
Advances in the Study of Olfaction in Eusocial Ants


{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
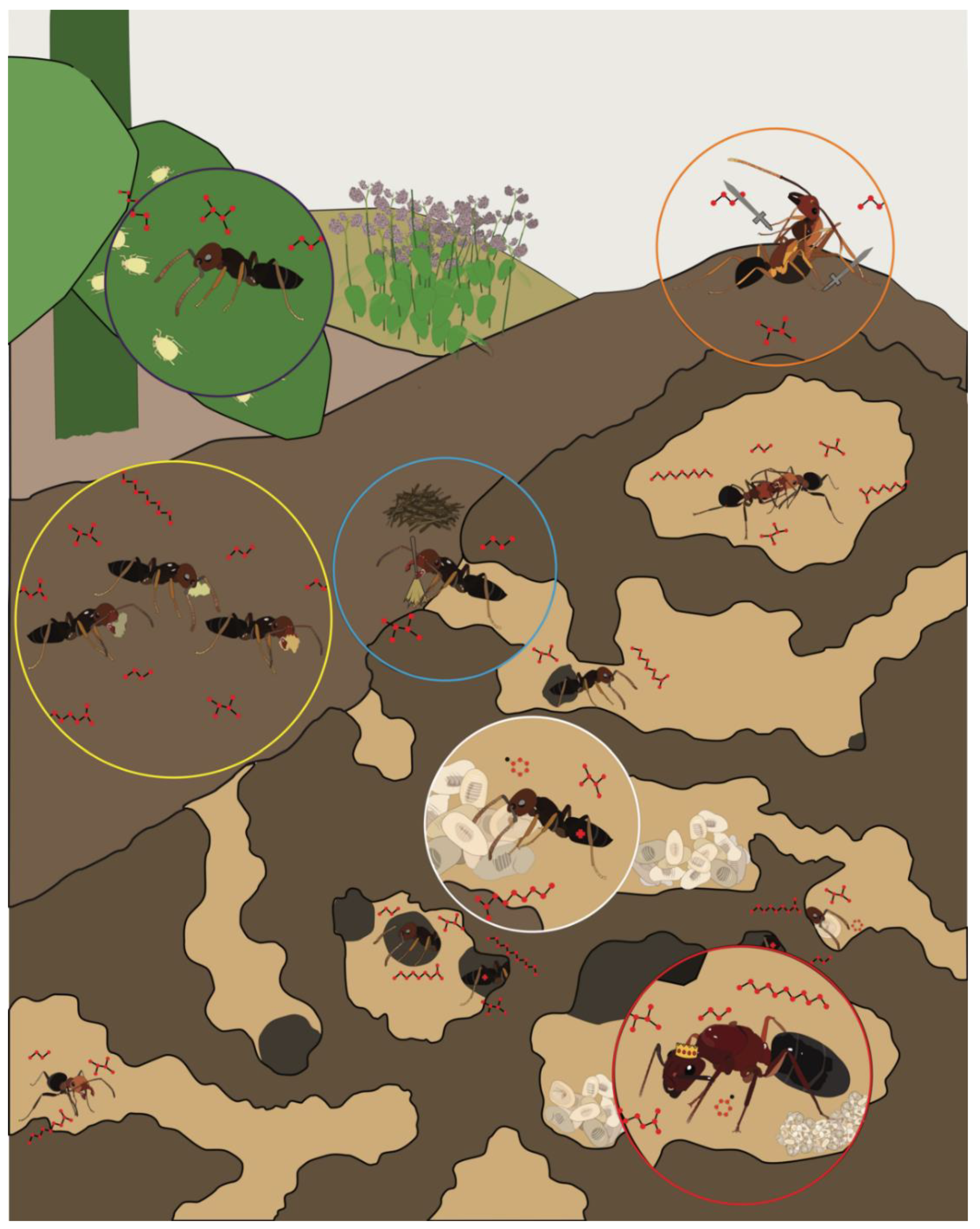
1.1. A Remarkable Olfactory Sense
1.2. Aim and Scope of This Review
2. The Peripheral Olfactory System
2.1. Untangling Odor Coding in the Peripheral Sensilla
2.2. Identifying Odor Ligands through the Deorphanization of Chemoreceptors
3. Central Olfactory System
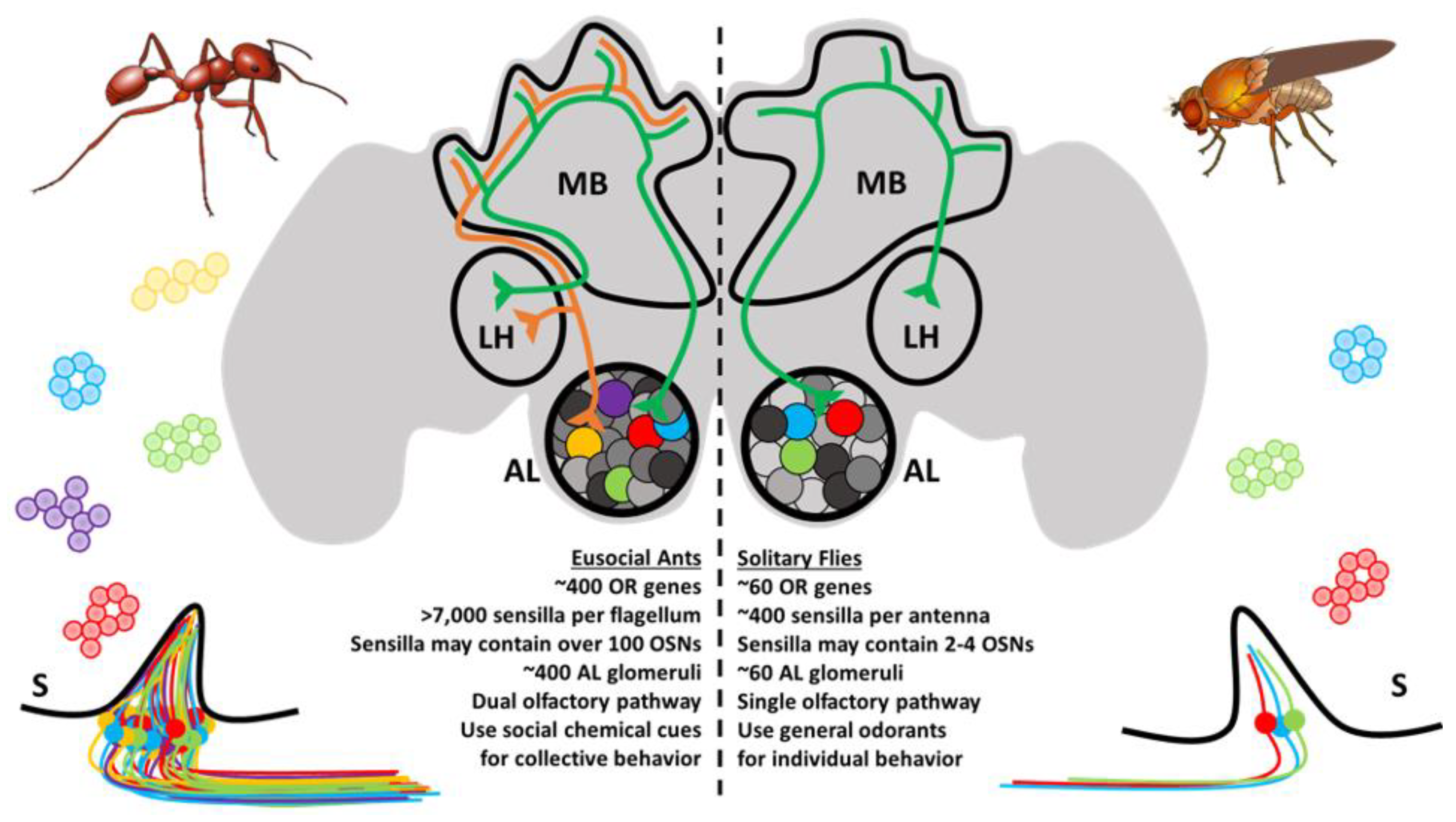
3.1. An Overview of the Central Olfactory System in Insects
3.2. Structure and Function of the Antennal Lobe
3.3. Olfactory Sensory Neurons and the Ontogeny of the Antennal Lobe
4. Genomics, Evolution, and the Regulation of Chemosensory Genes
4.1. Targeted Gene Editing in Formicidae
4.2. The Technical Challenges of Gene Editing in Eusocial Hymenoptera
4.3. Innovative Variations and Alternatives to Gene Editing
4.4. Advances in Epigenetic Engineering
4.5. Artificially Induced Histone Modifications Dramatically Alter Ant Behavior
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hölldobler, B.; Wilson, E.O. The Ants; Belknap Press of Harvard University Press: Cambridge, MA, USA, 1990; p. 732. [Google Scholar]
- Weber, N.A. Gardening Ants, the Attines; American Philosophical Society: Philadelphia, PA, USA, 1972; p. 146. [Google Scholar]
- Kronauer, D.J.C. Army Ants: Nature’s Ultimate Social Hunters; Harvard University Press: Cambridge, MA, USA, 2020; p. 384. [Google Scholar]
- Pratt, S.C.; Mallon, E.B.; Sumpter, D.J.; Franks, N.R. Quorum sensing, recruitment, and collective decision-making during colony emigration by the ant Leptothorax albipennis. Behav. Ecol. Sociobiol. 2002, 52, 117–127. [Google Scholar] [CrossRef]
- Ferguson, S.T.; Anand, A.; Zwiebel, J. Olfactory genomics of eusociality within the Hymenoptera. In Insect Pheromone Biochemistry and Molecular Biology, 2nd ed.; Blomquist, G.J., Vogt, R.G., Eds.; Elsevier: Cambridge, MA, USA, 2020; pp. 507–546. [Google Scholar]
- Sprenger, P.P.; Menzel, F. Cuticular hydrocarbons in ants (Hymenoptera: Formicidae) and other insects: How and why they differ among individuals, colonies, and species. Myrmecol. News 2020, 30, 1–26. [Google Scholar] [CrossRef]
- Greene, M.J.; Pinter-Wollman, N.; Gordon, D.M. Interactions with combined chemical cues inform harvester ant foragers’ decisions to leave the nest in search of food. PLoS ONE 2013, 8, e52219. [Google Scholar] [CrossRef] [Green Version]
- Morel, L.; Vandermeer, R.K.; Lavine, B.K. Ontogeny of Nestmate Recognition Cues in the Red Carpenter Ant (Camponotus-Floridanus)—Behavioral and Chemical Evidence for the Role of Age and Social Experience. Behav. Ecol. Sociobiol. 1988, 22, 175–183. [Google Scholar] [CrossRef]
- Arrese, E.L.; Soulages, J.L. Insect fat body: Energy, metabolism, and regulation. Annu. Rev. Entomol. 2010, 55, 207–225. [Google Scholar] [CrossRef] [Green Version]
- Roma, G.C.; Bueno, O.C.; Camargo-Mathias, M.I. Morpho-physiological analysis of the insect fat body: A review. Micron 2010, 41, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Zurek, L.; Dykstra, M.J.; Schal, C. Hydrocarbon synthesis by enzymatically dissociated oenocytes of the abdominal integument of the German Cockroach, Blattella germanica. Naturwissenschaften 2003, 90, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Soroker, V.; Hefetz, A. Hydrocarbon site of synthesis and circulation in the desert ant Cataglyphis niger. J. Insect Physiol. 2000, 46, 1097–1102. [Google Scholar] [CrossRef]
- Bagnères, A.G.; Morgan, E.D. The postpharyngeal glands and the cuticle of Formicidae contain the same characteristic hydrocarbons. Experientia 1991, 47, 106–111. [Google Scholar] [CrossRef]
- Lenz, E.L.; Krasnec, M.O.; Breed, M.D. Identification of Undecane as an Alarm Pheromone of the Ant Formica argentea. J. Insect Behav. 2013, 26, 101–108. [Google Scholar] [CrossRef]
- Löfqvist, J. Formic acid and saturated hydrocarbons as alarm pheromones for the ant Formica rufa. J. Insect Physiol. 1976, 22, 1331–1346. [Google Scholar] [CrossRef]
- Fujiwara-Tsujii, N.; Yamagata, N.; Takeda, T.; Mizunami, M.; Yamaoka, R. Behavioral responses to the alarm pheromone of the ant Camponotus obscuripes (Hymenoptera: Formicidae). Zool. Sci. 2006, 23, 353–358. [Google Scholar] [CrossRef]
- Fox, E. Venom Toxins of Fire Ants. In Venom Genomics and Proteomics; Gopalakrishnakone, P., Calvete, J.J., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 149–167. [Google Scholar]
- Hashmi, A.A. A revision of the Neotropical ant subgenus Myrmothrix of genus Camponotus. Studia Entomol. 1973, 16, 1–140. [Google Scholar] [CrossRef]
- Haak, U.; Holldobler, B.; Bestmann, H.J.; Kern, F. Species-specificity in trail pheromones and Dufour’s gland contents of Camponotus atriceps and C. floridanus (Hymenoptera: Formicidae). Chemoecology 1996, 7, 85–93. [Google Scholar] [CrossRef]
- Bonasio, R.; Zhang, G.; Ye, C.; Mutti, N.S.; Fang, X.; Qin, N.; Donahue, G.; Yang, P.; Li, Q.; Li, C.; et al. Genomic comparison of the ants Camponotus floridanus and Harpegnathos saltator. Science 2010, 329, 1068–1071. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Slone, J.D.; Rokas, A.; Berger, S.L.; Liebig, J.; Ray, A.; Reinberg, D.; Zwiebel, L.J. Phylogenetic and transcriptomic analysis of chemosensory receptors in a pair of divergent ant species reveals sex-specific signatures of odor coding. PLoS Genet. 2012, 8, e1002930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, X.; Rokas, A.; Berger, S.L.; Liebig, J.; Ray, A.; Zwiebel, L.J. Chemoreceptor Evolution in Hymenoptera and Its Implications for the Evolution of Eusociality. Genome Biol. Evol. 2015, 7, 2407–2416. [Google Scholar] [CrossRef] [Green Version]
- Vosshall, L.B.; Hansson, B.S. A unified nomenclature system for the insect olfactory coreceptor. Chem. Senses 2011, 36, 497–498. [Google Scholar] [CrossRef] [Green Version]
- Slone, J.D.; Pask, G.M.; Ferguson, S.T.; Millar, J.G.; Berger, S.L.; Reinberg, D.; Liebig, J.; Ray, A.; Zwiebel, L.J. Functional characterization of odorant receptors in the ponerine ant, Harpegnathos saltator. Proc. Natl. Acad. Sci. USA 2017, 114, 8586–8591. [Google Scholar] [CrossRef] [Green Version]
- Pask, G.M.; Slone, J.D.; Millar, J.G.; Das, P.; Moreira, J.A.; Zhou, X.; Bello, J.; Berger, S.L.; Bonasio, R.; Desplan, C.; et al. Specialized odorant receptors in social insects that detect cuticular hydrocarbon cues and candidate pheromones. Nat. Commun. 2017, 8, 297. [Google Scholar] [CrossRef]
- Clyne, P.J.; Warr, C.G.; Carlson, J.R. Candidate taste receptors in Drosophila. Science 2000, 287, 1830–1834. [Google Scholar] [CrossRef] [PubMed]
- Kremer, J.M.M.; Nooten, S.S.; Cook, J.M.; Ryalls, J.M.W.; Barton, C.V.M.; Johnson, S.N. Elevated atmospheric carbon dioxide concentrations promote ant tending of aphids. J. Anim. Ecol. 2018, 87, 1475–1483. [Google Scholar] [CrossRef] [PubMed]
- Römer, D.; Bollazzi, M.; Roces, F. Carbon dioxide sensing in an obligate insect-fungus symbiosis: CO2 preferences of leaf-cutting ants to rear their mutualistic fungus. PLoS ONE 2017, 12, e0174597. [Google Scholar] [CrossRef] [PubMed]
- Kleineidam, C.; Tautz, J. Perception of carbon dioxide and other “air-condition” parameters in the leaf cutting ant Atta cephalotes. Naturwissenschaften 1996, 83, 566–568. [Google Scholar] [CrossRef]
- Robertson, H.M.; Kent, L.B. Evolution of the gene lineage encoding the carbon dioxide receptor in insects. J. Insect Sci. 2009, 9, 19. [Google Scholar] [CrossRef] [Green Version]
- Benton, R.; Vannice, K.; Gomezdiaz, C.; Vosshall, L. Variant Ionotropic Glutamate Receptors as Chemosensory Receptors in Drosophila. Cell 2009, 136, 149–162. [Google Scholar] [CrossRef] [Green Version]
- Abuin, L.; Bargeton, B.; Ulbrich, M.H.; Isacoff, E.Y.; Kellenberger, S.; Benton, R. Functional architecture of olfactory ionotropic glutamate receptors. Neuron 2011, 69, 44–60. [Google Scholar] [CrossRef] [Green Version]
- Croset, V.; Rytz, R.; Cummins, S.F.; Budd, A.; Brawand, D.; Kaessmann, H.; Gibson, T.J.; Benton, R. Ancient protostome origin of chemosensory ionotropic glutamate receptors and the evolution of insect taste and olfaction. PLoS Genet. 2010, 6, e1001064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishida, Y.; Leal, W.S. Rapid inactivation of a moth pheromone. Proc. Natl. Acad. Sci. USA 2005, 102, 14075–14079. [Google Scholar] [CrossRef] [Green Version]
- Vogt, R.G.; Riddiford, L.M. Pheromone binding and inactivation by moth antennae. Nature 1981, 293, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Vogt, R.G.; Riddiford, L.M.; Prestwich, G.D. Kinetic properties of a sex pheromone-degrading enzyme: The sensillar esterase of Antheraea polyphemus. Proc. Natl. Acad. Sci. USA 1985, 82, 8827–8831. [Google Scholar] [CrossRef] [Green Version]
- Larter, N.K.; Sun, J.S.; Carlson, J.R. Organization and function of Drosophila odorant binding proteins. Elife 2016, 5. [Google Scholar] [CrossRef]
- Xiao, S.K.; Sun, J.S.; Carlson, J.R. Robust olfactory responses in the absence of odorant binding proteins. Elife 2019, 8. [Google Scholar] [CrossRef]
- Pask, G.; Ray, A. Insect Olfactory Receptors: An Interface between Chemistry and Biology. In Chemosensory Transduction: The Detection of Odors, Tastes, and Other Chemostimuli, 1st ed.; Zufall, F., Munger, S.D., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 101–122. [Google Scholar]
- Nakanishi, A.; Nishino, H.; Watanabe, H.; Yokohari, F.; Nishikawa, M. Sex-specific antennal sensory system in the ant Camponotus japonicus: Structure and distribution of sensilla on the flagellum. Cell Tissue Res. 2009, 338, 79–97. [Google Scholar] [CrossRef]
- McKenzie, S.K.; Fetter-Pruneda, I.; Ruta, V.; Kronauer, D.J.C. Transcriptomics and neuroanatomy of the clonal raider ant implicate an expanded clade of odorant receptors in chemical communication. Proc. Natl. Acad. Sci. USA 2016, 113, 14091–14096. [Google Scholar] [CrossRef] [Green Version]
- Ozaki, M.; Wada-Katsumata, A.; Fujikawa, K.; Iwasaki, M.; Yokohari, F.; Satoji, Y.; Nisimura, T.; Yamaoka, R. Ant nestmate and non-nestmate discrimination by a chemosensory sensillum. Science 2005, 309, 311–314. [Google Scholar] [CrossRef] [Green Version]
- Sharma, K.R.; Enzmann, B.L.; Schmidt, Y.; Moore, D.; Jones, G.R.; Parker, J.; Berger, S.L.; Reinberg, D.; Zwiebel, L.J.; Breit, B.; et al. Cuticular Hydrocarbon Pheromones for Social Behavior and Their Coding in the Ant Antenna. Cell Rep. 2015, 12, 1261–1271. [Google Scholar] [CrossRef]
- Esslen, J.; Kaissling, K.E. Zahl und Verteilung antennaler Sensillen bei der Honigbiene (Apis mellifera L.). Zoomorphology 1976, 83, 227–251. [Google Scholar] [CrossRef]
- Nishino, H.; Nishikawa, M.; Mizunami, M.; Yokohari, F. Functional and topographic segregation of glomeruli revealed by local staining of antennal sensory neurons in the honeybee Apis mellifera. J. Comp. Neurol. 2009, 515, 161–180. [Google Scholar] [CrossRef] [PubMed]
- Guillem, R.M.; Drijfhout, F.P.; Martin, S.J. Species-Specific Cuticular Hydrocarbon Stability within European Myrmica Ants. J. Chem. Ecol. 2016, 42, 1052–1062. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.J.; Helanterä, H.; Drijfhout, F.P. Evolution of species-specific cuticular hydrocarbon patterns in Formica ants. Biol. J. Linn. Soc. 2008, 95, 131–140. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.J.; Helantera, H.; Drijfhout, F.P. Colony-specific hydrocarbons identify nest mates in two species of Formica ant. J. Chem. Ecol. 2008, 34, 1072–1080. [Google Scholar] [CrossRef]
- Guerrieri, F.J.; Nehring, V.; Jorgensen, C.G.; Nielsen, J.; Galizia, C.G.; d’Ettorre, P. Ants recognize foes and not friends. Proc. R. Soc. B-Biol. Sci. 2009, 276, 2461–2468. [Google Scholar] [CrossRef] [Green Version]
- Greene, M.J.; Gordon, D.M. Social insects: Cuticular hydrocarbons inform task decisions. Nature 2003, 423, 32. [Google Scholar] [CrossRef]
- Endler, A.; Liebig, J.; Schmitt, T.; Parker, J.E.; Jones, G.R.; Schreier, P.; Hölldobler, B. Surface hydrocarbons of queen eggs regulate worker reproduction in a social insect. Proc. Natl. Acad. Sci. USA 2004, 101, 2945–2950. [Google Scholar] [CrossRef] [Green Version]
- Takeichi, Y.; Uebi, T.; Miyazaki, N.; Murata, K.; Yasuyama, K.; Inoue, K.; Suzaki, T.; Kubo, H.; Kajimura, N.; Takano, J.; et al. Putative Neural Network Within an Olfactory Sensory Unit for Nestmate and Non-nestmate Discrimination in the Japanese Carpenter Ant: The Ultra-structures and Mathematical Simulation. Front. Cell. Neurosci. 2018, 12, 310. [Google Scholar] [CrossRef]
- Su, C.Y.; Menuz, K.; Reisert, J.; Carlson, J.R. Non-synaptic inhibition between grouped neurons in an olfactory circuit. Nature 2012, 492, 66–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandstaetter, A.S.; Rössler, W.; Kleineidam, C.J. Friends and Foes from an Ant Brain’s Point of View—Neuronal Correlates of Colony Odors in a Social Insect. PLoS ONE 2011, 6, e21383–e21392. [Google Scholar] [CrossRef] [Green Version]
- Brandstaetter, A.S.; Kleineidam, C.J. Distributed representation of social odors indicates parallel processing in the antennal lobe of ants. J. Neurophysiol. 2011, 106, 2437–2449. [Google Scholar] [CrossRef] [Green Version]
- Bos, N.; d’Ettorre, P. Recognition of Social Identity in Ants. Front. Psychol. 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- d’Ettorre, P.; Deisig, N.; Sandoz, J.-C. Decoding ants’ olfactory system sheds light on the evolution of social communication. Proc. Natl. Acad. Sci. USA 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hildebrand, J.G.; Shepherd, G.M. Mechanisms of olfactory discrimination: Converging evidence for common principles across phyla. Annu. Rev. Neurosci. 1997, 20, 595–631. [Google Scholar] [CrossRef]
- Strausfeld, N.J.; Hirth, F. Deep homology of arthropod central complex and vertebrate basal ganglia. Science 2013, 340, 157–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galizia, C.G.; Sachse, S. Odor Coding in Insects. In The Neurobiology of Olfaction; Menini, A., Ed.; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Wilson, R.I.; Mainen, Z.F. Early events in olfactory processing. Annu. Rev. Neurosci. 2006, 29, 163–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Couto, A.; Alenius, M.; Dickson, B.J. Molecular, anatomical, and functional organization of the Drosophila olfactory system. Curr. Biol. 2005, 15, 1535–1547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fishilevich, E.; Vosshall, L.B. Genetic and functional subdivision of the Drosophila antennal lobe. Curr. Biol. 2005, 15, 1548–1553. [Google Scholar] [CrossRef] [Green Version]
- Younger, M.A.; Herre, M.; Ehrlich, A.R.; Gong, Z.; Gilbert, Z.N.; Rahiel, S.; Matthews, B.J.; Vosshall, L.B. Non-canonical odor coding ensures unbreakable mosquito attraction to humans. bioRxiv 2020. [Google Scholar] [CrossRef]
- Task, D.; Lin, C.-C.; Afify, A.; Li, H.; Vulpe, A.; Menuz, K.; Potter, C.J. Widespread Polymodal Chemosensory Receptor Expression in Drosophila Olfactory Neurons. bioRxiv 2020. [Google Scholar] [CrossRef]
- Christensen, T.A.; Waldrop, B.R.; Hildebrand, J.G. GABAergic mechanisms that shape the temporal response to odors in moth olfactory projection neurons. Ann. N. Y. Acad. Sci. 1998, 855, 475–481. [Google Scholar] [CrossRef]
- Sachse, S.; Galizia, C.G. Role of inhibition for temporal and spatial odor representation in olfactory output neurons: A calcium imaging study. J. Neurophysiol. 2002, 87, 1106–1117. [Google Scholar] [CrossRef] [Green Version]
- Sachse, S.; Peele, P.; Silbering, A.F.; Gühmann, M.; Galizia, C.G. Role of histamine as a putative inhibitory transmitter in the honeybee antennal lobe. Front. Zool. 2006, 3, 22. [Google Scholar] [CrossRef] [Green Version]
- Olsen, S.R.; Bhandawat, V.; Wilson, R.I. Excitatory interactions between olfactory processing channels in the Drosophila antennal lobe. Neuron 2007, 54, 89–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, Y.; Claridge-Chang, A.; Sjulson, L.; Pypaert, M.; Miesenböck, G. Excitatory local circuits and their implications for olfactory processing in the fly antennal lobe. Cell 2007, 128, 601–612. [Google Scholar] [CrossRef] [Green Version]
- Kirschner, S.; Kleineidam, C.J.; Zube, C.; Rybak, J.; Grünewald, B.; Rössler, W. Dual olfactory pathway in the honeybee, Apis mellifera. J. Comp. Neurol. 2006, 499, 933–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zube, C.; Kleineidam, C.J.; Kirschner, S.; Neef, J.; Rossler, W. Organization of the olfactory pathway and odor processing in the antennal lobe of the ant Camponotus floridanus. J. Comp. Neurol. 2008, 506, 425–441. [Google Scholar] [CrossRef] [Green Version]
- Hammer, M.; Menzel, R. Learning and memory in the honeybee. J. Neurosci. 1995, 15, 1617–1630. [Google Scholar] [CrossRef] [Green Version]
- Erber, J.; Masuhr, T.; Menzel, R. Localization of short-term memory in the brain of the bee, Apis mellifera. Physiol. Entomol. 1980, 5, 343–358. [Google Scholar] [CrossRef]
- Schultzhaus, J.N.; Saleem, S.; Iftikhar, H.; Carney, G.E. The role of the Drosophila lateral horn in olfactory information processing and behavioral response. J. Insect Physiol. 2017, 98, 29–37. [Google Scholar] [CrossRef]
- Maresh, A.; Rodriguez Gil, D.; Whitman, M.C.; Greer, C.A. Principles of Glomerular Organization in the Human Olfactory Bulb—Implications for Odor Processing. PLoS ONE 2008, 3, e2640. [Google Scholar] [CrossRef] [Green Version]
- Serizawa, S.; Miyamichi, K.; Sakano, H. One neuron-one receptor rule in the mouse olfactory system. Trends Genet. 2004, 20, 648–653. [Google Scholar] [CrossRef]
- Royet, J.P.; Souchier, C.; Jourdan, F.; Ploye, H. Morphometric study of the glomerular population in the mouse olfactory bulb: Numerical density and size distribution along the rostrocaudal axis. J. Comp. Neurol. 1988, 270, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Buck, L.; Axel, R. A novel multigene family may encode odorant receptors: A molecular basis for odor recognition. Cell 1991, 65, 175–187. [Google Scholar] [CrossRef]
- Stocker, R.F.; Lienhard, M.C.; Borst, A.; Fischbach, K.F. Neuronal architecture of the antennal lobe in Drosophila melanogaster. Cell Tissue Res. 1990, 262, 9–34. [Google Scholar] [CrossRef] [PubMed]
- Kelber, C.; Rössler, W.; Kleineidam, C.J. Phenotypic plasticity in number of glomeruli and sensory innervation of the antennal lobe in leaf-cutting ant workers (A. vollenweideri). Dev. Neurobiol. 2010, 70, 222–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, S.M.; Napper, R.M.; Mercer, A.R. Foraging experience, glomerulus volume, and synapse number: A stereological study of the honey bee antennal lobe. J. Neurobiol. 2004, 60, 40–50. [Google Scholar] [CrossRef]
- Sigg, D.; Thompson, C.M.; Mercer, A.R. Activity-dependent changes to the brain and behavior of the honey bee, Apis mellifera (L.). J. Neurosci. 1997, 17, 7148–7156. [Google Scholar] [CrossRef] [Green Version]
- Arnold, G.; Budharugsa, S.; Masson, C. Organization of the Antennal Lobe in the Queen Honey Bee, Apis-Mellifera L (Hymenoptera, Apidae). Int. J. Insect Morphol. 1988, 17, 185–195. [Google Scholar] [CrossRef]
- Mysore, K.; Subramanian, K.A.; Sarasij, R.C.; Suresh, A.; Shyamala, B.V.; VijayRaghavan, K.; Rodrigues, V. Caste and sex specific olfactory glomerular organization and brain architecture in two sympatric ant species Camponotus sericeus and Camponotus compressus (Fabricius, 1798). Arthropod Struct. Dev. 2009, 38, 485–497. [Google Scholar] [CrossRef]
- Jernigan, C.M.; Halby, R.; Gerkin, R.C.; Sinakevitch, I.; Locatelli, F.; Smith, B.H. Experience-dependent tuning of early olfactory processing in the adult honey bee, Apis mellifera. J. Exp. Biol. 2020, 223. [Google Scholar] [CrossRef]
- Kleineidam, C.J.; Obermayer, M.; Halbich, W.; Rossler, W. A macroglomerulus in the antennal lobe of leaf-cutting ant workers and its possible functional significance. Chem. Senses 2005, 30, 383–392. [Google Scholar] [CrossRef]
- Nishikawa, M.; Nishino, H.; Misaka, Y.; Kubota, M.; Tsuji, E.; Satoji, Y.; Ozaki, M.; Yokohari, F. Sexual dimorphism in the antennal lobe of the ant Camponotus japonicus. Zool. Sci. 2008, 25, 195–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, A.; Nishino, H.; Watanabe, H.; Yokohari, F.; Nishikawa, M. Sex-specific antennal sensory system in the ant Camponotus japonicus: Glomerular organizations of antennal lobes. J. Comp. Neurol. 2010, 518, 2186–2201. [Google Scholar] [CrossRef]
- Arnold, G.; Masson, C.; Budharugsa, S. Comparative-Study of the Antennal Lobes and Their Afferent Pathway in the Worker Bee and the Drone (Apis-Mellifera). Cell Tissue Res. 1985, 242, 593–605. [Google Scholar] [CrossRef]
- Sandoz, J.C. Odour-evoked responses to queen pheromone components and to plant odours using optical imaging in the antennal lobe of the honey bee drone Apis mellifera L. J. Exp. Biol. 2006, 209, 3587–3598. [Google Scholar] [CrossRef] [Green Version]
- Dupuy, F.; Josens, R.; Giurfa, M.; Sandoz, J.-C. Calcium imaging in the ant Camponotus fellah reveals a conserved odour-similarity space in insects and mammals. BMC Neurosci. 2010, 11, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galizia, C.G.; Sachse, S.; Rappert, A.; Menzel, R. The glomerular code for odor representation is species specific in the honeybee Apis mellifera. Nat. Neurosci. 1999, 2, 473–478. [Google Scholar] [CrossRef] [Green Version]
- Galizia, C.G.; Menzel, R.; Holldobler, B. Optical imaging of odor-evoked glomerular activity patterns in the antennal lobes of the ant camponotus rufipes. Naturwissenschaften 1999, 86, 533–537. [Google Scholar] [CrossRef]
- Yamagata, N.; Nishino, H.; Mizunami, M. Pheromone-sensitive glomeruli in the primary olfactory centre of ants. Proc. Biol. Sci. 2006, 273, 2219–2225. [Google Scholar] [CrossRef]
- Hansson, B.S.; Christensen, T.A.; Hildebrand, J.G. Functionally distinct subdivisions of the macroglomerular complex in the antennal lobe of the male sphinx moth Manduca sexta. J. Comp. Neurol. 1991, 312, 264–278. [Google Scholar] [CrossRef]
- Varela, N.; Avilla, J.; Gemeno, C.; Anton, S. Ordinary glomeruli in the antennal lobe of male and female tortricid moth Grapholita molesta (Busck) (Lepidoptera: Tortricidae) process sex pheromone and host-plant volatiles. J. Exp. Biol. 2011, 214, 637. [Google Scholar] [CrossRef] [Green Version]
- Jefferis, G.S.; Vyas, R.M.; Berdnik, D.; Ramaekers, A.; Stocker, R.F.; Tanaka, N.K.; Ito, K.; Luo, L. Developmental origin of wiring specificity in the olfactory system of Drosophila. Development 2004, 131, 117–130. [Google Scholar] [CrossRef] [Green Version]
- Larsson, M.C.; Domingos, A.I.; Jones, W.D.; Chiappe, M.E.; Amrein, H.; Vosshall, L.B. Or83b encodes a broadly expressed odorant receptor essential for Drosophila olfaction. Neuron 2004, 43, 703–714. [Google Scholar] [CrossRef] [Green Version]
- Lodovichi, C.; Belluscio, L. Odorant Receptors in the Formation of the Olfactory Bulb Circuitry. Physiology 2012, 27, 200–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monti-Graziadei, G.A.; Graziadei, P.P.C. Neurogenesis and neuron regeneration in the olfactory system of mammals. II. Degeneration and reconstitution of the olfactory sensory neurons after axotomy. J. Neurocytol. 1979, 8, 197–213. [Google Scholar] [CrossRef]
- Yan, H.; Opachaloemphan, C.; Mancini, G.; Yang, H.; Gallitto, M.; Mlejnek, J.; Leibholz, A.; Haight, K.; Ghaninia, M.; Huo, L.; et al. An Engineered orco Mutation Produces Aberrant Social Behavior and Defective Neural Development in Ants. Cell 2017, 170, 736–747.e9. [Google Scholar] [CrossRef] [PubMed]
- Trible, W.; Olivos-Cisneros, L.; McKenzie, S.K.; Saragosti, J.; Chang, N.C.; Matthews, B.J.; Oxley, P.R.; Kronauer, D.J.C. Orco Mutagenesis Causes Loss of Antennal Lobe Glomeruli and Impaired Social Behavior in Ants. Cell 2017, 170, 727–735.e10. [Google Scholar] [CrossRef] [PubMed]
- Ryba, A.R.; McKenzie, S.K.; Olivos-Cisneros, L.; Clowney, E.J.; Pires, P.M.; Kronauer, D.J.C. Comparative Development of the Ant Chemosensory System. Curr. Biol. 2020, 30, 3223–3230.e4. [Google Scholar] [CrossRef]
- Galizia, C.G.; McIlwrath, S.L.; Menzel, R. A digital three-dimensional atlas of the honeybee antennal lobe based on optical sections acquired by confocal microscopy. Cell Tissue Res. 1999, 295, 383–394. [Google Scholar] [CrossRef] [Green Version]
- Branstetter, M.G.; Childers, A.K.; Cox-Foster, D.; Hopper, K.R.; Kapheim, K.M.; Toth, A.L.; Worley, K.C. Genomes of the Hymenoptera. Curr. Opin. Insect Sci. 2018, 25, 65–75. [Google Scholar] [CrossRef] [PubMed]
- Favreau, E.; Martinez-Ruiz, C.; Santiago, L.R.; Hammond, R.L.; Wurm, Y. Genes and genomic processes underpinning the social lives of ants. Curr. Opin. Insect Sci. 2018, 25, 83–90. [Google Scholar] [CrossRef]
- Smith, C.R.; Smith, C.D.; Robertson, H.M.; Helmkampf, M.; Zimin, A.; Yandell, M.; Holt, C.; Hu, H.; Abouheif, E.; Benton, R.; et al. Draft genome of the red harvester ant Pogonomyrmex barbatus. Proc. Natl. Acad. Sci. USA 2011, 108, 5667–5672. [Google Scholar] [CrossRef] [Green Version]
- Robertson, H.M.; Wanner, K.W. The chemoreceptor superfamily in the honey bee, Apis mellifera: Expansion of the odorant, but not gustatory, receptor family. Genome Res. 2006, 16, 1395–1403. [Google Scholar] [CrossRef] [Green Version]
- Wurm, Y.; Wang, J.; Riba-Grognuz, O.; Corona, M.; Nygaard, S.; Hunt, B.G.; Ingram, K.; Falquet, L.; Nipitwattanaphon, M.; Gotzek, D.; et al. The genome of the fire ant Solenopsis invicta. Proc. Natl. Acad. Sci. USA 2011, 108, 5679–5684. [Google Scholar] [CrossRef] [Green Version]
- Oxley, P.R.; Ji, L.; Fetter-Pruneda, I.; McKenzie, S.K.; Li, C.; Hu, H.; Zhang, G.; Kronauer, D.J. The genome of the clonal raider ant Cerapachys biroi. Curr. Biol. 2014, 24, 451–458. [Google Scholar] [CrossRef] [Green Version]
- Werren, J.H.; Richards, S.; Desjardins, C.A.; Niehuis, O.; Gadau, J.; Colbourne, J.K.; Nasonia Genome Working, G.; Werren, J.H.; Richards, S.; Desjardins, C.A.; et al. Functional and evolutionary insights from the genomes of three parasitoid Nasonia species. Science 2010, 327, 343–348. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.D.; Zimin, A.; Holt, C.; Abouheif, E.; Benton, R.; Cash, E.; Croset, V.; Currie, C.R.; Elhaik, E.; Elsik, C.G.; et al. Draft genome of the globally widespread and invasive Argentine ant (Linepithema humile). Proc. Natl. Acad. Sci. USA 2011, 108, 5673–5678. [Google Scholar] [CrossRef] [Green Version]
- Bonasio, R.; Li, Q.; Lian, J.; Mutti, N.S.; Jin, L.; Zhao, H.; Zhang, P.; Wen, P.; Xiang, H.; Ding, Y.; et al. Genome-wide and caste-specific DNA methylomes of the ants Camponotus floridanus and Harpegnathos saltator. Curr. Biol. 2012, 22, 1755–1764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, H.M.; Waterhouse, R.M.; Walden, K.K.O.; Ruzzante, L.; Reijnders, M.J.M.F.; Coates, B.S.; Legeai, F.; Gress, J.C.; Biyiklioglu, S.; Weaver, D.K.; et al. Genome Sequence of the Wheat Stem Sawfly, Cephus cinctus, Representing an Early-Branching Lineage of the Hymenoptera, Illuminates Evolution of Hymenopteran Chemoreceptors. Genome Biol. Evol. 2018, 10, 2997–3011. [Google Scholar] [CrossRef] [Green Version]
- Robertson, H.M.; Gadau, J.; Wanner, K.W. The insect chemoreceptor superfamily of the parasitoid jewel wasp Nasonia vitripennis. Insect Mol. Biol. 2010, 19, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Terrapon, N.; Li, C.; Robertson, H.M.; Ji, L.; Meng, X.; Booth, W.; Chen, Z.; Childers, C.P.; Glastad, K.M.; Gokhale, K.; et al. Molecular traces of alternative social organization in a termite genome. Nat. Commun. 2014, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, H.M.; Baits, R.L.; Walden, K.K.O.; Wada-Katsumata, A.; Schal, C. Enormous expansion of the chemosensory gene repertoire in the omnivorous German cockroach Blattella germanica. J. Exp. Zool. Part B Mol. Dev. Evol. 2018, 330, 265–278. [Google Scholar] [CrossRef] [Green Version]
- Chiu, Y.K.; Hsu, J.C.; Chang, T.; Huang, Y.C.; Wang, J. Mutagenesis mediated by CRISPR/Cas9 in the red imported fire ant, Solenopsis invicta. Insectes Sociaux 2020, 67, 317–326. [Google Scholar] [CrossRef]
- Higashi, S.; Yamauchi, K. Influence of a supercolonial ant Formica (Formica) yessensis Forel on the Distribution of Other Ants in Ishikari Coast. Jpn. J. Ecol. 1979, 29, 257–264. [Google Scholar] [CrossRef]
- Hölldobler, B.; Wilson, E.O. The Superorganism: The Beauty, Elegance, and Strangeness of Insect Societies; W. W. Norton & Company: New York, NY, USA, 2008; p. 544. [Google Scholar]
- Chaverra-Rodriguez, D.; Macias, V.M.; Hughes, G.L.; Pujhari, S.; Suzuki, Y.; Peterson, D.R.; Kim, D.; McKeand, S.; Rasgon, J.L. Targeted delivery of CRISPR-Cas9 ribonucleoprotein into arthropod ovaries for heritable germline gene editing. Nat. Commun. 2018, 9, 3008. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heu, C.C.; McCullough, F.M.; Luan, J.; Rasgon, J.L. CRISPR-Cas9-Based Genome Editing in the Silverleaf Whitefly (Bemisia tabaci). CRISPR J. 2020, 3, 89–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaverra-Rodriguez, D.; Dalla Benetta, E.; Heu, C.C.; Rasgon, J.L.; Ferree, P.M.; Akbari, O.S. Germline mutagenesis of Nasonia vitripennis through ovarian delivery of CRISPR-Cas9 ribonucleoprotein. Insect Mol. Biol. 2020, 29, 569–577. [Google Scholar] [CrossRef]
- Macias, V.M.; McKeand, S.; Chaverra-Rodriguez, D.; Hughes, G.L.; Fazekas, A.; Pujhari, S.; Jasinskiene, N.; James, A.A.; Rasgon, J.L. Cas9-Mediated Gene-Editing in the Malaria Mosquito Anopheles stephensi by ReMOT Control. G3 2020, 10, 1353–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, C.A.; Challagulla, A.; Jenkins, K.A.; Wise, T.G.; O’Neil, T.E.; Morris, K.R.; Tizard, M.L.; Doran, T.J. Generation of gene edited birds in one generation using sperm transfection assisted gene editing (STAGE). Transgenic Res. 2017, 26, 331–347. [Google Scholar] [CrossRef]
- den Boer, S.P.A.; Boomsma, J.J.; Baer, B. A technique to artificially inseminate leafcutter ants. Insectes Sociaux 2013, 60, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, S.T.; Park, K.Y.; Ruff, A.A.; Bakis, I.; Zwiebel, L.J. Odor coding of nestmate recognition in the eusocial ant Camponotus floridanus. J. Exp. Biol. 2020, 223, jeb215400. [Google Scholar] [CrossRef]
- Luger, K.; Mäder, A.W.; Richmond, R.K.; Sargent, D.F.; Richmond, T.J. Crystal structure of the nucleosome core particle at 2.8 Å resolution. Nature 1997, 389, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Kouzarides, T. Chromatin modifications and their function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef] [Green Version]
- Cedar, H.; Bergman, Y. Linking DNA methylation and histone modification: Patterns and paradigms. Nat. Rev. Genet. 2009, 10, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Clowney, E.J.; Magklara, A.; Colquitt, B.M.; Pathak, N.; Lane, R.P.; Lomvardas, S. High-throughput mapping of the promoters of the mouse olfactory receptor genes reveals a new type of mammalian promoter and provides insight into olfactory receptor gene regulation. Genome Res. 2011, 21, 1249–1259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magklara, A.; Yen, A.; Colquitt, B.M.; Clowney, E.J.; Allen, W.; Markenscoff-Papadimitriou, E.; Evans, Z.A.; Kheradpour, P.; Mountoufaris, G.; Carey, C.; et al. An epigenetic signature for monoallelic olfactory receptor expression. Cell 2011, 145, 555–570. [Google Scholar] [CrossRef] [Green Version]
- Sim, C.K.; Perry, S.; Tharadra, S.K.; Lipsick, J.S.; Ray, A. Epigenetic regulation of olfactory receptor gene expression by the Myb-MuvB/dREAM complex. Genes Dev. 2012, 26, 2483–2498. [Google Scholar] [CrossRef] [Green Version]
- Glastad, K.M.; Hunt, B.G.; Goodisman, M.A.D. DNA Methylation and Chromatin Organization in Insects: Insights from the Ant Camponotus floridanus. Genome Biol. Evol. 2015, 7, 931–942. [Google Scholar] [CrossRef] [Green Version]
- Simola, D.F.; Ye, C.; Mutti, N.S.; Dolezal, K.; Bonasio, R.; Liebig, J.; Reinberg, D.; Berger, S.L. A chromatin link to caste identity in the carpenter ant Camponotus floridanus. Genome Res. 2013, 23, 486–496. [Google Scholar] [CrossRef] [Green Version]
- Simola, D.F.; Graham, R.J.; Brady, C.M.; Enzmann, B.L.; Desplan, C.; Ray, A.; Zwiebel, L.J.; Bonasio, R.; Reinberg, D.; Liebig, J.; et al. Epigenetic (re)programming of caste-specific behavior in the ant Camponotus floridanus. Science 2016, 351, aac6633. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferguson, S.T.; Bakis, I.; Zwiebel, L.J. Advances in the Study of Olfaction in Eusocial Ants. Insects 2021, 12, 252. https://doi.org/10.3390/insects12030252
Ferguson ST, Bakis I, Zwiebel LJ. Advances in the Study of Olfaction in Eusocial Ants. Insects. 2021; 12(3):252. https://doi.org/10.3390/insects12030252
Chicago/Turabian StyleFerguson, Stephen T., Isaac Bakis, and Laurence J. Zwiebel. 2021. "Advances in the Study of Olfaction in Eusocial Ants" Insects 12, no. 3: 252. https://doi.org/10.3390/insects12030252
APA StyleFerguson, S. T., Bakis, I., & Zwiebel, L. J. (2021). Advances in the Study of Olfaction in Eusocial Ants. Insects, 12(3), 252. https://doi.org/10.3390/insects12030252