop Management Reduce Pest Predation in Austrian Vineyards" Insects 12, no. 3: 220. https://doi.org/10.3390/insects12030220
APA StyleReiff, J. M., Kolb, S., Entling, M. H., Herndl, T., Möth, S., Walzer, A., Kropf, M., Hoffmann, C., & Winter, S. (2021). Organic Farming and Cover-Crop Management Reduce Pest Predation in Austrian Vineyards. Insects, 12(3), 220. https://doi.org/10.3390/insects12030220