
Ant Diversity and Community Composition in Alpine Tree Line Ecotones



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Sampling
2.3. Statistics
3. Results
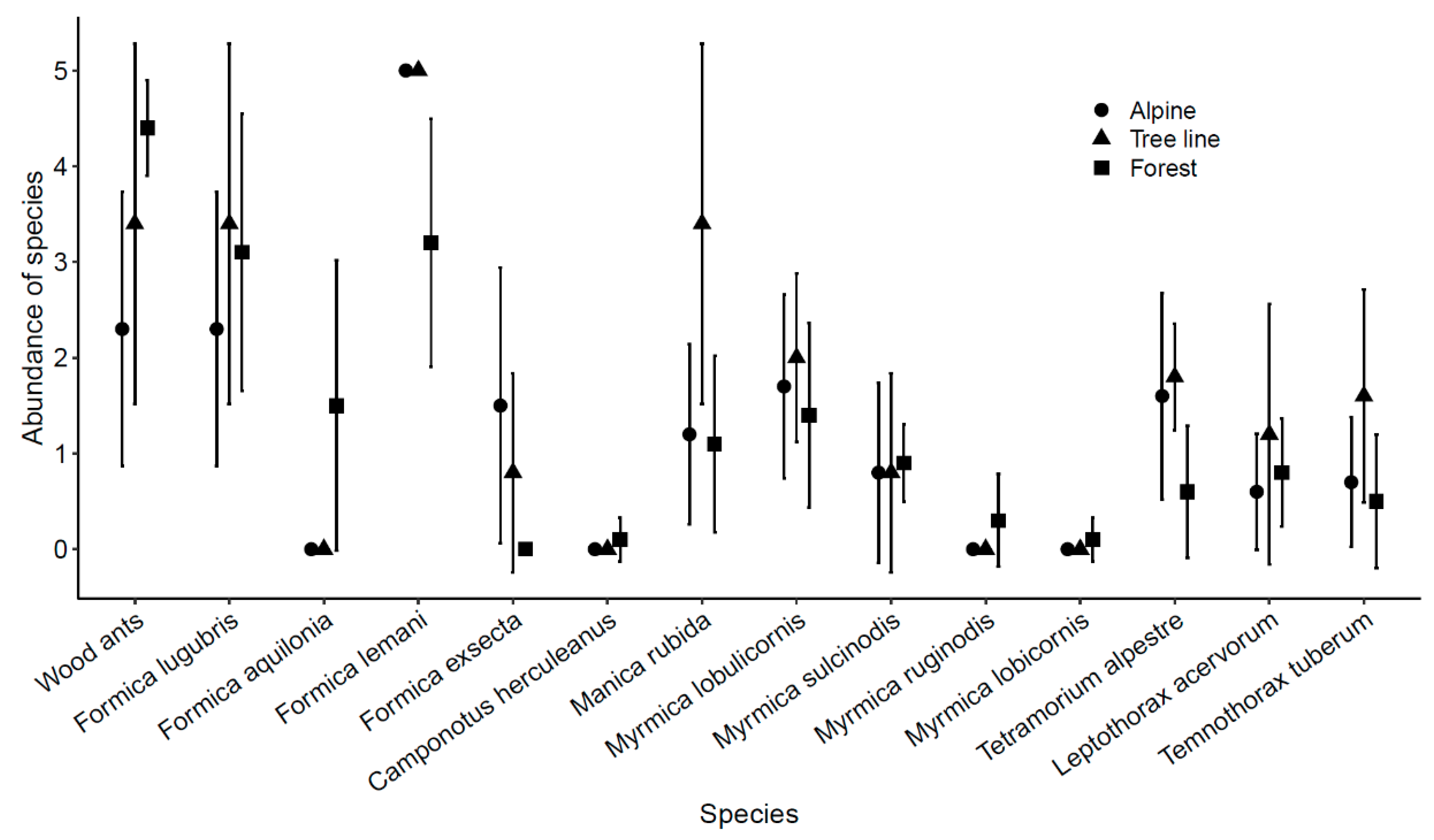
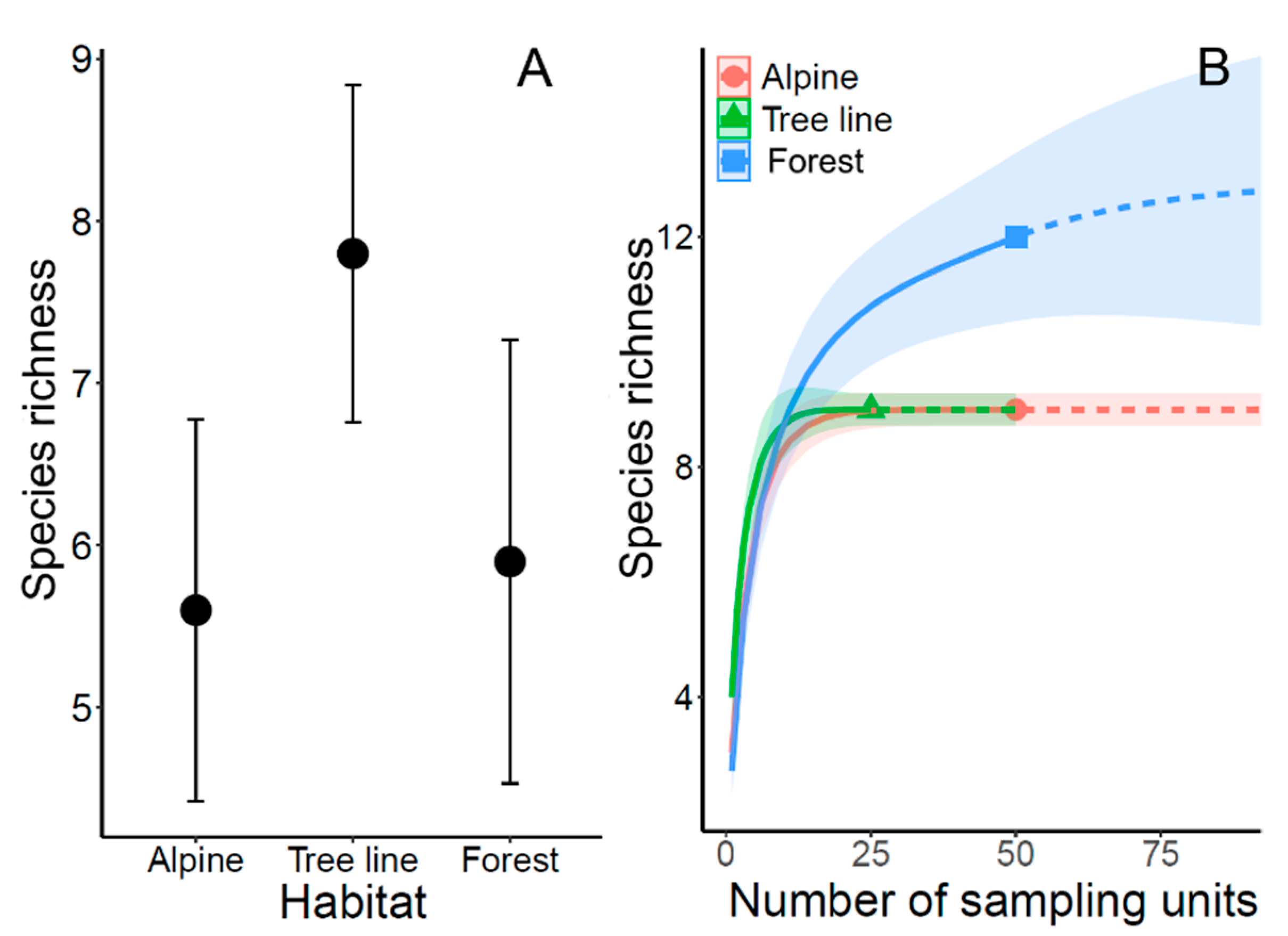
3.1. Species Richness and Diversity
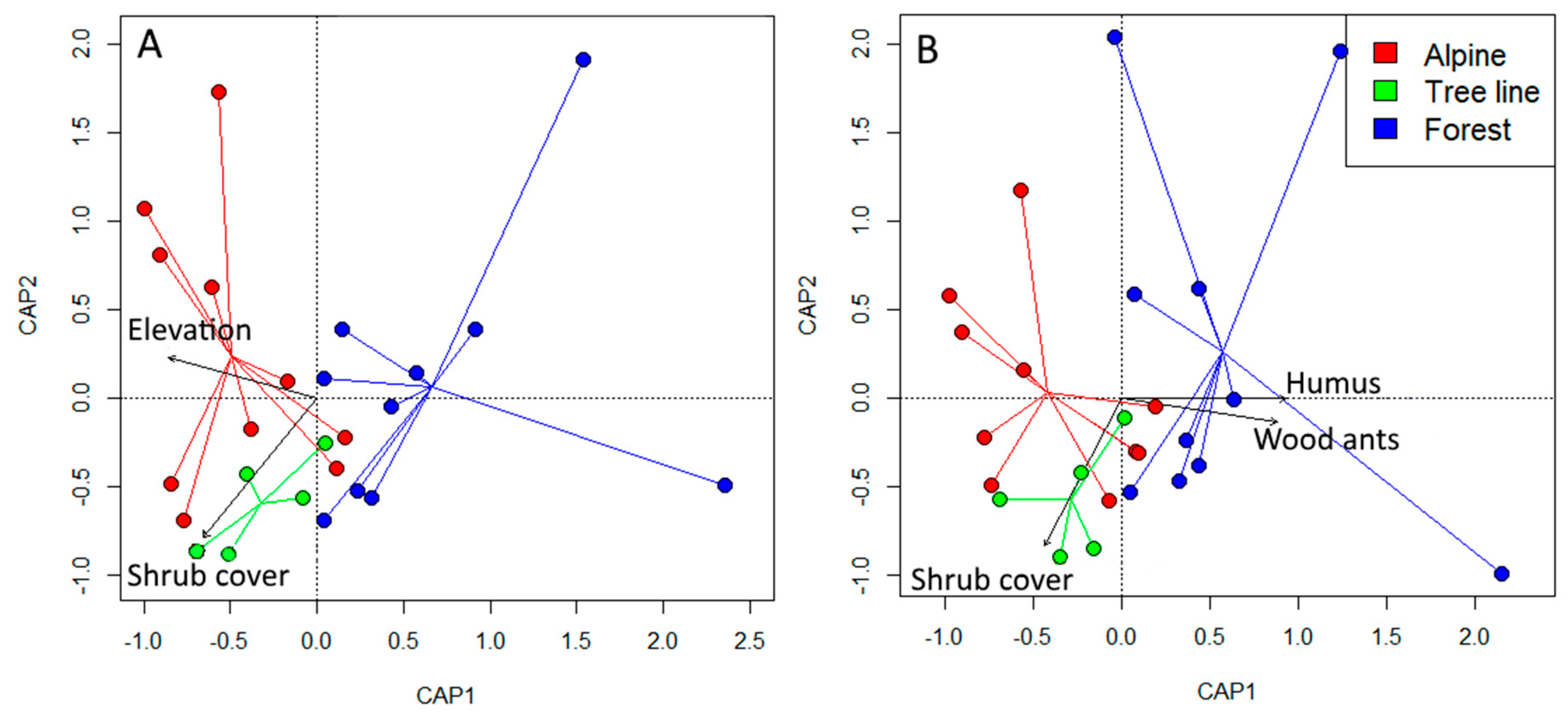
3.2. Community Composition
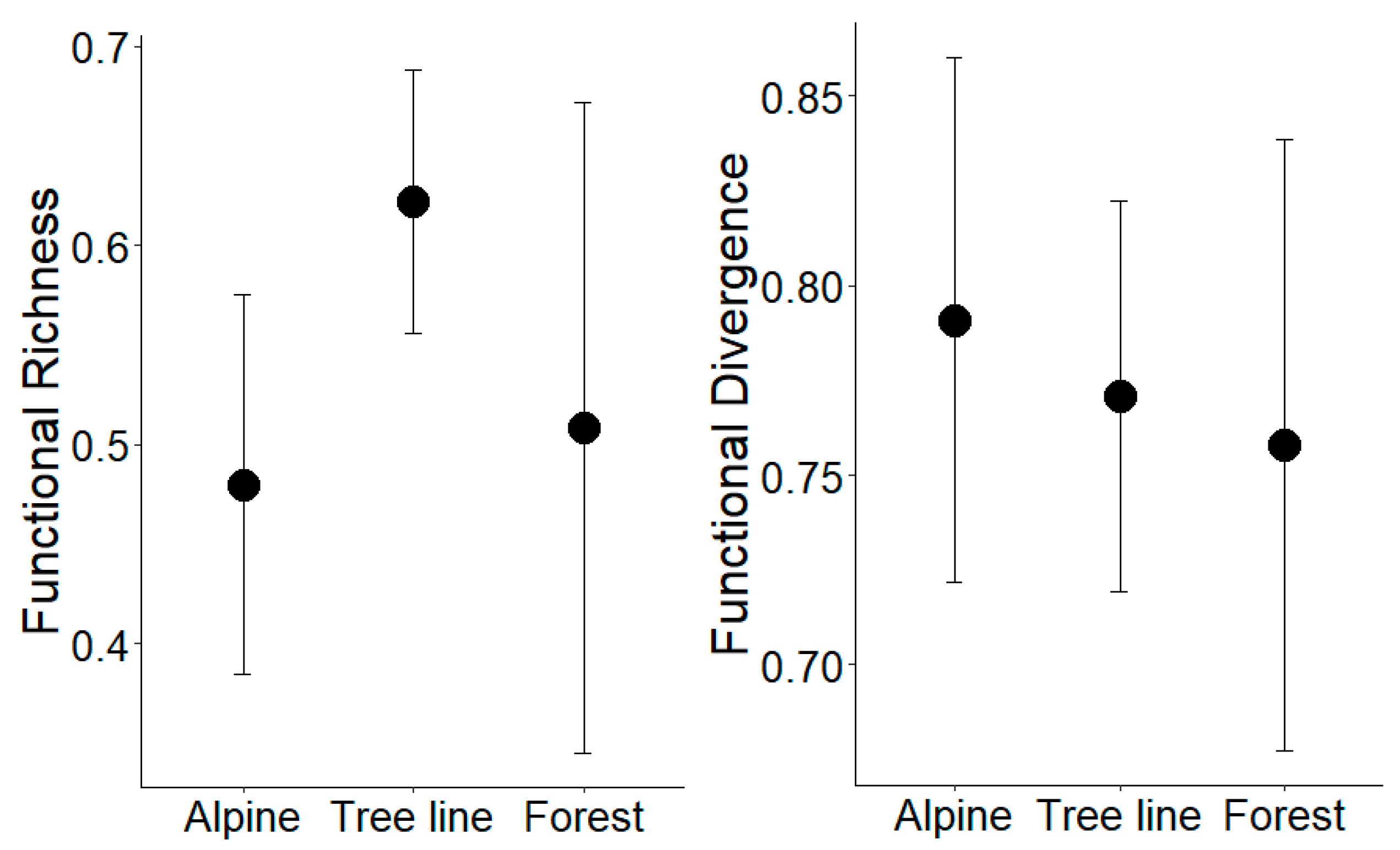
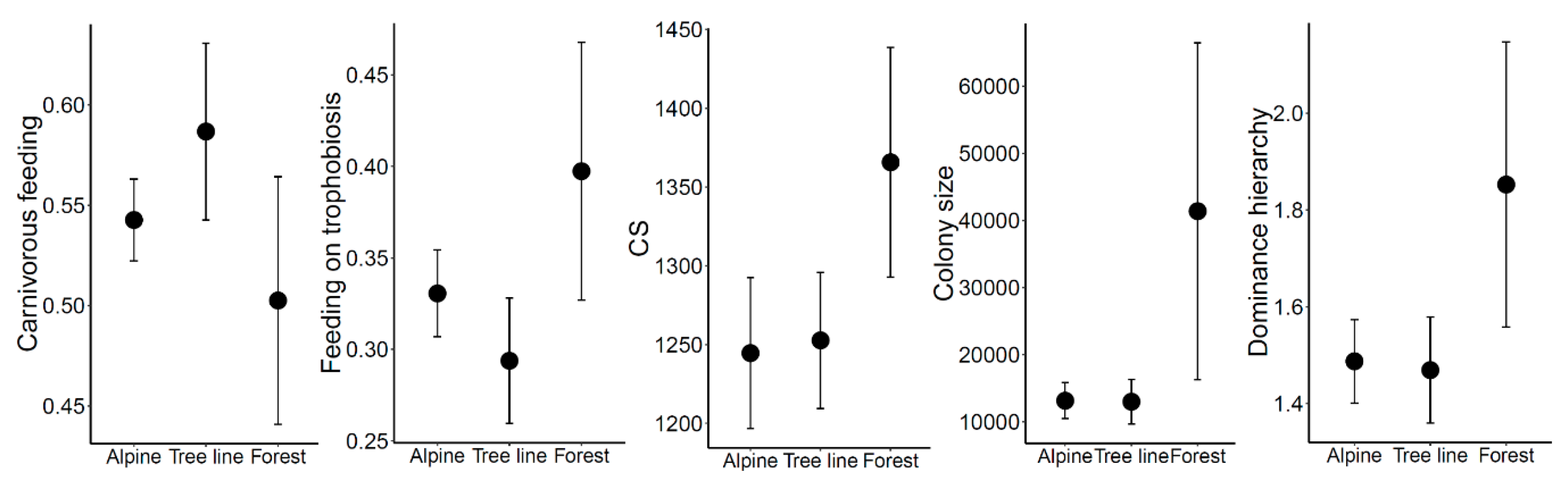
3.3. Functional Diversity and Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dunn, R.R.; Agosti, D.; Andersen, A.N.; Arnan, X.; Bruhl, C.A.; Cerdá, X.; Ellison, A.M.; Fisher, B.L.; Fitzpatrick, M.C.; Gibb, H.; et al. Climatic drivers of hemispheric asymmetry in global patterns of ant species richness. Ecol. Lett. 2009, 12, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Fisher, B.L. Biogeography. In Ant Ecology; Lach, L., Parr, C.L., Abbott, K.L., Eds.; Oxford University Press: New York, NY, USA, 2009; pp. 18–31. [Google Scholar]
- Francoeur, A. The ant fauna near the tree-line in northern Québec (Formicidae, Hymenoptera). Nordicana 1983, 47, 177–180. [Google Scholar]
- Heinze, J.; Hölldobler, B. Ants in the cold. Memorab. Zool. 1994, 48, 99–108. [Google Scholar]
- Reymond, A.; Purcell, J.; Cherix, D.; Guisan, A.; Pellissier, L. Functional diversity decreases with temperature in high elevation ant fauna: Functional diversity in high elevation ant. Ecol. Entomol. 2013, 38, 364–373. [Google Scholar] [CrossRef]
- Spotti, F.A.; Castracani, C.; Grasso, D.A.; Mori, A. Daily activity patterns and food preferences in an alpine ant community. Ethol. Ecol. Evol. 2015, 27, 306–324. [Google Scholar] [CrossRef]
- Geraghty, M.J.; Dunn, R.R.; Sanders, N.J. Body size, colony size, and range size in ants (Hymenoptera: Formicidae): Are patterns along elevational and latitudinal gradients consistent with Bergmann’s rule. Myrmecol. News 2007, 10, 51–58. [Google Scholar]
- Dunn, R.R.; Guenard, B.; Weiser, M.D.; Sanders, N.J.; Lach, L.; Parr, C.L.; Abbott, K.L. Geographic gradients. In Ant Ecology; Lach, L., Parr, C.L., Abbott, K.L., Eds.; Oxford University Press: New York, NY, USA, 2009; pp. 38–58. [Google Scholar]
- Körner, C. The use of ‘altitude’ in ecological research. Trends Ecol. Evol. 2007, 22, 569–574. [Google Scholar] [CrossRef]
- Glaser, F. Biogeography, diversity, and vertical distribution of ants (Hymenoptera: Formicidae) in Vorarlberg, Austria. Myrmecol. Nachr. 2006, 8, 263–270. [Google Scholar]
- Fontana, V.; Guariento, E.; Hilpold, A.; Niedrist, G.; Steinwandter, M.; Spitale, D.; Nascimbene, J.; Tappeiner, U.; Seeber, J. Species richness and beta diversity patterns of multiple taxa along an elevational gradient in pastured grasslands in the European Alps. Sci. Rep. 2020, 10, 12516. [Google Scholar] [CrossRef] [PubMed]
- Szewczyk, T.; McCain, C.M. A Systematic Review of Global Drivers of Ant Elevational Diversity. PLoS ONE 2016, 11, e0155404. [Google Scholar]
- Machac, A.; Janda, M.; Dunn, R.R.; Sanders, N.J. Elevational gradients in phylogenetic structure of ant communities reveal the interplay of biotic and abiotic constraints on diversity. Ecography 2011, 34, 364–371. [Google Scholar] [CrossRef]
- Sanders, N.J.; Lessard, J.-P.; Fitzpatrick, M.C.; Dunn, R.R. Temperature, but not productivity or geometry, predicts elevational diversity gradients in ants across spatial grains. Glob. Ecol. Biogeogr. 2007, 16, 640–649. [Google Scholar] [CrossRef]
- Bishop, T.R.; Robertson, M.P.; Van Rensburg, B.J.; Parr, C.L. Coping with the cold: Minimum temperatures and thermal tolerances dominate the ecology of mountain ants: Thermal tolerances of mountain ants. Ecol. Entomol. 2016, 42, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Guariento, E.; Strutzenberger, P.; Truxa, C.; Fiedler, K. The trinity of ecological contrasts: A case study on rich insect assemblages by means of species, functional and phylogenetic diversity measures. BMC Ecol. 2020, 20, 29. [Google Scholar] [CrossRef] [PubMed]
- Dethier, M.; Cherix, D. Note sur les Formicidae du Parc national suisse. Mitteilungen der Schweizerischen Entomol. Gesellschaft 1982, 55, 125–138. [Google Scholar]
- Seifert, B. The ecology of Central European non-arboreal ants—37 years of a broad-spectrum analysis under permanent taxonomic control. Soil Org. 2017, 89, 1–67. [Google Scholar]
- Wagner, H.C. Die Ameisen Kärntens; Naturwissenschaftlicher Verein für Kärnten: Graz, Austria, 2012; p. 462. [Google Scholar]
- La Sorte, F.A.; Jetz, W. Projected range contractions of montane biodiversity under global warming. Proc. R. Soc. B Biol. Sci. 2010, 277, 3401–3410. [Google Scholar] [CrossRef] [Green Version]
- Pauli, H.; Gottfried, M.; Grabherr, G. Effects of climate change on the alpine and nival vegetation of the Alps. J. Mt. Ecol. 2014, 7, 9–12. [Google Scholar]
- Körner, C.; Riedl, S. Alpine Treelines: Functional Ecology of the Global High Elevation Tree Limits; Springer: Basel, Switzerland, 2012; p. 357. [Google Scholar]
- Leonelli, G.; Pelfini, M.; Cella, U.M.D.; Garavaglia, V. Climate warming and the recent treeline shift in the european alps: The role of geomorphological factors in high-altitude sites. Ambio 2011, 40, 264–273. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, E.R.S.; Duarte, L.d.S.; Diehl, E.; Hartz, S.M. Edge effects on epigeic ant assemblages in a grassland–forest mosaic in southern Brazil. Acta Oecol. 2010, 36, 365–371. [Google Scholar] [CrossRef]
- Kotze, D.J.; Samways, M.J. No general edge effects for invertebrates at Afromontane forest/grassland ecotones. Biodivers. Conserv. 2001, 10, 443–466. [Google Scholar] [CrossRef]
- Steiner, F.M.; Schlick-Steiner, B.C. Edge effects on the diversity of ant assemblages in a xeric alluvial habitat in central Europe (Hymenoptera: Formicidae). Entomol. Gen. 2004, 1, 55–62. [Google Scholar] [CrossRef]
- Dauber, J.; Wolters, V. Edge effects on ant community structure and species richness in an agricultural landscape. Biodivers. Conserv. 2004, 13, 901–915. [Google Scholar] [CrossRef]
- Stuble, K.L.; Rodriguez-Cabal, M.A.; McCormick, G.L.; Jurić, I.; Dunn, R.R.; Sanders, N.J. Tradeoffs, competition, and coexistence in eastern deciduous forest ant communities. Oecologia 2013, 171, 981–992. [Google Scholar] [CrossRef] [PubMed]
- Lessard, J.; Stuble, K.L.; Sanders, N.J. Do Dominant Ants Affect Secondary Productivity, Behavior and Diversity in a Guild of Woodland Ants? Diversity 2020, 12, 460. [Google Scholar] [CrossRef]
- Sanders, N.J. Elevational gradients in ant species richness: Area, geometry, and Rapoport’s rule. Ecography 2002, 25, 25–32. [Google Scholar] [CrossRef]
- Guariento, E.; Rabl, D.; Ballini, S.; Fiedler, K. Wood ants ( Formicidae, Insecta ) at the alpine tree-line ecotone: Negative and positive associations with other arthropods. Gredleriana 2018, 18, 103–120. [Google Scholar]
- Vandegehuchte, M.L.; Wermelinger, B.; Fraefel, M.; Baltensweiler, A.; Düggelin, C.; Brändli, U.-B.; Freitag, A.; Bernasconi, C.; Cherix, D.; Risch, A.C. Distribution and habitat requirements of red wood ants in Switzerland: Implications for conservation. Biol. Conserv. 2017, 212, 366–375. [Google Scholar] [CrossRef]
- Savolainen, R.; Vepsäläinen, K. A Competition Hierarchy among Boreal Ants: Impact on Resource Partitioning and Community Structure. Oikos 1988, 51, 135. [Google Scholar] [CrossRef]
- Stadler, B.; Dixon, A. Mutualism: Ants and Their Insect Partners; Cambridge University Press: Cambridge, UK, 2008; p. 219. [Google Scholar]
- Domisch, T.; Risch, A.C.; Robinson, E.J.H. Wood ant foraging and mutualism with aphids. In Wood Ant Ecology and Conservation; Stockan, J.A., Robinson, E.J., Eds.; Cambridge University Press: Cambridge, UK, 2016; pp. 145–176. [Google Scholar]
- Tasser, E.; Tappeiner, U.; Cernusca, A. Südtirols Almen im Wandel: Ökologische Folgen von Landnutzungsänderungen; Europäische Akademie Bozen: Bozen, Italy, 2001; Volume 28, p. 269. [Google Scholar]
- Landolt, E. Ökologische Zeigerwerte Zur Schweizer Flora; Veröffentlichungen des Geobotanischen Institutes der Eidg. Techn. Hochschule, Stiftung Rübel: Zürich, Switzerland, 1977; p. 208. [Google Scholar]
- Guariento, E. Ecology of the Ant Community on the Alpine Tree Line Ecotone; University of Vienna: Vienna, Austria, 2018; p. 60. [Google Scholar]
- Delabie, J.H.C.; Fisher, B.L.; Majer, J.D.; Wright, I.W. Sampling effort and choice of methods. In Ants: Standard Methods for Measuring and Monitoring Biodiversity; Agosti, D., Majer, J.D., Alonso, L.E., Eds.; Smithsonian Institution Press: Washington, WA, USA, 2000; pp. 145–154. [Google Scholar]
- Hunter, M.D. Multiple approaches to estimating the relative importanceof top-down and bottom-up forces on insect populations: Experiments, life tables, and time-series analysis. Basic Appl. Ecol. 2001, 2, 295–309. [Google Scholar] [CrossRef]
- Bestelmeyer, B.T.; Agosti, D.; Alonso, L.E.; Brandão, C.R.F.; Brown, W.L.; Delabie, J.H.C.; Silvestre, R. Field techniques for the study of ground-dwelling ant: An overview, description, and evaluation. In Ants: Standard Methods for Measuring and Monitoring Biodiversity; Agosti, D., Majer, J.D., Alonso, L.E., Eds.; Smithsonian Institution Press: Washington, WA, USA, 2000; pp. 122–144. [Google Scholar]
- Fowler, D.; Lessard, J.-P.; Sanders, N.J. Niche filtering rather than partitioning shapes the structure of temperate forest ant communities. J. Anim. Ecol. 2014, 83, 943–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, M.K.; Mayr, A.; Röder, J.; Sanders, N.J.; Steffan-Dewenter, I. Variation in nutrient use in ant assemblages along an extensive elevational gradient on Mt Kilimanjaro. J. Biogeogr. 2014, 41, 2245–2255. [Google Scholar] [CrossRef]
- Tista, M.; Fiedler, K. How to evaluate and reduce sampling effort for ants. J. Insect Conserv. 2011, 15, 547–559. [Google Scholar] [CrossRef]
- Seifert, B. Die Ameisen Mittel-und Nordeuropas; Lutra Verlags-und Vertriebsgesellschaft Tauer: Görlitz, Germany, 2007; p. 368. [Google Scholar]
- Wagner, H.C.; Arthofer, W.; Seifert, B.; Muster, C.; Steiner, F.M.; Schlick-Steiner, B.C. Light at the end of the tunnel: Integrative taxonomy delimits cryptic species in the Tetramorium caespitum complex (Hymenoptera: Formicidae). Myrmecol. News 2017, 25, 95–129. [Google Scholar]
- R Core Team. R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria; Available online: http://www.R-project.org/ (accessed on 3 March 2021).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; p. 260. [Google Scholar]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: iNterpolation and EXTrapolation for Species Diversity; R Package Version 2.0.20. Available online: http://chao.stat.nthu.edu.tw/wordpress/software-download/ (accessed on 3 March 2021).
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Package Vegan: Community Ecology Package; R Package Version 2.5-6. Available online: https://CRAN.R-project.org/package=vegan (accessed on 3 March 2021).
- Clarke, K.R.; Somerfield, P.J.; Chapman, M.G. On resemblance measures for ecological studies, including taxonomic dissimilarities and a zero-adjusted Bray-Curtis coefficient for denuded assemblages. J. Exp. Mar. Bio. Ecol. 2006, 330, 55–80. [Google Scholar] [CrossRef]
- Wagenmakers, E.-J.; Farrell, S. AIC model selection using Akaike weights. Psychon. Bull. Rev. 2004, 11, 192–196. [Google Scholar] [CrossRef] [PubMed]
- Legendre, P.; Oksanen, J.; ter Braak, C.J.F. Testing the significance of canonical axes in redundancy analysis. Methods Ecol. Evol. 2011, 2, 269–277. [Google Scholar] [CrossRef]
- Villéger, S.; Mason, N.W.H.; Mouillot, D. New multidimensional functional diversity indices for a multifaceted framework in functional ecology. Ecology 2008, 89, 2290–2301. [Google Scholar] [CrossRef] [Green Version]
- Legendre, P.; Laliberté, E. A distance-based framework for measuring functional diversity from multiple traits. Ecology 2010, 91, 299–305. [Google Scholar]
- Laliberté, E.; Legendre, P.; Bill, S.; Shipley, B. FD: Measuring Functional Diversity from Multiple Traits, and Other Tools for Functional Ecology; R Package Version 1.0-12. Available online: https://CRAN.R-project.org/package=FD (accessed on 3 March 2021).
- Maggini, R.; Guisan, A.; Cherix, D. A stratified approach for modeling the distribution of a threatened ant species in the Swiss National Park. Biodivers. Conserv. 2002, 11, 2117–2141. [Google Scholar] [CrossRef]
- Stockan, J.A.; Robinson, E.J.H.; Trager, J.C.; Yao, I.; Seifert, B. Introducing wood ants: Evolution, phylogeny, indentification and distribution. In Wood Ant Ecology and Conservation; Stockan, J.A., Robinson, E.J., Eds.; Cambridge University Press: Cambridge, UK, 2016; pp. 1–37. [Google Scholar]
- Hölldobler, B.; Wilson, E.O. The Ants; Havard University Press: Cambridge, MA, USA, 1990; p. 733. [Google Scholar]
- Cerda, X.; Arnan, X.; Retana, J. Is competition a significant hallmark of ant (Hymenoptera: Formicidae) ecology. Myrmecol. News 2013, 18, 131–147. [Google Scholar]
- Johansson, T.; Gibb, H. Interspecific competition and coexistence between wood ants. In Wood Ant Ecology and Conservation; Stockan, J.A., Robinson, E.J., Eds.; Cambridge University Press: Cambridge, UK, 2016; pp. 123–144. [Google Scholar]
- Blüthgen, N.; Stork, N.E.; Fiedler, K. Bottom-up control and co-occurrence in complex communities: Honeydew and nectar determine a rainforest ant mosaic. Oikos 2004, 106, 344–358. [Google Scholar] [CrossRef]
- Hooper, D.U.; Solan, M.; Symstad, A.; Diaz, S.; Gessner, M.O.; Buchmann, N.; Degrange, V.; Grime, P.; Hulot, F.; Mermillod-Blondin, F.; et al. Species diversity, functional diversity and ecosystem functioning. Biodivers. Ecosyst. Funct. Synth. Perspect. 2002, 17, 195–208. [Google Scholar]
- Dayan, T.; Simberloff, D. Ecological and community-wide character displacement: The next generation. Ecol. Lett. 2005, 8, 875–894. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fixed Factors | Nr. Variables | Full Community | Community without Wood Ants | ||||
|---|---|---|---|---|---|---|---|
| AICc | w | R2 adj. | AICc | w | R2 adj. | ||
| Wood ants + Humus + Shrub + Elevation | 5 | 30.12 | 0.08 | 0.41 | |||
| Wood ants + Humus + Shrub | 4 | 28.49 | 0.19 | 0.40 | |||
| Wood ants + Humus + Elevation | 4 | 31.98 | 0.03 | 0.28 | |||
| Wood ant + Shrub + Elevation | 4 | 30.77 | 0.06 | 0.32 | |||
| Humus + Shrub + Elevation | 4 | 30.54 | 0.16 | 0.32 | 29.82 | 0.10 | 0.36 |
| Humus + Shrub | 3 | 31.69 | 0.09 | 0.22 | 29.87 | 0.09 | 0.30 |
| Humus + Elevation | 3 | 31.38 | 0.11 | 0.23 | 31.61 | 0.04 | 0.23 |
| Shrub + Elevation | 3 | 29.17 | 0.32 | 0.31 | 30.54 | 0.07 | 0.27 |
| Wood ants + Elevation | 3 | 32.14 | 0.03 | 0.21 | |||
| Wood ants + Humus | 2 | 30.53 | 0.07 | 0.27 | |||
| Wood ants + Shrub | 2 | 29.75 | 0.10 | 0.30 | |||
| Wood ants | 2 | 31.61 | 0.04 | 0.18 | |||
| Humus | 2 | 32.30 | 0.07 | 0.14 | 31.48 | 0.04 | 0.18 |
| Shrub | 2 | 32.70 | 0.06 | 0.13 | 32.86 | 0.02 | 0.13 |
| Elevation | 2 | 30.41 | 0.17 | 0.21 | 31.82 | 0.04 | 0.17 |
| Null Model | 1 | 34.53 | 0.02 | 0 | 34.45 | 0.01 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guariento, E.; Fiedler, K. Ant Diversity and Community Composition in Alpine Tree Line Ecotones. Insects 2021, 12, 219. https://doi.org/10.3390/insects12030219
Guariento E, Fiedler K. Ant Diversity and Community Composition in Alpine Tree Line Ecotones. Insects. 2021; 12(3):219. https://doi.org/10.3390/insects12030219
Chicago/Turabian StyleGuariento, Elia, and Konrad Fiedler. 2021. "Ant Diversity and Community Composition in Alpine Tree Line Ecotones" Insects 12, no. 3: 219. https://doi.org/10.3390/insects12030219
APA StyleGuariento, E., & Fiedler, K. (2021). Ant Diversity and Community Composition in Alpine Tree Line Ecotones. Insects, 12(3), 219. https://doi.org/10.3390/insects12030219