
Colony Suppression and Possible Colony Elimination of the Subterranean Termites Coptotermes formosanus and Reticulitermes speratus by Discontinuous Soil Treatment Using a Diluent of Fipronil Suspension Concentrate

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
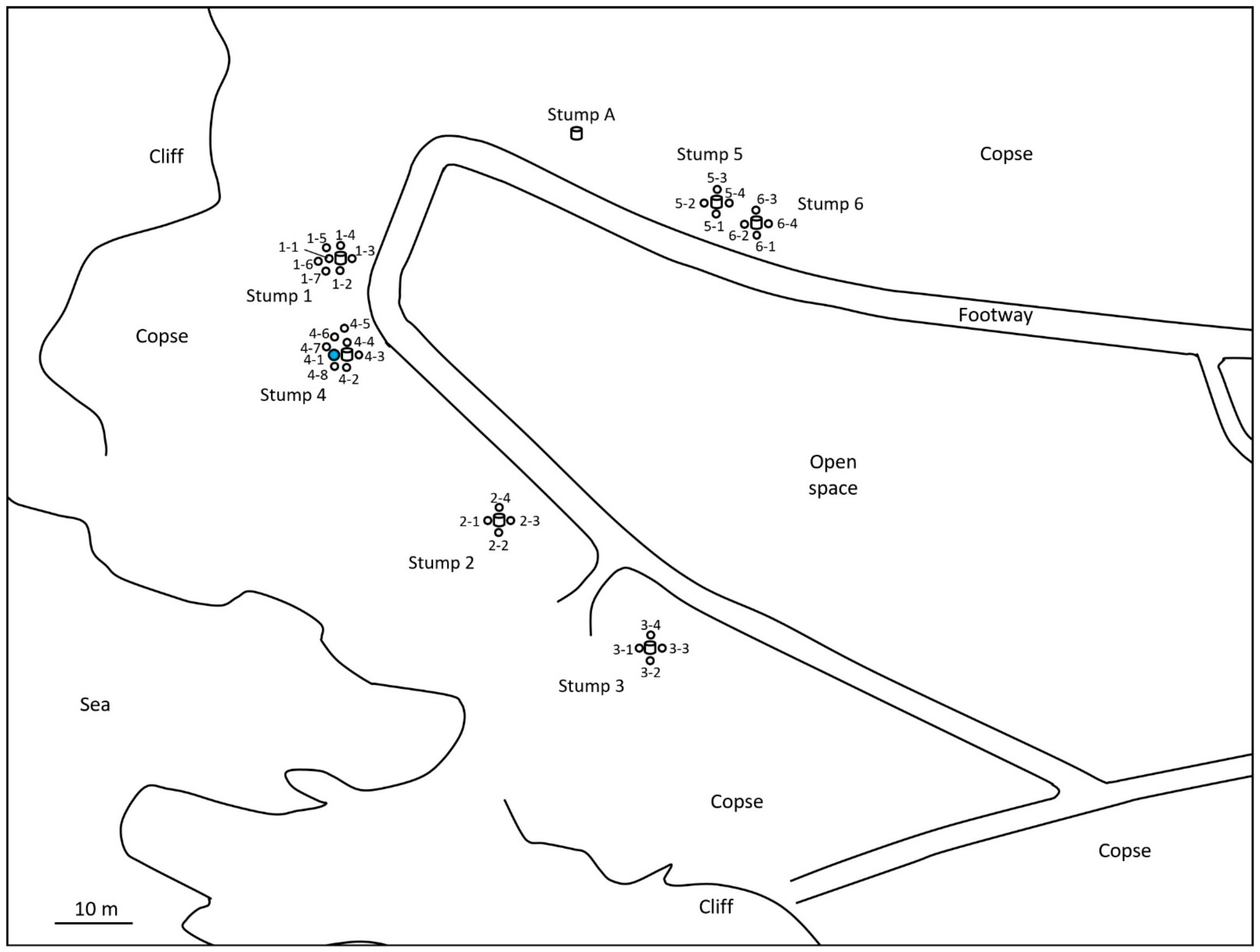
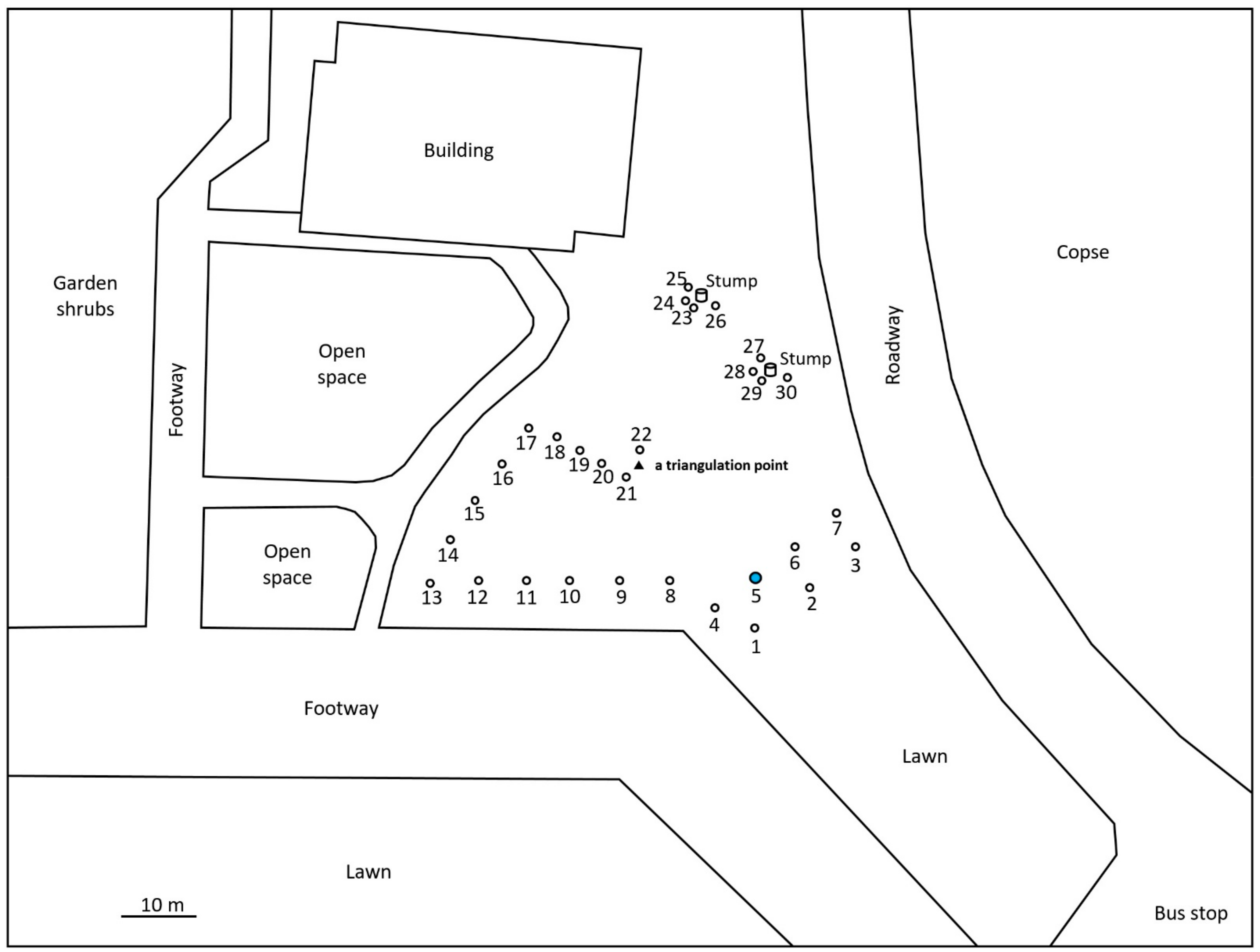
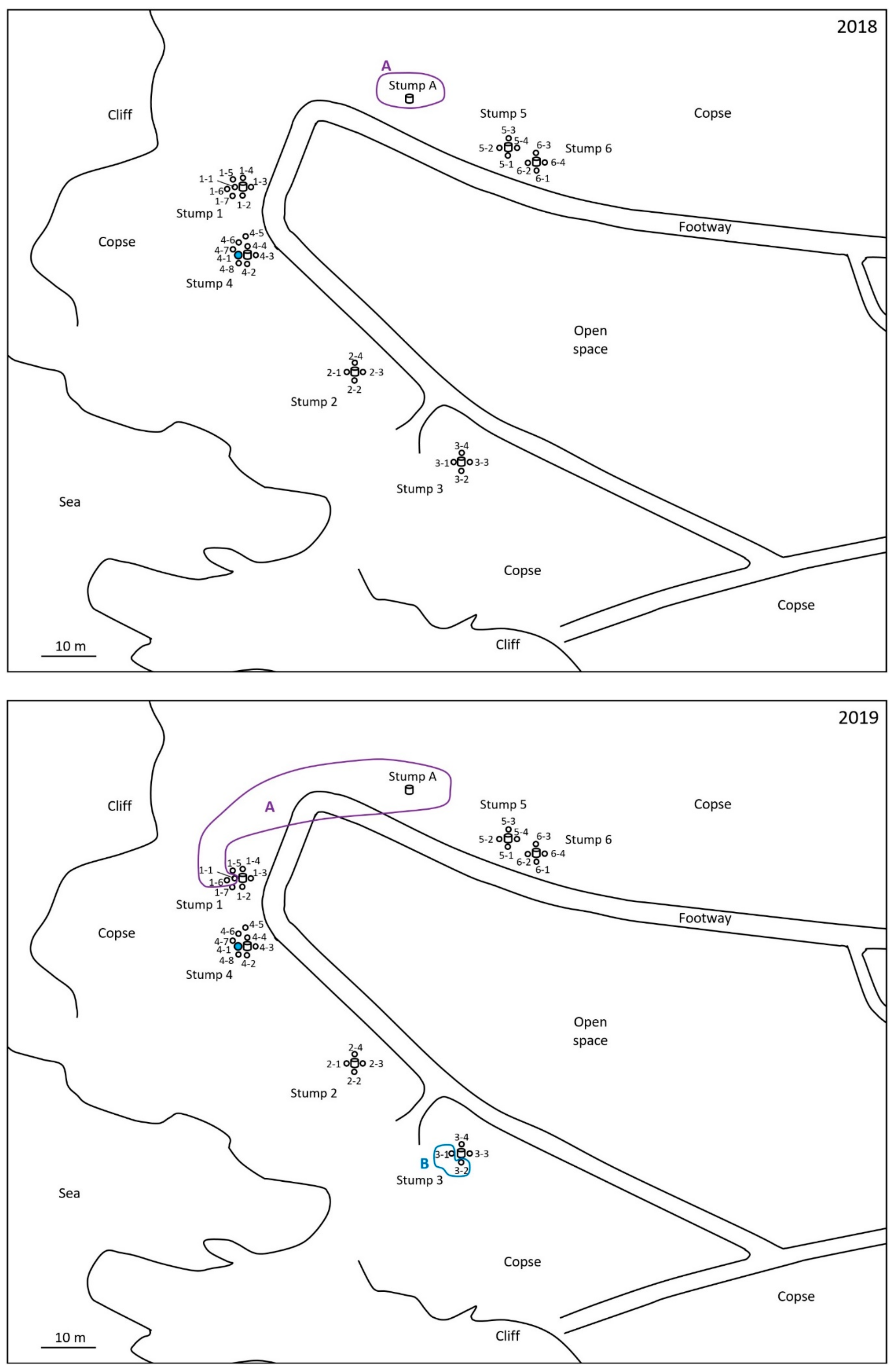
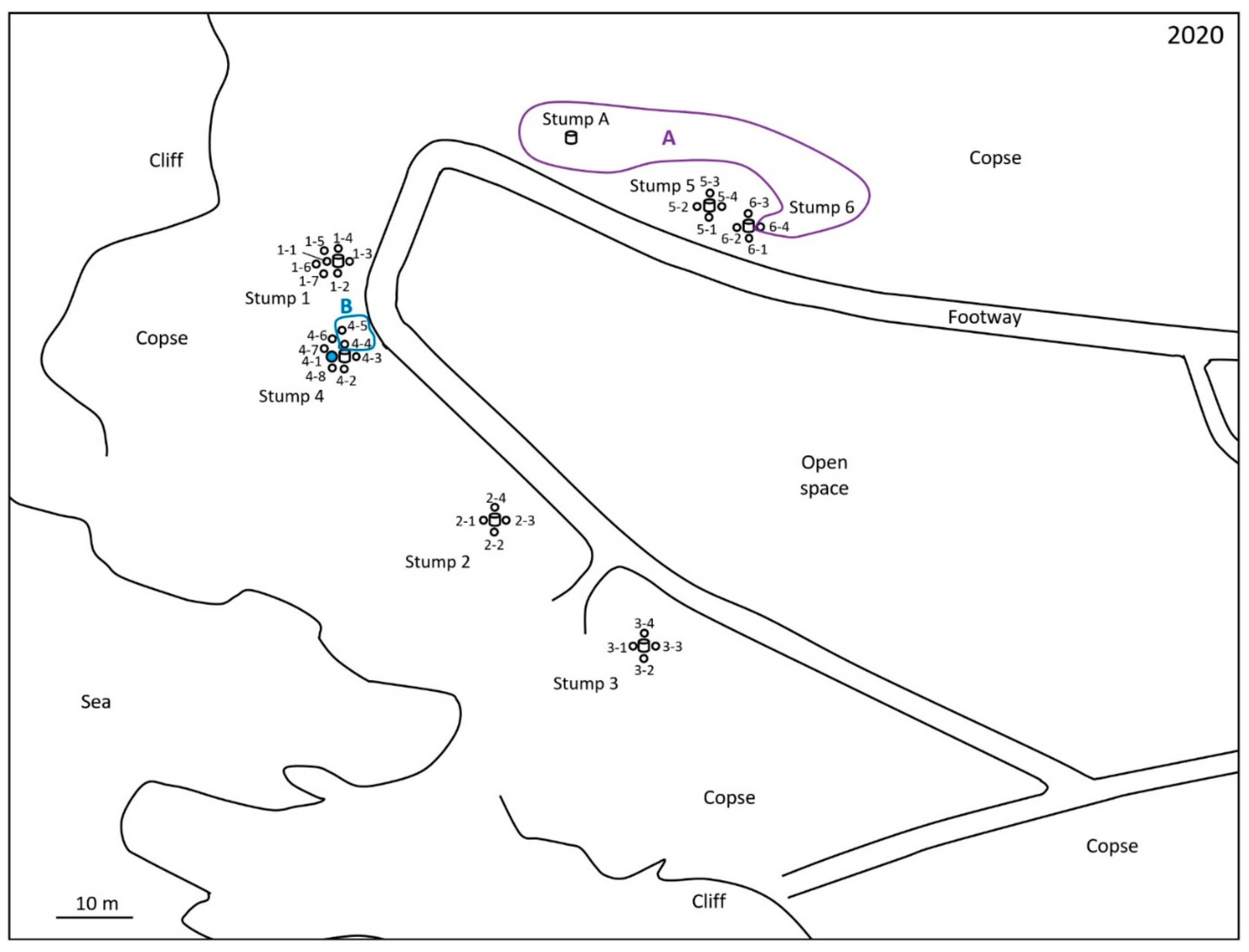
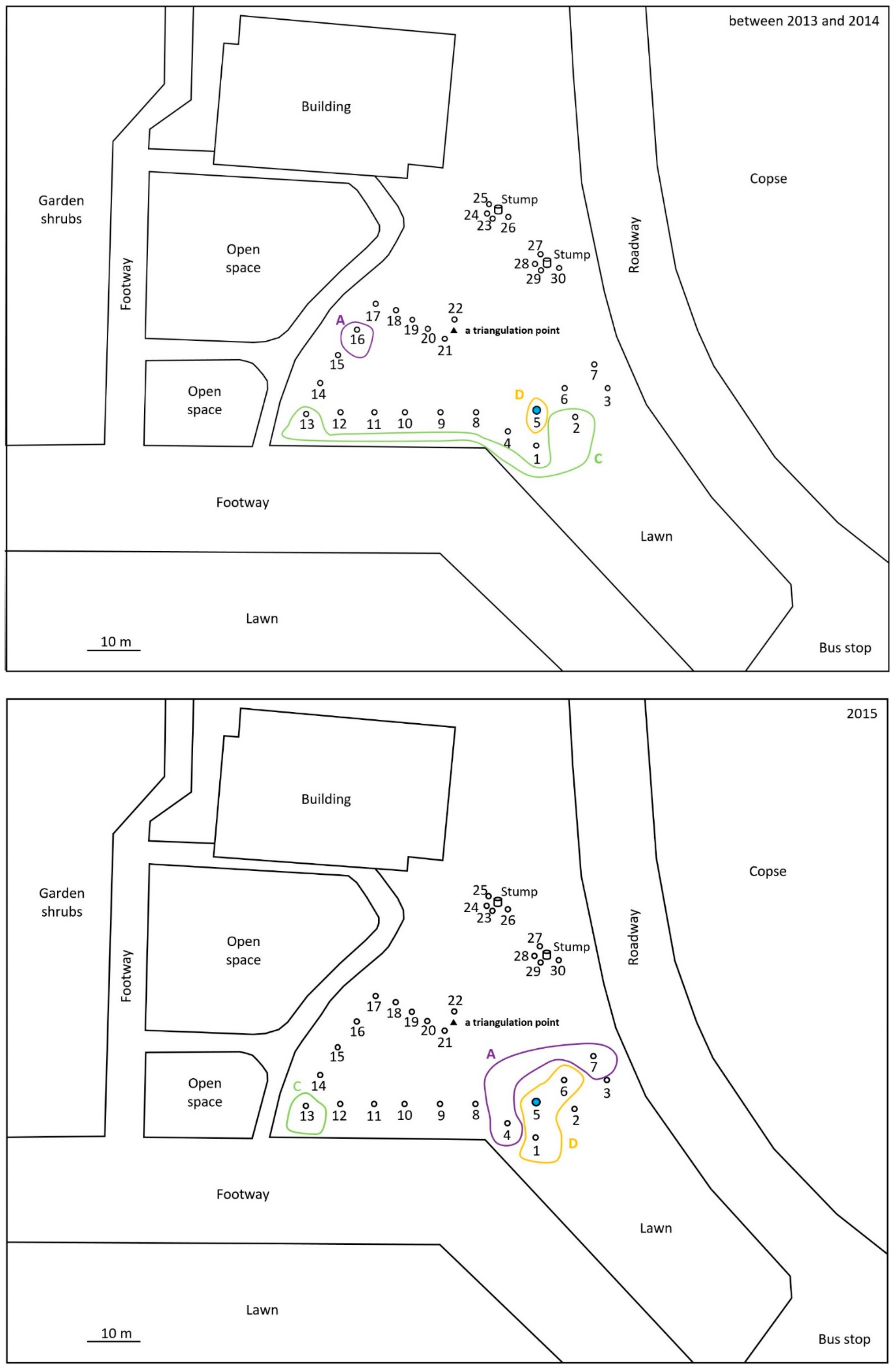
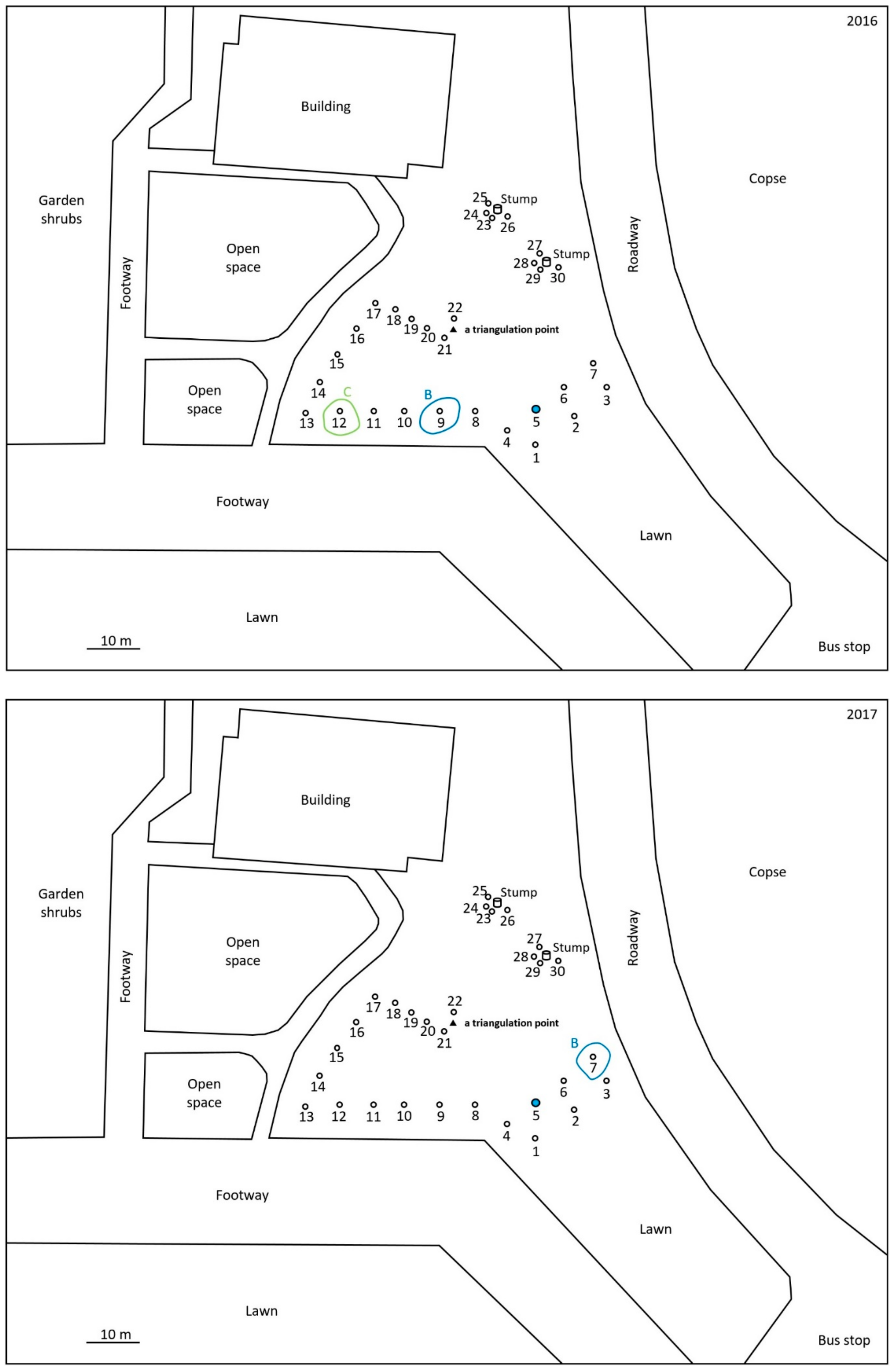
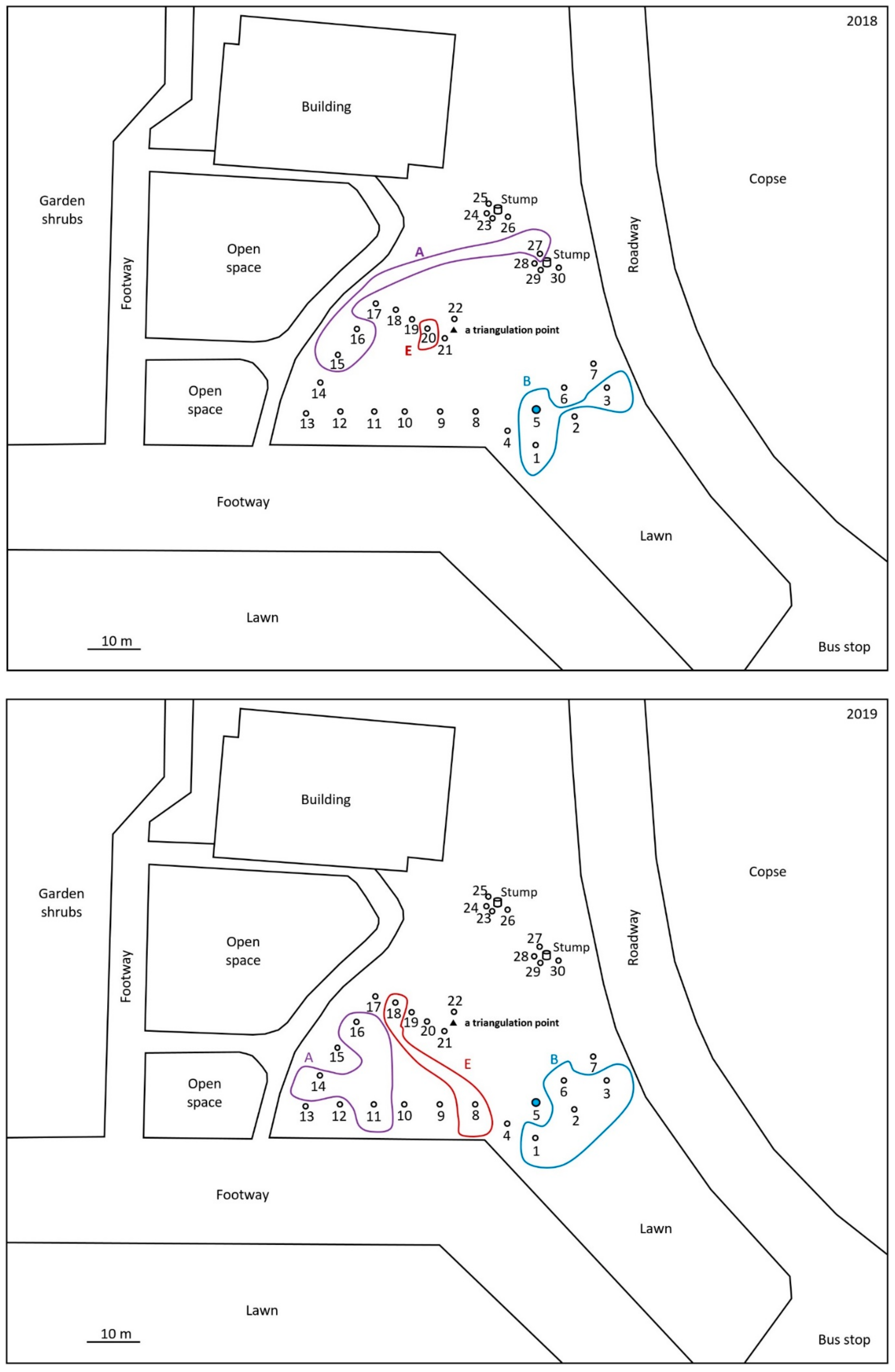
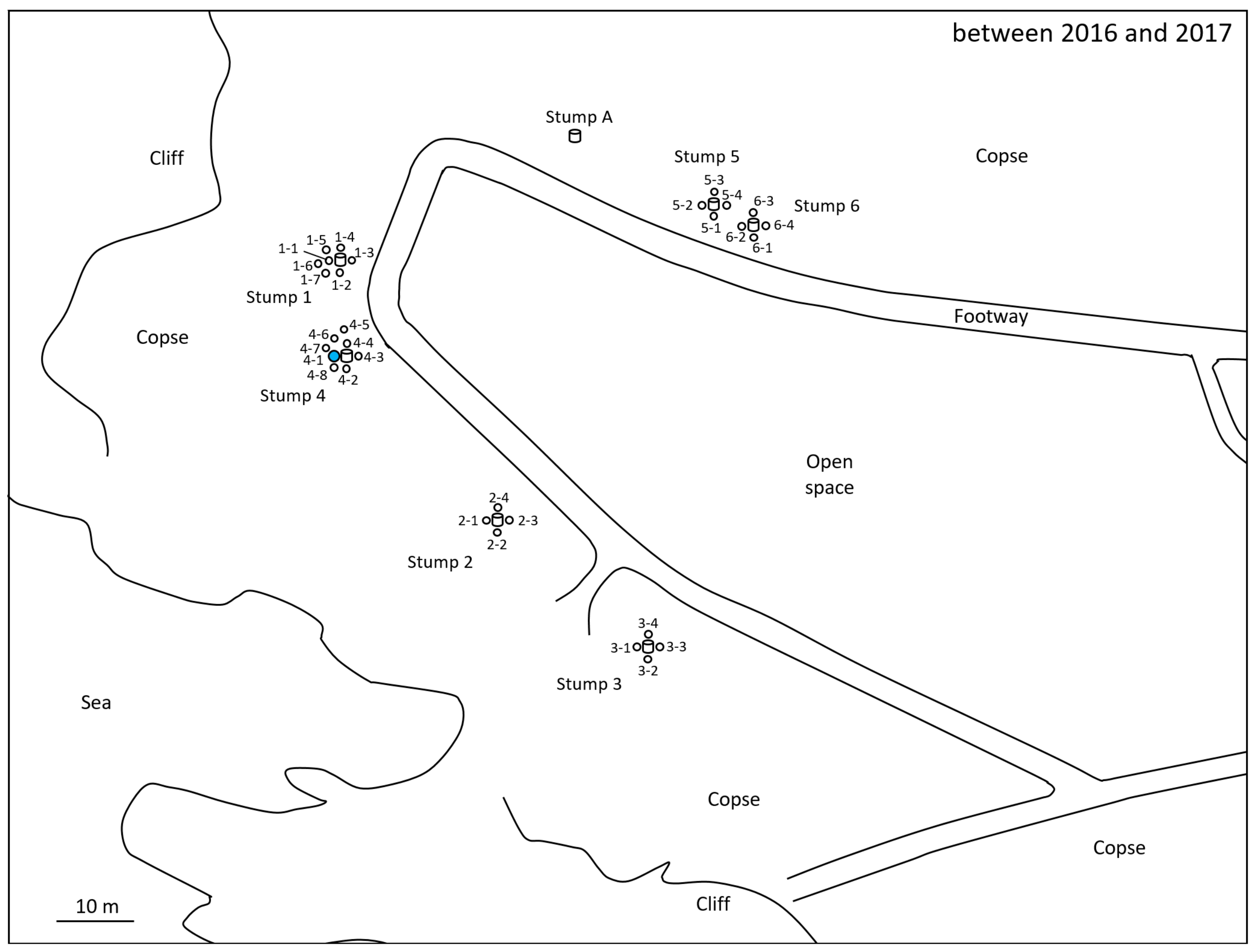
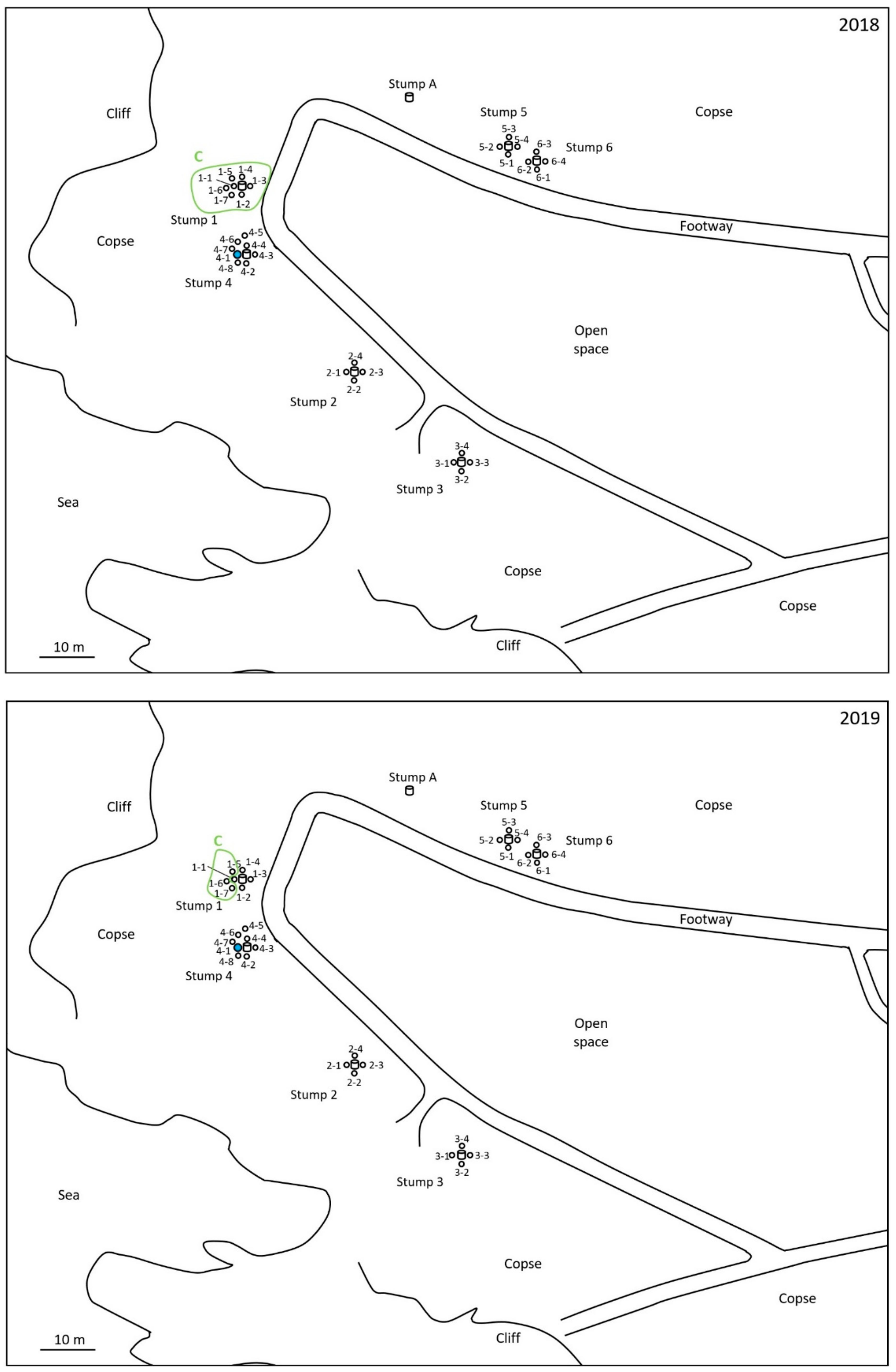
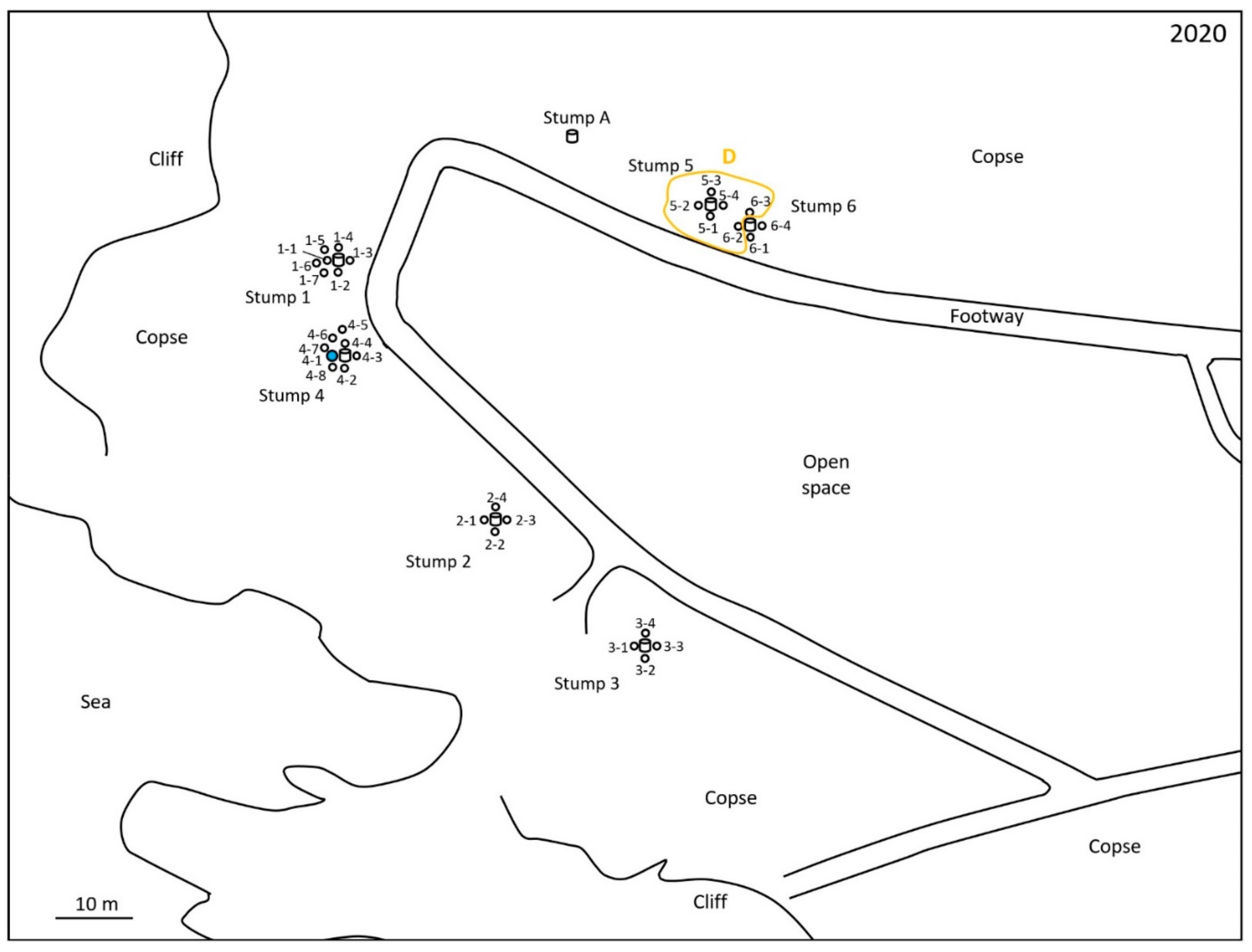
2.1. Monitoring and Collection of Termites
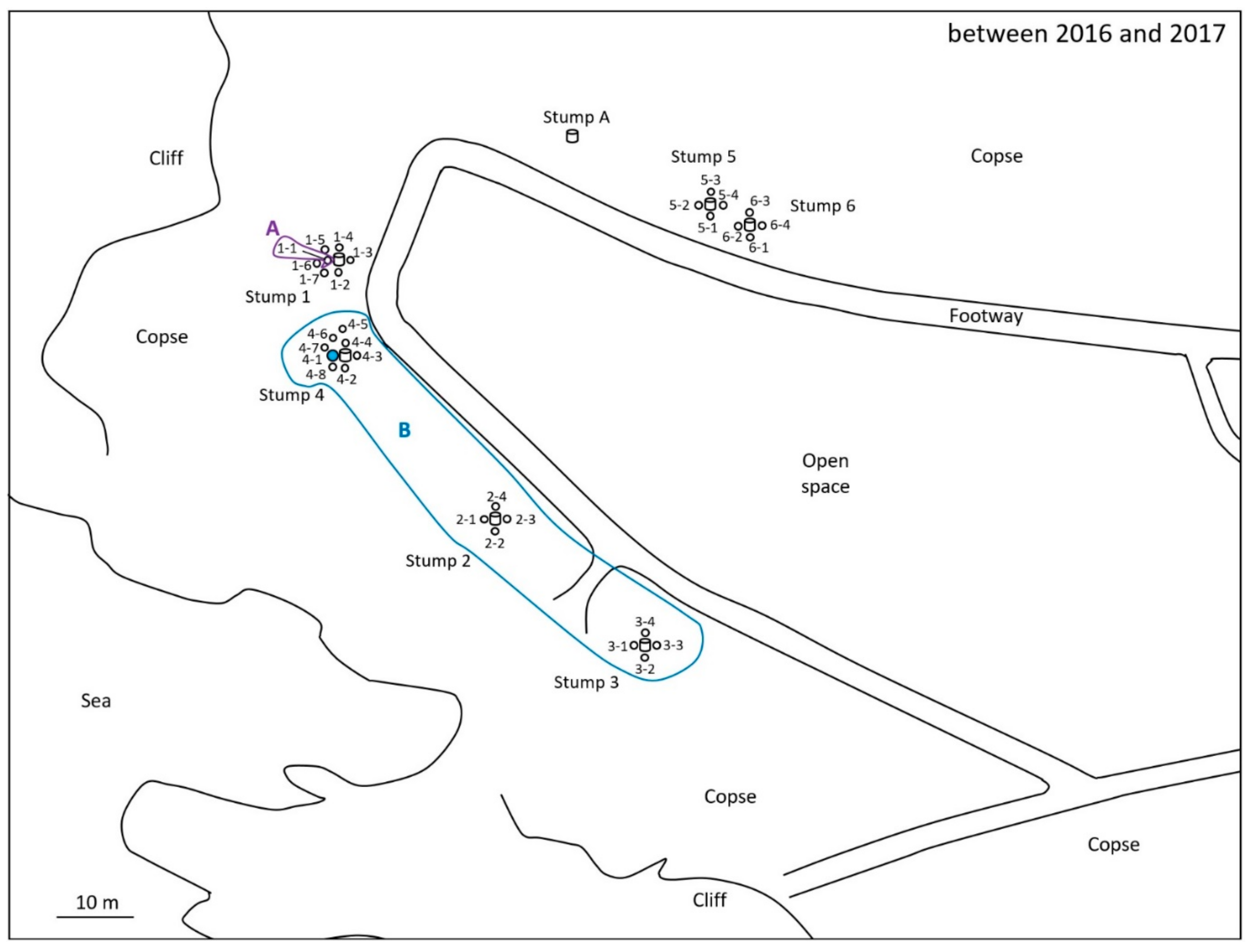
2.2. Discontinuous Soil Treatment
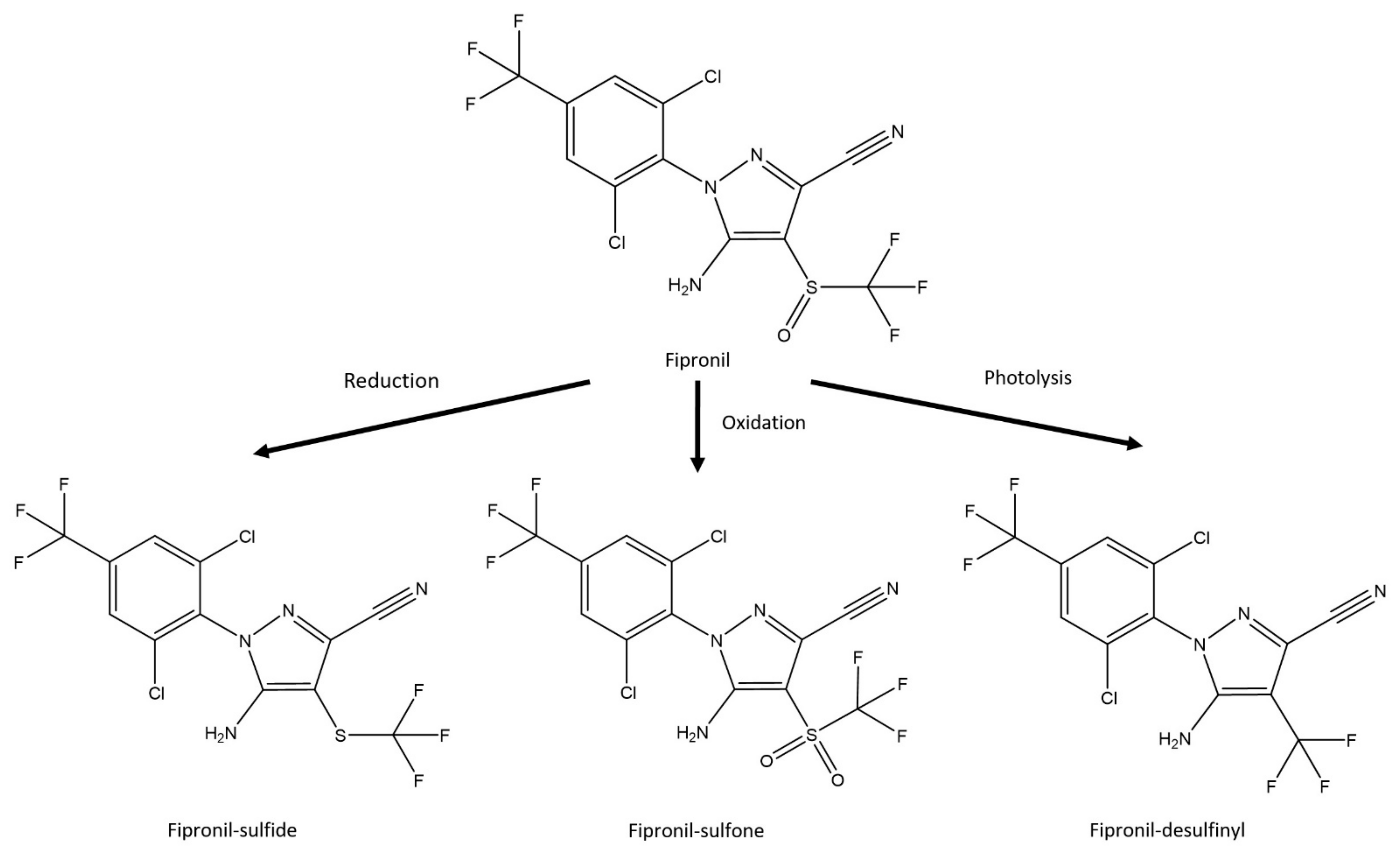
2.3. Determination of Fipronil and Its Derivatives on the Surface of Termites and in Termites
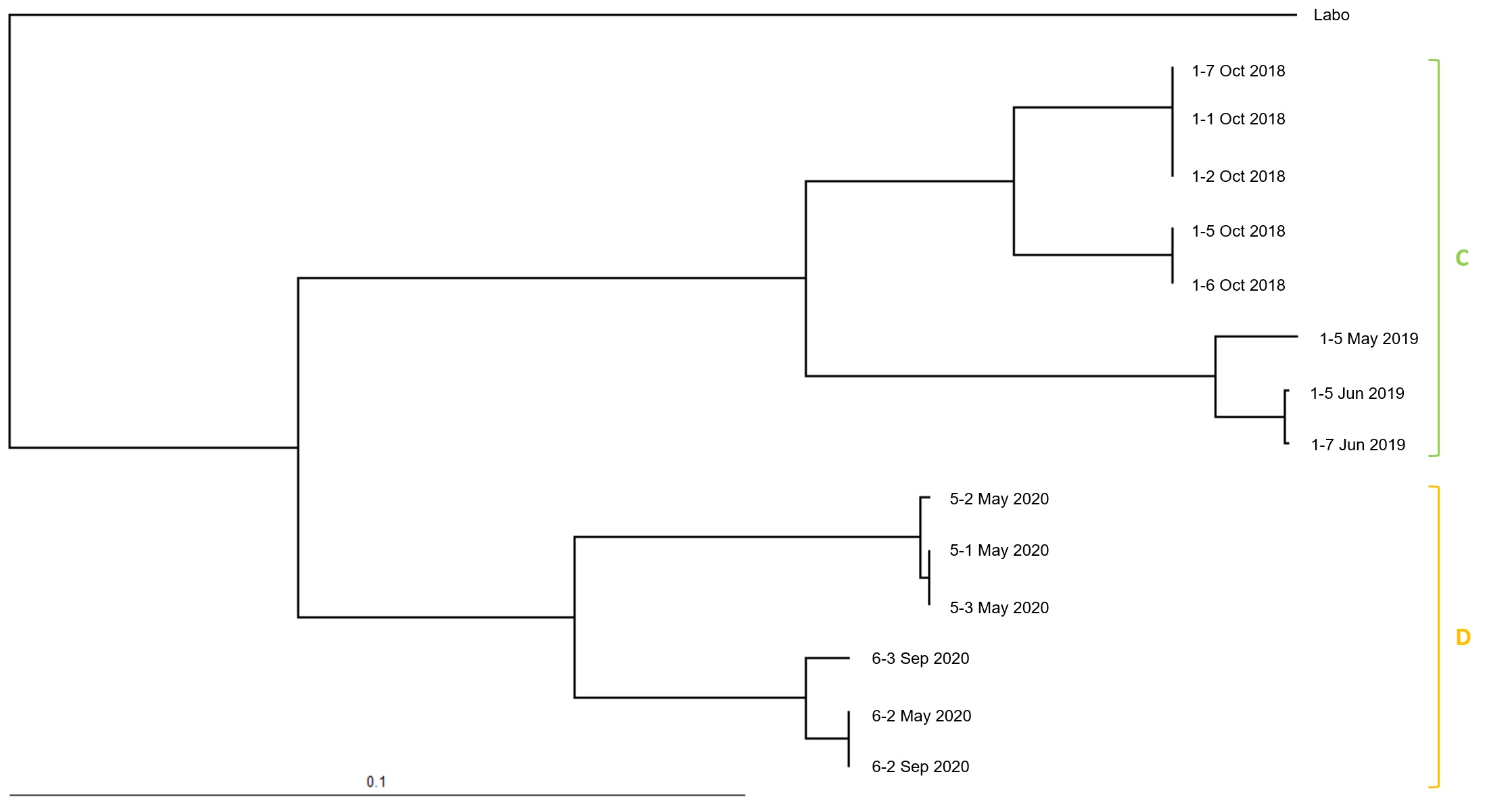
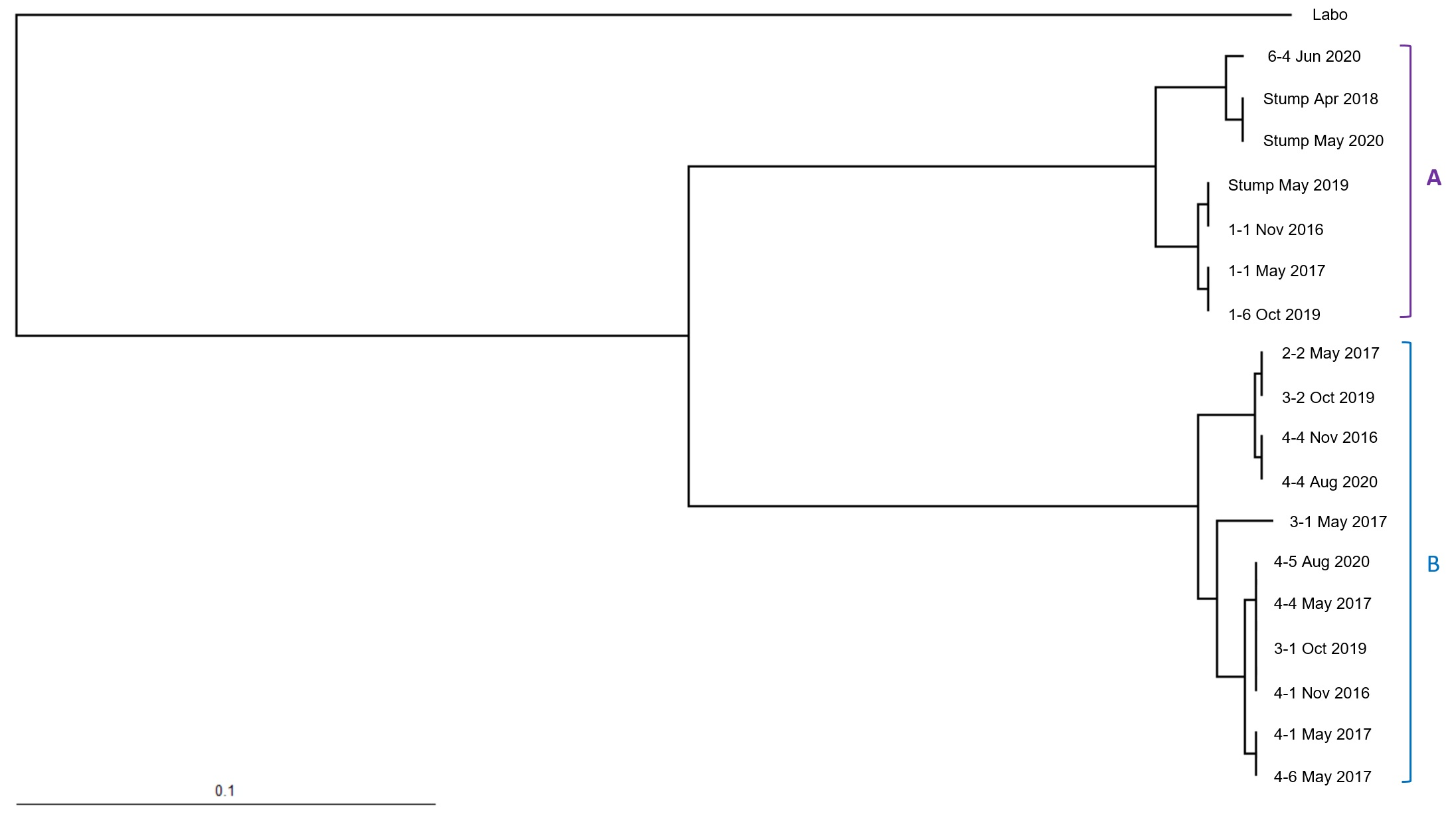
2.4. Microsatellite Genotyping for Termite Colony Affiliation
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bignell, D.E.; Eggleton, P. Termites in ecosystems. In Termites: Evolution, Sociality, Symbioses, Ecology; Abe, T., Bignell, D.E., Higashi, M., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 363–387. [Google Scholar]
- Holt, J.A.; Lepage, M. Termites and soil properties. In Termites: Evolution, Sociality, Symbioses, Ecology; Abe, T., Bignell, D.E., Higashi, M., Eds.; Springer: Dordrecht, The Netherlands, 2000; pp. 389–407. [Google Scholar]
- Jouquet, P.; Traoré, S.; Choosai, C.; Hartmann, C.; Bignell, D. Influence of termites on ecosystem functioning. Ecosystem services provided by termites. Eur. J. Soil Biol. 2011, 47, 215–222. [Google Scholar] [CrossRef]
- Rouland-Lefèvre, C. Termites as pests of agriculture. In Biology of Termites: A Modern Synthesis; Bignel, D.E., Roisin, Y., Lo, N., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 499–517. [Google Scholar]
- Rust, M.K.; Su, N.Y. Managing social insects of urban importance. Annu. Rev. Entomol. 2012, 57, 355–375. [Google Scholar] [CrossRef] [PubMed]
- Chouvenc, T.; Li, H.-F.; Austin, J.; Bordereau, C.; Bourguignon, T.; Cameron, S.; Cancello, E.; Constantino, R.; Costa-Leonardo, A.; Eggleton, P.; et al. Revisiting Coptotermes (Isoptera: Rhinotermitidae): A global taxonomic road map for species validity and distribution of an economically important subterranean termite genus. Syst. Entomol. 2016, 41, 299–306. [Google Scholar] [CrossRef]
- Hu, X.P. Liquid Termiticides: Their Role in Subterranean Termite Management; CABI: Wallingford, UK, 2011; pp. 114–132. [Google Scholar]
- Vargo, E.L.; Parman, V. Effect of fipronil on subterranean termite colonies (Isoptera: Rhinotermitidae) in the field. J. Econ. Entomol. 2012, 105, 523–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osbrink, W.; Cornelius, M.; Lax, A. Effect of imidacloprid soil treatments on occurrence of formosan subterranean termites (Isoptera: Rhinotermitidae) in independent monitors. J. Econ. Entomol. 2006, 98, 2160–2168. [Google Scholar] [CrossRef]
- Spomer, N.A.; Kamble, S.T. Temporal changes in chlorantraniliprole and indoxacarb in four midwestern soils and bioefficacy against the eastern subterranean termite (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2011, 104, 990–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neoh, K.B.; Hu, J.; Yeoh, B.H.; Lee, C.Y. Toxicity and horizontal transfer of chlorantraniliprole against the Asian subterranean termite Coptotermes gestroi (Wasmann): Effects of donor:recipient ratio, exposure duration and soil type. Pest Manag. Sci. 2012, 68, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Parman, V.; Vargo, E.L. Colony-level effects of imidacloprid in subterranean termites (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2010, 103, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.C.; Vargo, E.L.; Keefer, T.C.; Labadie, P.; Scherer, C.W.; Gallagher, N.T.; Gold, R.E. Efficacy of chlorantraniliprole in controlling structural infestations of the eastern subterranean termite in the USA. Insects 2017, 8, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, E.G., Jr.; Spink, W.T. Foraging galleries of the formosan subterranean termite, Coptotermes formosanus, 1 in Louisiana2. Ann. Entomol. Soc. Am. 1969, 62, 536–542. [Google Scholar] [CrossRef]
- Ehrhorn, E.M. The Termites of Hawaii, Their Economic Significance and Control, and the Distribution of Termites by Commerce; University of California Press: Berkeley, CA, USA, 1934; pp. 321–333. [Google Scholar]
- Ezaki, T.; Niimi, K.; Doi, T.; Momohara, I. The termite control standard specifications for new buildngs (in Japanese). In Basic Knowledge for Control Methods of Termite and Wood Decay; The Japan Termite Control Association: Tokyo, Japan, 2016; pp. 176–189. [Google Scholar]
- Vargo, E.; Husseneder, C.; Grace, J. Colony and population genetic structure of the Formosan subterranean termite, Coptotermes formosanus, in Japan. Mol. Ecol. 2003, 12, 2599–2608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Husseneder, C.; Simms, D.M.; Riegel, C. Evaluation of treatment success and patterns of reinfestation of the Formosan subterranean termite (Isoptera: Rhinotermitidae). J. Econ. Entomol. 2007, 100, 1370–1380. [Google Scholar] [CrossRef]
- Vargo, E.L.; Henderson, G. Identification of polymorphic microsatellite loci in the formosan subterranean termite Coptotermes formosanus Shiraki. Mol. Ecol. 2000, 9, 1935–1938. [Google Scholar] [CrossRef]
- Hayashi, Y.; Kitade, O.; Kojima, J.I. Microsatellite loci in the Japanese subterranean termite, Reticulitermes speratus. Mol. Ecol. Notes 2002, 2, 518–520. [Google Scholar] [CrossRef]
- Indrayani, Y.; Matsumura, K.; Yoshimura, T.; Imamura, Y.; Itakura, S. Development of microsatellite markers for the drywood termite Incisitermes minor (Hagen). Mol. Ecol. Notes 2006, 6, 1249–1251. [Google Scholar] [CrossRef]
- Weir, B.S. Estimating F-statistics: A historical view. Philos. Sci. 2012, 79, 637–643. [Google Scholar] [CrossRef] [Green Version]
- Lewis, P.O.; Zaykin, D. Genetic Data Analysis: Computer Program for the Analysis of Allelic Data; Version 1.0; North Carolina State University: Raleigh, NC, USA, 2001. [Google Scholar]
- Vargo, E.L.; Husseneder, C. Biology of subterranean termites: Insights from molecular studies of Reticulitermes and Coptotermes. Annu. Rev. Entomol. 2008, 54, 379–403. [Google Scholar] [CrossRef]
- Thorne, B.L.; Traniello, J.F.A.; Adams, E.S.; Bulmer, M. Reproductive dynamics and colony structure of subterranean termites of the genus Reticulitermes (Isoptera Rhinotermitidae): A review of the evidence from behavioral, ecological, and genetic studies. Ethol. Ecol. Evol. 1999, 11, 149–169. [Google Scholar] [CrossRef]
- Messenger, M.; Su, N.-Y.; Husseneder, C.; Grace, J. Elimination and reinvasion studies with Coptotermes formosanus (Isoptera: Rhinotermitidae) in Louisiana. J. Econ. Entomol. 2005, 98, 916–929. [Google Scholar] [CrossRef] [PubMed]
- Husseneder, C.; Powell, J.E.; Grace, J.K.; Vargo, E.L.; Matsuura, K. Worker size in the formosan subterranean termite in relation to colony breeding structure as inferred from molecular markers. Environ. Entomol. 2008, 3, 400–408. [Google Scholar] [CrossRef]
- Hayashi, Y.; Kitade, O.; Gonda, M.; Kondo, T.; Miyata, H.; Urayama, K. Diverse colony genetic structures in the Japanese subterranean termite Reticulitermes speratus (Isoptera: Rhinotermitidae). Sociobiology 2005, 46, 175–184. [Google Scholar]
- Majid, A.H.A.; Kamble, S.T.; Chen, H. Breeding patterns and population genetics of eastern Subterranean termites Reticulitermes flavipes in urban environment of Nebraska, United States. Sociobiology 2018, 65, 506–514. [Google Scholar] [CrossRef]
- Jenkins, T.M.; Basten, C.J.; Kresovich, S.; Forschler, B.T. Mitochondrial gene sequence questions Reticulitermes sp. social structure (Isoptera: Rhinotermitidae). Sociobiology 1999, 34, 161–172. [Google Scholar]
- Matsuura, K.; Fujimoto, M.; Goka, K. Sexual and asexual colony foundation and the mechanism of facultative parthenogenesis in the termite Reticulitermes speratus (Isoptera, Rhinotermitidae). Insectes Sociaux 2004, 51, 325–332. [Google Scholar] [CrossRef]
- Hayashi, Y.; Kitade, O.; Kojima, J.-i. Parthenogenetic reproduction in neotenics of the subterranean termite Reticulitermes speratus (Isoptera: Rhinotermitidae). Entomol. Sci. 2003, 6, 253–257. [Google Scholar] [CrossRef]
- Su, N.-Y. Response of the formosan subterranean termites (Isoptera: Rhinotermitidae) to baits or nonrepellent termiticides in extended foraging arenas. J. Econ. Entomol. 2006, 98, 2143–2152. [Google Scholar] [CrossRef]
- Husseneder, C.; Simms, D.M.; Delatte, J.R.; Wang, C.; Grace, J.K.; Vargo, E.L. Genetic diversity and colony breeding structure in native and introduced ranges of the Formosan subterranean termite, Coptotermes formosanus. Biol. Invasions 2012, 14, 419–437. [Google Scholar] [CrossRef] [Green Version]
- Bernard, S.; Osbrink, W.; Su, N.-Y. Response of the Formosan subterranean termite to neighboring con-specific populations after baiting with noviflumuron. J. Econ. Entomol. 2017, 110, 575–583. [Google Scholar] [CrossRef] [PubMed]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cohort 1 | Population | n | NA | FIS | FIT | FST | He | Ho | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Station No | Collection Date | Cf4-4 | Cf4-9A | Cf8-4 | Cf10-5 | |||||||
| 1-1 | Nov 2016 | A | 10 | 1 | 1 | 2 | 2 | −0.340 | −0.035 | 0.226 | 0.250 | 0.250 |
| 1-1 | May 2017 | A | 11 | 2 | 1 | 2 | 2 | −0.312 | −0.017 | 0.225 | 0.292 | 0.409 |
| 1-6 | Oct 2019 | A | 10 | 2 | 1 | 2 | 2 | −0.330 | −0.025 | 0.230 | 0.272 | 0.275 |
| 2-2 | May 2017 | B | 6 | 2 | 2 | 2 | 2 | −0.314 | −0.016 | 0.227 | 0.337 | 0.458 |
| 3-1 | May 2017 | B | 8 | 2 | 2 | 2 | 2 | −0.318 | −0.013 | 0.232 | 0.388 | 0.500 |
| 3-1 | Oct 2019 | B | 10 | 2 | 2 | 1 | 2 | −0.315 | −0.014 | 0.229 | 0.324 | 0.450 |
| 3-2 | Oct 2019 | B | 10 | 2 | 2 | 1 | 2 | −0.317 | −0.017 | 0.228 | 0.286 | 0.375 |
| 4-1 | Nov 2016 | B | 12 | 2 | 2 | 1 | 2 | −0.314 | −0.012 | 0.229 | 0.322 | 0.438 |
| 4-1 | May 2017 | B | 10 | 2 | 2 | 1 | 2 | −0.306 | −0.012 | 0.224 | 0.339 | 0.500 |
| 4-4 | Nov 2016 | B | 16 | 2 | 2 | 1 | 2 | −0.329 | −0.026 | 0.228 | 0.201 | 0.234 |
| 4-4 | May 2017 | B | 3 | 2 | 2 | 1 | 2 | −0.316 | −0.020 | 0.225 | 0.367 | 0.500 |
| 4-4 | Aug 2020 | B | 6 | 2 | 2 | 1 | 2 | −0.322 | −0.021 | 0.228 | 0.280 | 0.333 |
| 4-5 | Aug 2020 | B | 4 | 2 | 2 | 1 | 2 | −0.322 | −0.022 | 0.227 | 0.277 | 0.313 |
| 4-6 | May 2017 | B | 6 | 2 | 2 | 1 | 2 | −0.312 | −0.018 | 0.224 | 0.314 | 0.458 |
| 6-4 | Jun 2020 | A | 8 | 2 | 1 | 2 | 2 | −0.322 | −0.025 | 0.226 | 0.371 | 0.438 |
| Stump A | Apr 2018 | A | 17 | 2 | 1 | 2 | 2 | −0.328 | −0.024 | 0.230 | 0.313 | 0.368 |
| Mar 2019 | A | 8 | 1 | 1 | 2 | 2 | −0.311 | −0.018 | 0.225 | 0.246 | 0.313 | |
| May 2020 | A | 8 | 2 | 1 | 2 | 2 | −0.309 | −0.017 | 0.223 | 0.338 | 0.500 | |
| Laboratory | 10 | 2 | 2 | 2 | 2 | −0.309 | −0.103 | 0.158 | 0.397 | 0.550 | ||
| Mean ± SD (without laboratory) | 9.1 ± 3.6 | 1.9 ± 0.3 | 1.6 ± 0.5 | 1.5 ± 0.5 | 2.0 ± 0 | −0.319 ± 0.009 | −0.020 ± 0.006 | 0.227 ± 0.002 | 0.306 ± 0.048 | 0.395 ± 0.090 | ||
| Cohort 1 | Population | n | NA | FIS | FIT | FST | He | Ho | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Station No | Collection Date | Rs02 | Rs03 | Rs05 | Rs07 | |||||||
| 1-1 | Oct 2018 | C | 8 | 2 | 1 | 2 | 1 | −0.054 | 0.204 | 0.233 | 0.248 | 0.406 |
| 1-2 | Oct 2018 | C | 8 | 2 | 1 | 2 | 2 | −0.058 | 0.202 | 0.243 | 0.292 | 0.375 |
| 1-5 | Oct 2018 | C | 7 | 2 | 1 | 2 | 2 | −0.061 | 0.198 | 0.228 | 0.332 | 0.321 |
| 1-5 | Mar 2019 | C | 8 | 2 | 2 | 2 | 1 | −0.025 | 0.221 | 0.231 | 0.367 | 0.625 |
| 1-5 | Jun 2019 | C | 11 | 2 | 2 | 2 | 1 | −0.053 | 0.225 | 0.246 | 0.348 | 0.545 |
| 1-6 | Oct 2018 | C | 8 | 2 | 1 | 2 | 2 | −0.060 | 0.200 | 0.213 | 0.348 | 0.344 |
| 1-7 | Oct 2018 | C | 8 | 2 | 1 | 2 | 2 | −0.060 | 0.200 | 0.242 | 0.273 | 0.344 |
| 1-7 | Jun 2019 | C | 7 | 2 | 2 | 2 | 2 | −0.049 | 0.215 | 0.246 | 0.442 | 0.607 |
| 5-1 | May 2020 | D | 8 | 3 | 2 | 1 | 1 | −0.078 | 0.185 | 0.232 | 0.208 | 0.219 |
| 5-2 | May 2020 | D | 9 | 3 | 1 | 1 | 1 | −0.062 | 0.184 | 0.223 | 0.165 | 0.250 |
| 5-3 | May 2020 | D | 8 | 3 | 1 | 1 | 1 | −0.067 | 0.183 | 0.225 | 0.169 | 0.219 |
| 6-2 | May 2020 | D | 8 | 3 | 1 | 1 | 1 | −0.086 | 0.166 | 0.224 | 0.158 | 0.063 |
| 6-2 | Sep 2020 | D | 8 | 3 | 1 | 1 | 1 | −0.073 | 0.176 | 0.224 | 0.177 | 0.188 |
| 6-3 | Sep 2020 | D | 8 | 3 | 1 | 1 | 1 | −0.067 | 0.179 | 0.223 | 0.127 | 0.156 |
| Laboratory | 12 | 3 | 4 | 3 | 2 | −0.109 | 0.110 | 0.203 | 0.569 | 0.646 | ||
| Mean ± SD (without laboratory) | 8.1 ± 0.9 | 2.4 ± 0.5 | 1.3 ± 0.5 | 1.6 ± 0.5 | 1.4 ± 0.5 | −0.061 ± 0.014 | 0.196 ± 0.017 | 0.231 ± 0.012 | 0.261 ± 0.097 | 0.333 ± 0.169 | ||
| Cohort 1 | Population | n | NA | FIS | FIT | FST | He | Ho | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Station No | Collection Date | Rs02 | Rs03 | Rs05 | Rs07 | |||||||
| 1 | Oct 2015 | D | 9 | 2 | 3 | 1 | 1 | −0.138 | 0.213 | 0.312 | 0.301 | 0.444 |
| 1 | Nov 2018 | B | 8 | 1 | 2 | 1 | 1 | −0.146 | 0.211 | 0.313 | 0.100 | 0.125 |
| 1 | Dec 2018 | B | 8 | 1 | 2 | 1 | 1 | −0.146 | 0.211 | 0.314 | 0.131 | 0.156 |
| 1 | May 2019 | B | 12 | 1 | 2 | 1 | 1 | −0.146 | 0.207 | 0.310 | 0.040 | 0.042 |
| 1 | Nov 2019 | B | 11 | 3 | 2 | 1 | 1 | −0.142 | 0.215 | 0.316 | 0.205 | 0.250 |
| 2 | Dec 2014 | C | 12 | 3 | 3 | 1 | 2 | −0.112 | 0.229 | 0.312 | 0.438 | 0.458 |
| 2 | Nov 2019 | B | 12 | 2 | 2 | 1 | 1 | −0.145 | 0.211 | 0.313 | 0.119 | 0.146 |
| 3 | Nov 2018 | B | 8 | 1 | 2 | 1 | 1 | −0.144 | 0.213 | 0.313 | 0.125 | 0.188 |
| 3 | Dec 2018 | B | 8 | 1 | 2 | 1 | 1 | −0.146 | 0.211 | 0.314 | 0.131 | 0.156 |
| 3 | May 2019 | B | 12 | 2 | 2 | 1 | 1 | −0.145 | 0.211 | 0.313 | 0.126 | 0.146 |
| 4 | Oct 2015 | A | 14 | 3 | 3 | 2 | 1 | −0.154 | 0.211 | 0.320 | 0.307 | 0.321 |
| 5 | Dec 2014 | D | 12 | 2 | 3 | 1 | 1 | −0.146 | 0.207 | 0.312 | 0.257 | 0.271 |
| 5 | Oct 2015 | D | 17 | 2 | 3 | 1 | 1 | −0.138 | 0.210 | 0.309 | 0.291 | 0.382 |
| 5 | Nov 2018 | B | 8 | 1 | 2 | 1 | 1 | −0.144 | 0.213 | 0.313 | 0.125 | 0.188 |
| 6 | Oct 2015 | D | 19 | 2 | 3 | 1 | 1 | −0.143 | 0.197 | 0.299 | 0.209 | 0.250 |
| 6 | Nov 2019 | B | 10 | 2 | 2 | 1 | 1 | −0.142 | 0.213 | 0.314 | 0.158 | 0.200 |
| 7 | Oct 2015 | A | 4 | 3 | 3 | 3 | 2 | −0.162 | 0.205 | 0.317 | 0.580 | 0.438 |
| 7 | Nov 2017 | B | 12 | 2 | 2 | 1 | 1 | −0.144 | 0.210 | 0.312 | 0.093 | 0.104 |
| 8 | Nov 2019 | E | 8 | 2 | 2 | 2 | 1 | −0.142 | 0.213 | 0.312 | 0.273 | 0.344 |
| 9 | May 2016 | B | 12 | 2 | 2 | 1 | 1 | −0.146 | 0.210 | 0.312 | 0.093 | 0.104 |
| 11 | May 2019 | A | 11 | 3 | 2 | 2 | 1 | −0.137 | 0.222 | 0.318 | 0.315 | 0.386 |
| 12 | May 2016 | C | 12 | 2 | 2 | 2 | 2 | −0.145 | 0.198 | 0.302 | 0.414 | 0.396 |
| 13 | Dec 2014 | C | 6 | 2 | 3 | 1 | 2 | −0.130 | 0.219 | 0.313 | 0.345 | 0.375 |
| 13 | Oct 2015 | C | 18 | 4 | 2 | 1 | 2 | −0.194 | 0.197 | 0.324 | 0.251 | 0.250 |
| 14 | May 2019 | A | 12 | 3 | 2 | 2 | 1 | −0.144 | 0.221 | 0.322 | 0.369 | 0.396 |
| 15 | Dec 2018 | A | 8 | 3 | 2 | 2 | 1 | −0.141 | 0.218 | 0.317 | 0.390 | 0.438 |
| 16 | Sept 2013 | A | 14 | 2 | 3 | 1 | 1 | −0.150 | 0.209 | 0.315 | 0.299 | 0.286 |
| 16 | Dec 2018 | A | 8 | 3 | 2 | 2 | 1 | −0.147 | 0.215 | 0.318 | 0.290 | 0.313 |
| 16 | May 2019 | A | 11 | 3 | 2 | 2 | 1 | −0.150 | 0.216 | 0.321 | 0.328 | 0.295 |
| 18 | Aug 2018 | A | 11 | 3 | 2 | 2 | 1 | −0.144 | 0.219 | 0.320 | 0.341 | 0.364 |
| 18 | Dec 2018 | - | 8 | 3 | 2 | 1 | 2 | −0.130 | 0.224 | 0.316 | 0.433 | 0.563 |
| 18 | Nov 2019 | E | 11 | 3 | 1 | 1 | 1 | −0.143 | 0.206 | 0.307 | 0.124 | 0.136 |
| 20 | Aug 2018 | E | 10 | 3 | 1 | 1 | 2 | −0.162 | 0.202 | 0.315 | 0.253 | 0.175 |
| 27 | Aug 2018 | A | 11 | 3 | 2 | 2 | 1 | −0.134 | 0.225 | 0.318 | 0.331 | 0.523 |
| Laboratory | 12 | 3 | 4 | 3 | 2 | −0.151 | 0.193 | 0.299 | 0.569 | 0.646 | ||
| Mean ± SD (without laboratory) | 10.8 ± 3.2 | 2.3 ± 0.8 | 2.2 ± 0.5 | 1.4 ± 0.5 | 1.2 ± 0.4 | −0.145 ± 0.012 | 0.212 ± 0.008 | 0.314 ± 0.005 | 0.252 ± 0.126 | 0.283 ± 0.133 | ||
| Station No./Caste | n | Sample | Fipronil | Fipronil-Sulfide (Reductive Metabolite) | Fipronil-Sulfone (Oxidative Metabolite) | Fipronil-Desulfinyl (Photodegradation Product) | Total |
|---|---|---|---|---|---|---|---|
| (pg/termite) 2 | |||||||
| 2-2/Worker | 14 (52.3 mg) | Water fraction | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| (0.5 ppb, 1.8 pg/termite) 1 | Acetonitrile fraction | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |
| Termite body fraction | 1.9 | 0.0 | 4.5 | 0.0 | 6.4 | ||
| Total | 1.9 | 0.0 | 4.5 | 0.0 | 6.4 | ||
| 2-2/Soldier | 2 (8.4 mg) | Water fraction | 0 | 0 | 0 | 0 | 0 |
| (3 ppb, 12 pg/termite) 1 | Acetonitrile fraction | 0 | 0 | 0 | 0 | 0 | |
| Termite body fraction | 0 | 0 | 0 | 0 | 0 | ||
| Total | 0 | 0 | 0 | 0 | 0 | ||
| 4-4/Soldier | 1 (3.4 mg) | Water fraction | 21 | 0 | 23 | 0 | 44 |
| (6 ppb, 21 pg/termite) 1 | Acetonitrile fraction | 32 | 0 | 38 | 162 | 231 | |
| Termite body fraction | 0 | 0 | 0 | 0 | 0 | ||
| Total | 53 | 0 | 60 | 162 | 275 | ||
| Station No. | n | Sample | Fipronil | Fipronil-Sulfide (Reductive Metabolite) | Fipronil-Sulfone (Oxidative Metabolite) | Fipronil-Desulfinyl (Photodegradation Product) | Total |
|---|---|---|---|---|---|---|---|
| (pg/termite) 2 | |||||||
| 1 | 163 (268.3 mg) | Water fraction | 0.0 | 0.0 | 1.6 | 0.0 | 1.6 |
| (0.5 ppb, 0.8 pg/termite) 1 | Acetonitrile fraction | 0.0 | 0.0 | 1.5 | 0.0 | 1.5 | |
| Termite body fraction | 1.8 | 0.0 | 12.6 | 0.0 | 14.4 | ||
| Total | 1.8 | 0.0 | 15.8 | 0.0 | 17.6 | ||
| 4 | 13 (26.8 mg) | Water fraction | 0 | 0 | 0 | 0 | 0 |
| (2 ppb, 4 pg/termite) 1 | Acetonitrile fraction | 0 | 0 | 0 | 0 | 0 | |
| Termite body fraction | 0 | 0 | 0 | 0 | 0 | ||
| Total | 0 | 0 | 0 | 0 | 0 | ||
| 5 | 59 (124.1 mg) | Water fraction | 0.0 | 0.0 | 3.3 | 0.0 | 3.3 |
| (0.5 ppb, 1.1 pg/termite) 1 | Acetonitrile fraction | 0.0 | 0.0 | 3.1 | 0.0 | 3.1 | |
| Termite body fraction | 24.8 | 0.0 | 90.0 | 0.0 | 114.8 | ||
| Total | 24.8 | 0.0 | 96.4 | 0.0 | 121.2 | ||
| 6 | 48 (84.2 mg) | Water fraction | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| (0.6 ppb, 1.1 pg/termite) 1 | Acetonitrile fraction | 0.0 | 0.0 | 1.3 | 0.0 | 1.3 | |
| Termite body fraction | 1.4 | 0.0 | 9.7 | 0.0 | 11.1 | ||
| Total | 1.4 | 0.0 | 11.0 | 0.0 | 12.4 | ||
| Station No. | n | Sample | Fipronil | Fipronil-Sulfide (Reductive Metabolite) | Fipronil-Sulfone (Oxidative Metabolite) | Fipronil-Desulfinyl (Photodegradation Product) | Total |
|---|---|---|---|---|---|---|---|
| (pg/termite) 2 | |||||||
| 1 | 135 (294 mg) | Water fraction | 0.0 | 0.0 | 2.2 | 0.0 | 2.2 |
| (0.5 ppb, 1.1 pg/termite) 1 | Acetonitrile fraction | 0.0 | 0.0 | 3.2 | 0.0 | 3.2 | |
| Termite body fraction | 0.0 | 0.0 | 18.0 | 0.0 | 18.0 | ||
| Total | 0.0 | 0.0 | 23.4 | 0.0 | 23.4 | ||
| 5 | 5 (9.3 mg) | Water fraction | 0 | 0 | 0 | 0 | 0 |
| (5 ppb, 9.3 pg/termite) 1 | Acetonitrile fraction | 0 | 0 | 0 | 0 | 0 | |
| Termite body fraction | 0 | 0 | 30 | 0 | 30 | ||
| Total | 0 | 0 | 30 | 0 | 30 | ||
| 6 | 71 (117.6 mg) | Water fraction | 0.0 | 0.0 | 1.6 | 0.0 | 1.6 |
| (0.5 ppb, 0.8 pg/termite) 1 | Acetonitrile fraction | 0.0 | 0.0 | 2.1 | 0.0 | 2.1 | |
| Termite body fraction | 0.0 | 0.0 | 4.2 | 0.0 | 4.2 | ||
| Total | 0.0 | 0.0 | 7.9 | 0.0 | 7.9 | ||
| Compound | Parent Ion (m/z) | Daughter Ion (m/z) | Collision Energy (eV) | S-Lens |
|---|---|---|---|---|
| Fipronil | 435 | 250 | 32 | 96 |
| Fipronil-Sulfide | 419 | 262 | 33 | 87 |
| Fipronil-Sulfone | 451 | 282 | 31 | 104 |
| Fipronil-Desulfinyl | 387 | 351 | 24 | 77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Itakura, S.; Ohdake, J.; Takino, T.; Umezawa, K. Colony Suppression and Possible Colony Elimination of the Subterranean Termites Coptotermes formosanus and Reticulitermes speratus by Discontinuous Soil Treatment Using a Diluent of Fipronil Suspension Concentrate. Insects 2021, 12, 334. https://doi.org/10.3390/insects12040334
Itakura S, Ohdake J, Takino T, Umezawa K. Colony Suppression and Possible Colony Elimination of the Subterranean Termites Coptotermes formosanus and Reticulitermes speratus by Discontinuous Soil Treatment Using a Diluent of Fipronil Suspension Concentrate. Insects. 2021; 12(4):334. https://doi.org/10.3390/insects12040334
Chicago/Turabian StyleItakura, Shuji, Johji Ohdake, Takashi Takino, and Kiwamu Umezawa. 2021. "Colony Suppression and Possible Colony Elimination of the Subterranean Termites Coptotermes formosanus and Reticulitermes speratus by Discontinuous Soil Treatment Using a Diluent of Fipronil Suspension Concentrate" Insects 12, no. 4: 334. https://doi.org/10.3390/insects12040334
APA StyleItakura, S., Ohdake, J., Takino, T., & Umezawa, K. (2021). Colony Suppression and Possible Colony Elimination of the Subterranean Termites Coptotermes formosanus and Reticulitermes speratus by Discontinuous Soil Treatment Using a Diluent of Fipronil Suspension Concentrate. Insects, 12(4), 334. https://doi.org/10.3390/insects12040334