
Concentration-Dependent Feeding Deterrence to 20-Hydroxyecdysone for Three Subterranean Termite Species (Blattodea: Rhinotermitidae)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Termites
2.2. Media Pad Preparation
2.3. Choice Test
2.4. Statistical Analysis
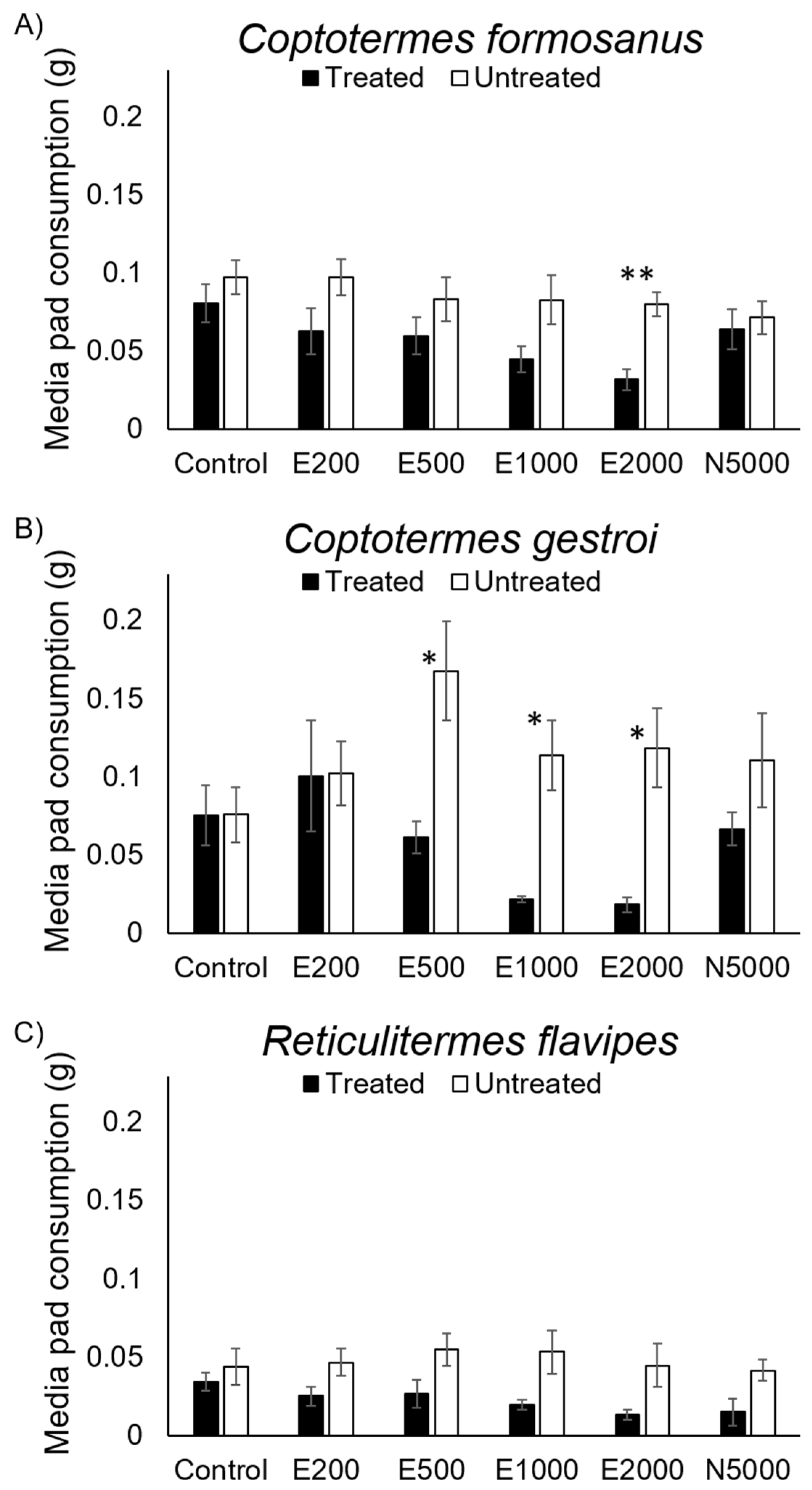
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rust, M.K.; Su, N.-Y. Managing social insects of urban importance. Annu. Rev. Entomol. 2012, 57, 355–375. [Google Scholar] [CrossRef] [PubMed]
- Su, N.-Y. Development of saits for population management of subterranean termites. Annu. Rev. Entomol. 2019. [Google Scholar] [CrossRef]
- Evans, T.A.; Iqbal, N. Termite (order Blattodea, infraorder Isoptera) baiting 20 years after commercial release. Pest Manag. Sci. 2015, 71, 897–906. [Google Scholar] [CrossRef] [PubMed]
- Su, N.-Y. Field evaluation of a hexaflumuron bait for population suppression of subterranean termites (Isoptera: Rhinotermitidae). J. Econ. Entomol. 1994, 87, 389–397. [Google Scholar] [CrossRef]
- Su, N.-Y.; Scheffrahn, R.H. Laboratory evaluation of two chitin synthesis inhibitors, hexaflumuron and diflubenzuron, as bait toxicants against Formosan and eastern subterranean termites (Isoptera: Rhinotermitidae). J. Econ. Entomol. 1993, 86, 1453–1457. [Google Scholar] [CrossRef]
- Su, N.-Y.; Scheffrahn, R.H. A review of subterranean termite control practices and prospects for integrated pest management programmes. Integr. Pest Manag. Rev. 1998, 3, 1–13. [Google Scholar] [CrossRef]
- Su, N.-Y.; Scheffrahn, R.H. Fate of subterranean termite colonies (Isoptera) after bait applications—An update and review. Sociobiology 1996, 26, 253–275. [Google Scholar]
- Grace, J.K.; Su, N.-Y. Evidence supporting the use of termite baiting systems for long-term structural protection (Isoptera). Sociobiology 2001, 37, 301–310. [Google Scholar]
- Su, N.-Y.; Guidry, E.; Mullins, A.J.; Cotonne, C. Reinvasion dynamics of subterranean termites (Isoptera: Rhinotermitidae) following the elimination of all detectable colonies in a large area. J. Econ. Entomol. 2016, 109, 809–814. [Google Scholar] [CrossRef]
- Chouvenc, T.; Su, N.-Y. Subterranean Termites feeding on CSI baits for a short duration still results in colony elimination. J. Econ. Entomol. 2017, 110, 2534–2538. [Google Scholar] [CrossRef] [PubMed]
- Bernard, S.; Osbrink, W.; Su, N.-Y. Response of the Formosan subterranean termite to neighboring con-specific populations after baiting with noviflumuron. J. Econ. Entomol. 2017, 110, 575–583. [Google Scholar] [CrossRef]
- Su, N.-Y.; Scheffrahn, R.H. Comparative effects of two chitin synthesis inhibitors, hexaflumuron and lufenuron, in a bait matrix against subterranean termites (Isoptera: Rhinotermitidae). J. Econ. Entomol. 1996, 89, 1156–1160. [Google Scholar] [CrossRef]
- Su, N.-Y.; Monteagudo, E.J. Hyperecdysonism in the Formosan subterranean termite and eastern subterranean termite (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2017, 110, 1736–1739. [Google Scholar] [CrossRef] [PubMed]
- Sheets, J.J.; Karr, L.L.; Dripps, J.E. Kinetics of uptake, clearance, transfer, and metabolism of hexaflumuron by eastern subterranean termites (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2000, 93, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Haagsma, K.A.; Rust, M.K. Effect of hexaflumuron on mortality of the Western subterranean termite (Isoptera: Rhinotermitidae) during and following exposure and movement of hexaflumuron in termite groups. Pest Manag. Sci. Former. Pestic. Sci. 2005, 61, 517–531. [Google Scholar] [CrossRef]
- Raina, A.; Park, Y.I.; Gelman, D. Molting in workers of the Formosan subterranean termite Coptotermes formosanus. J. Insect Physiol. 2008, 54, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Xing, L.; Chouvenc, T.; Su, N.-Y. Behavioral and histological changes in the Formosan subterranean termite (Isoptera: Rhinotermitidae) induced by the chitin synthesis inhibitor noviflumuron. J. Econ. Entomol. 2014, 107, 741–747. [Google Scholar] [CrossRef] [Green Version]
- Kakkar, G.; Su, N.-Y. Molting drives mortality in foraging populations of Formosan subterranean termites (Isoptera: Rhinotermitidae) baited with a chitin synthesis inhibitor, noviflumuron. Pest Manag. Sci. 2018, 74, 219–224. [Google Scholar] [CrossRef]
- Kakkar, G.; Osbrink, W.; Su, N.-Y. Molting site fidelity accounts for colony elimination of the Formosan subterranean termites (Isoptera: Rhinotermitidae) by chitin synthesis inhibitor baits. Sci. Rep. 2018, 8, 1259. [Google Scholar] [CrossRef] [Green Version]
- Carnohan, L.; Su, N.-Y. Mortality time trends of three subterranean termite species when exposed to 20-hydroxyecdysone, alone and in combination with noviflumuron. J. Econ. Entomol. 2018, 111, 2312–2316. [Google Scholar] [CrossRef]
- Su, N.-Y.; Lees, M. Biological activities of a bait toxicant for population management of subterranean termites. In Proceedings of the Pesticides in household, structural and residential pest management. Am. Chem. Soc. Symp. Ser. 2009, 1015, 87–96. [Google Scholar]
- Su, N.-Y.; Tamashiro, M.; Yates, J.R.; Haverty, M. Effect of behavior on the evaluation of insecticides for prevention of or remedial control of the Formosan subterranean termite. J. Econ. Entomol. 1982, 75, 188–193. [Google Scholar] [CrossRef]
- Su, N.-Y.; Scheffrahn, R.H. Toxicity and feeding deterrency of a dihaloalkyl arylsulfone biocide, A-9248, against the Formosan subterranean termite (Isoptera: Rhinotermitidae). J. Econ. Entomol. 1988, 81, 850–854. [Google Scholar] [CrossRef]
- Glenn, G.J.; Gold, R.E. Evaluation of commercial termite baiting systems for pest management of the Formosan subterranean termite (Isoptera: Rhinotermitidae). In Proceedings of the 4th International Conference on Urban Pests, Charleston, SC, USA, 7–10 July 2002; pp. 325–334. [Google Scholar]
- Su, N.-Y.; Scheffrahn, R.H.; Ban, P.M. Effects of sulfluramid-treated bait blocks on field colonies of the Formosan subterranean termite (Isoptera: Rhinotermitidae). J. Econ. Entomol. 1995, 88, 1343–1348. [Google Scholar] [CrossRef]
- Dhadialla, T.S.; Retnakaran, A.; Smagghe, G. Insect growth-and development-disrupting insecticides. In Insect Development: Morphogenesis, Molting and Metamorphosis; Academic Press: Amsterdam, The Netherlands, 2009; pp. 679–740. ISBN 0123751365. [Google Scholar]
- Tamashiro, M.; Fujii, J.K.; Lai, P.-Y. A simple method to observe, trap, and prepare large numbers of subterranean termites for laboratory and field experiments. Environ. Entomol. 1973, 2, 721–722. [Google Scholar] [CrossRef]
- Su, N.-Y.; Scheffrahn, R.H. A method to access, trap, and monitor field populations of the Formosan subterranean termite (Isoptera: Rhinotermitidae). Sociobiology 1986, 12, 299–304. [Google Scholar]
- Haverty, M.I. The proportion of soldiers in termite colonies: A list and a bibliography. Sociobiology 1977, 2, 199–216. [Google Scholar]
- IBM SPSS Inc. SPSS Statistics for Windows. IBM Corp. Released 2012 2016. [Google Scholar] [CrossRef]
- Cornelius, M.L.; Daigle, D.J.; Connick, W.J., Jr.; Parker, A.; Wunch, K. Responses of Coptotermes formosanus and Reticulitermes flavipes (Isoptera: Rhinotermitidae) to three types of wood rot fungi cultured on different substrates. J. Econ. Entomol. 2002, 95, 121–128. [Google Scholar] [CrossRef]
- Su, N.-Y. Technological needs for sustainable termite management. Sociobiology 2011, 58, 229. [Google Scholar]
- Chouvenc, T. Comparative impact of chitin synthesis inhibitor baits and non-repellent liquid termiticides on subterranean termite colonies over foraging distances: Colony elimination versus localized termite exclusion. J. Econ. Entomol. 2018, 111, 2317–2328. [Google Scholar] [CrossRef]
- Vargo, E.L.; Parman, V. Effect of fipronil on subterranean termite colonies (Isoptera: Rhinotermitidae) in the field. J. Econ. Entomol. 2012, 105, 523–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osbrink, W.L.A.; Cornelius, M.L.; Showler, A.T.; Pound, J.M. Effects of a fipronil spot treatment on field colonies of Coptotermes formosanus (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2014, 107, 727–740. [Google Scholar] [CrossRef]
- Su, N.-Y. Response of the Formosan subterranean termites (Isoptera: Rhinotermitidae) to baits or nonrepellent termiticides in extended foraging arenas. J. Econ. Entomol. 2005, 98, 2143–2152. [Google Scholar] [CrossRef] [PubMed]
- Ngee, P.-S.; Yoshimura, T.; Lee, C.-Y. Foraging populations and control strategies of subterranean termites in the urban environment, with special reference to baiting. Jpn. J. Environ. Entomol. Zool. 2004, 15, 197–215. [Google Scholar]



{kind=link}
{kind=link}
| Species | ||||
|---|---|---|---|---|
| C. formosanus | C. gestroi | R. flavipes | ||
| Treatment | Control | 178.38 ± 24.15 | 151.50 ± 39.58 | 78.82 ± 18.57 |
| 20E 200 ppm | 160.65 ± 27.59 | 203.03 ± 58.46 | 72.48 ± 16.12 | |
| 20E 500 ppm | 143.13 ± 27.50 | 229.38 ± 38.27 | 81.90 ± 17.62 | |
| 20E 1000 ppm | 127.80 ± 25.47 | 135.80 ± 27.86 | 73.16 ± 17.47 | |
| 20E 2000 ppm | 111.98 ± 15.72 | 137.23 ± 27.52 | 58.60 ± 18.03 | |
| Noviflumuron | 135.88 ± 24.68 | 177.50 ± 46.61 | 50.98 ± 14.82 | |
| Statistics | Mean square | 0.00248688 | 0.00594039 | 0.00073422 |
| F | 0.99 | 0.79 | 0.52 | |
| p | 0.4520 | 0.5681 | 0.7577 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carnohan, L.; Lee, S.-B.; Su, N.-Y. Concentration-Dependent Feeding Deterrence to 20-Hydroxyecdysone for Three Subterranean Termite Species (Blattodea: Rhinotermitidae). Insects 2021, 12, 218. https://doi.org/10.3390/insects12030218
Carnohan L, Lee S-B, Su N-Y. Concentration-Dependent Feeding Deterrence to 20-Hydroxyecdysone for Three Subterranean Termite Species (Blattodea: Rhinotermitidae). Insects. 2021; 12(3):218. https://doi.org/10.3390/insects12030218
Chicago/Turabian StyleCarnohan, Lucas, Sang-Bin Lee, and Nan-Yao Su. 2021. "Concentration-Dependent Feeding Deterrence to 20-Hydroxyecdysone for Three Subterranean Termite Species (Blattodea: Rhinotermitidae)" Insects 12, no. 3: 218. https://doi.org/10.3390/insects12030218
APA StyleCarnohan, L., Lee, S. -B., & Su, N. -Y. (2021). Concentration-Dependent Feeding Deterrence to 20-Hydroxyecdysone for Three Subterranean Termite Species (Blattodea: Rhinotermitidae). Insects, 12(3), 218. https://doi.org/10.3390/insects12030218