Antagonism between PTP1B and PTK Mediates Adults’ Insulin-Like Signaling Regulation of Egg Diapause in the Migratory Locust


 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Rearing Conditions
2.2. RNAi Treament
2.2.1. mRNA Quantification
2.2.2. RNAi
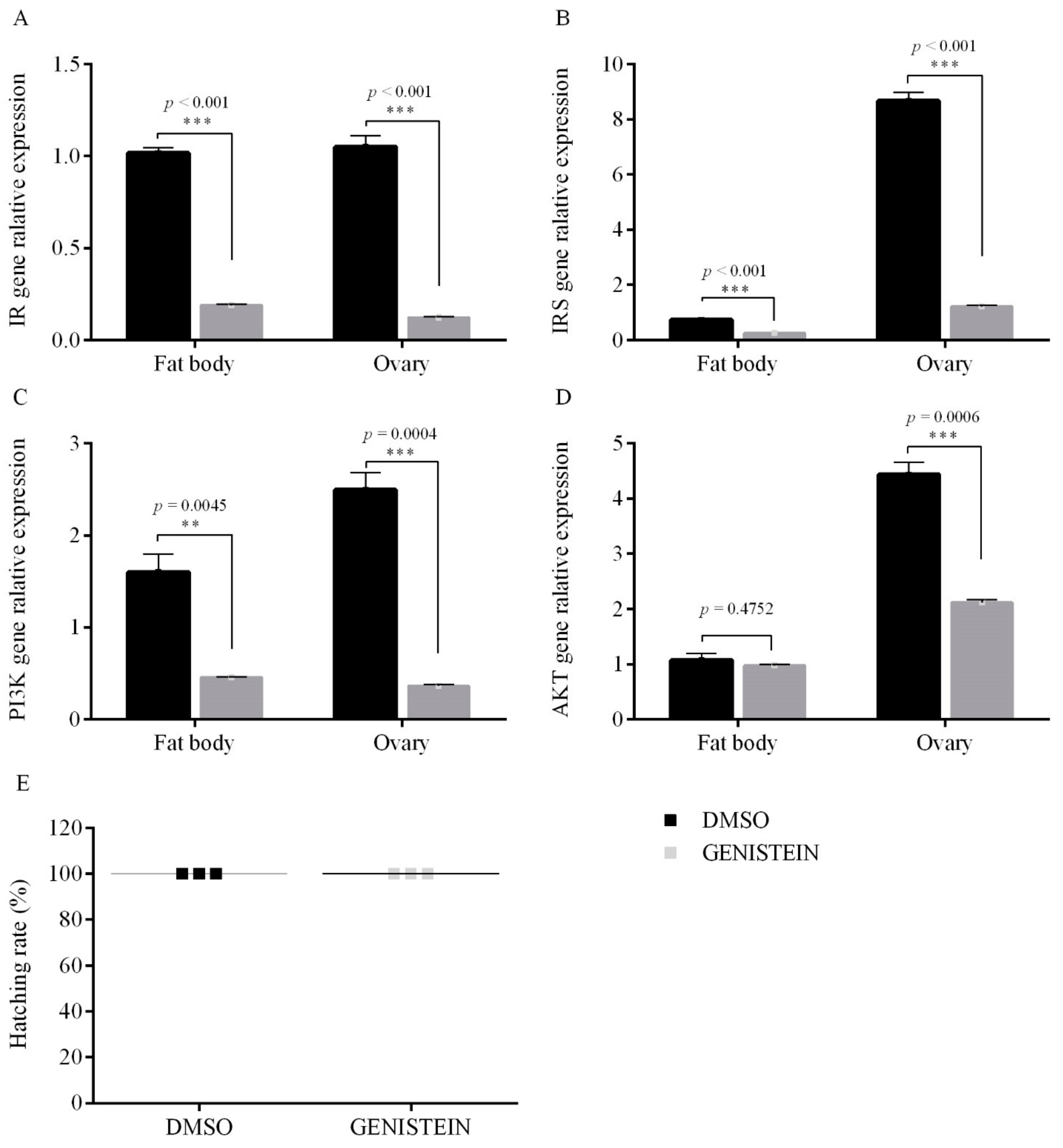
2.3. Inhibitor Treatments
2.4. Prx V Protein Purification and Injection
2.5. Collection of Eggs
2.6. Statistical Analysis
3. Results
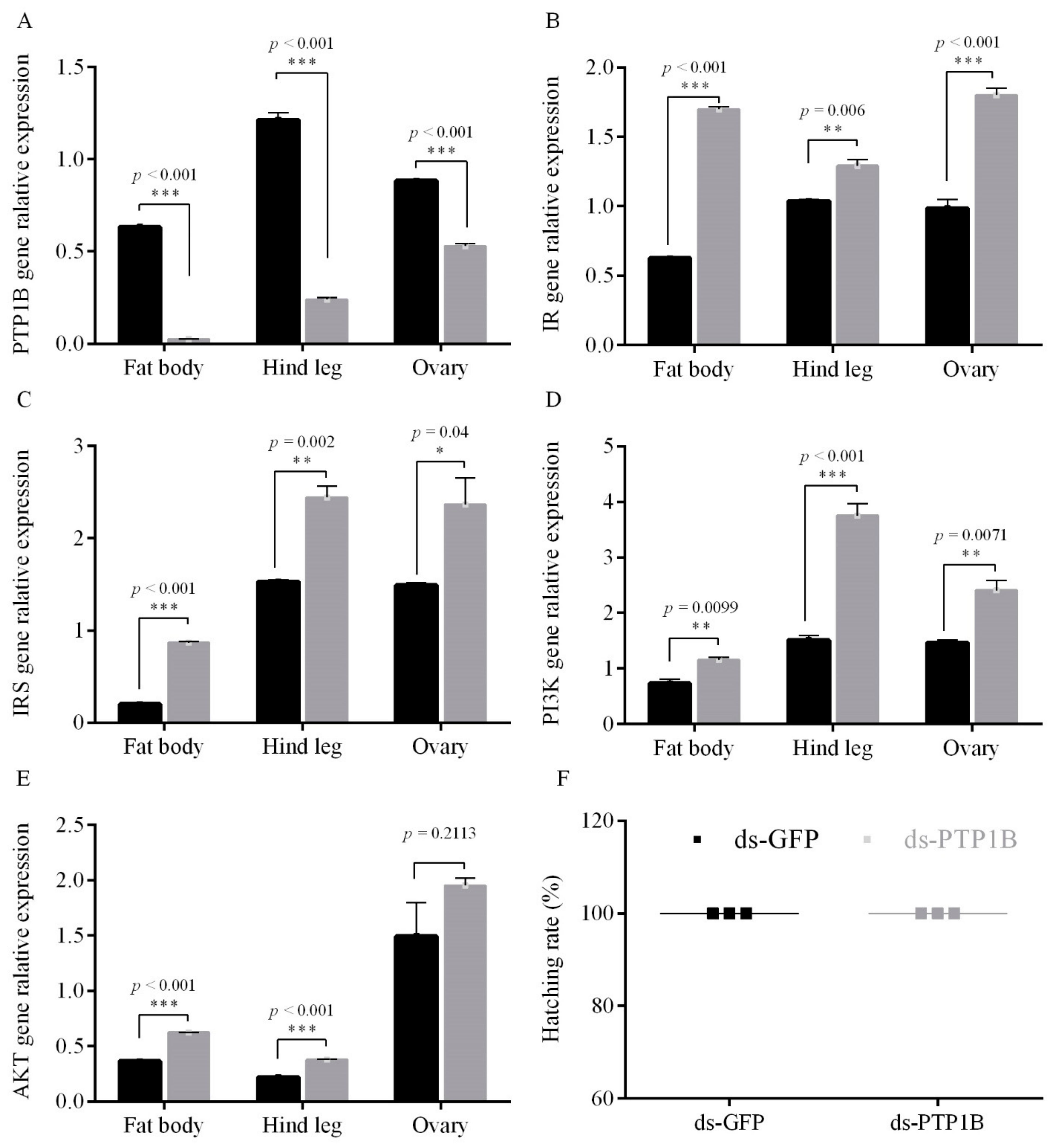
3.1. Female Adult PTP1B Promotes Egg Diapause under the Condition of Diapause Induction
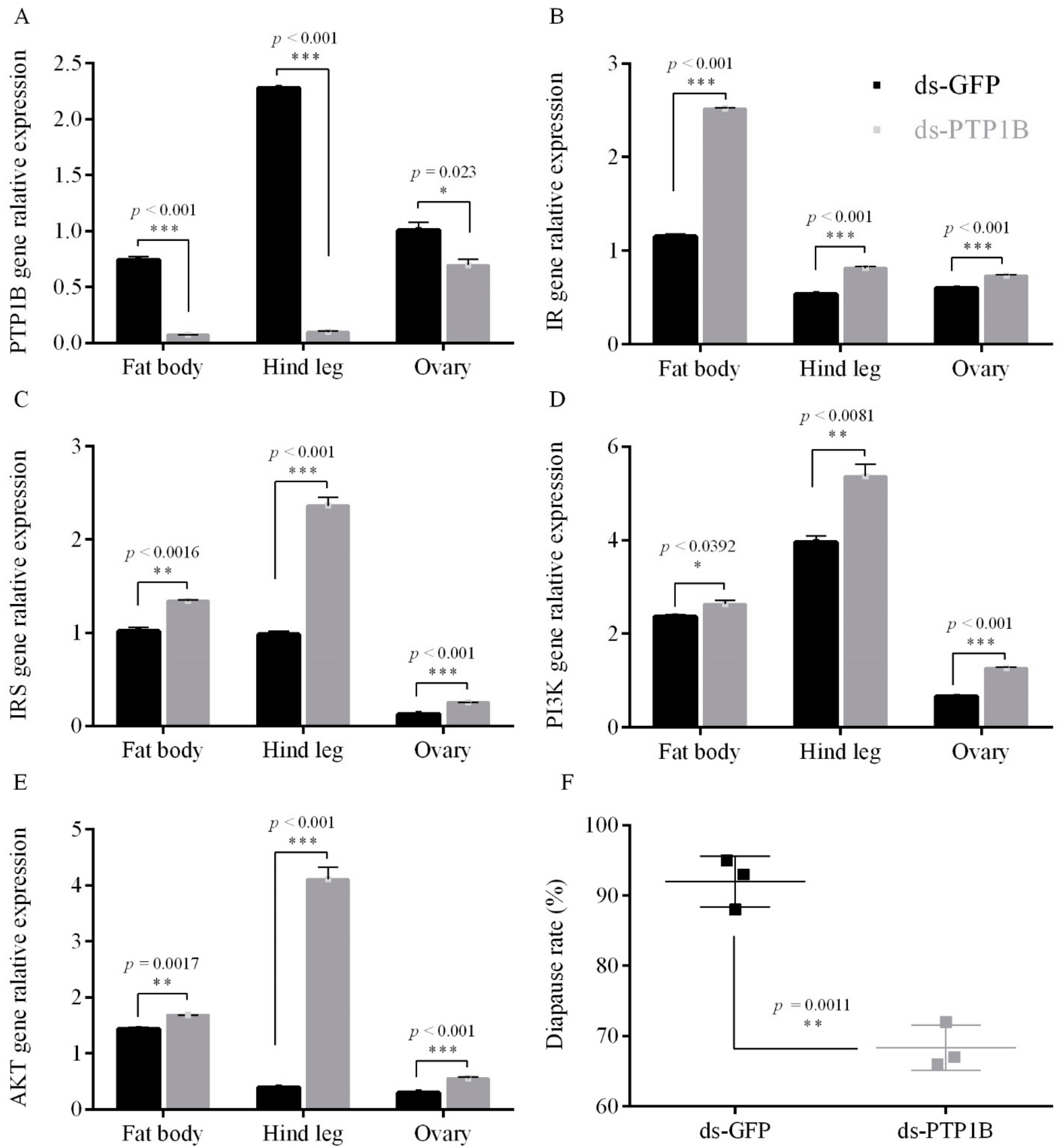
3.1.1. PTP1B RNAi Increased the Expression Level of the ISP Genes and Inhibted Diapause under Short Photoperiod Conditions
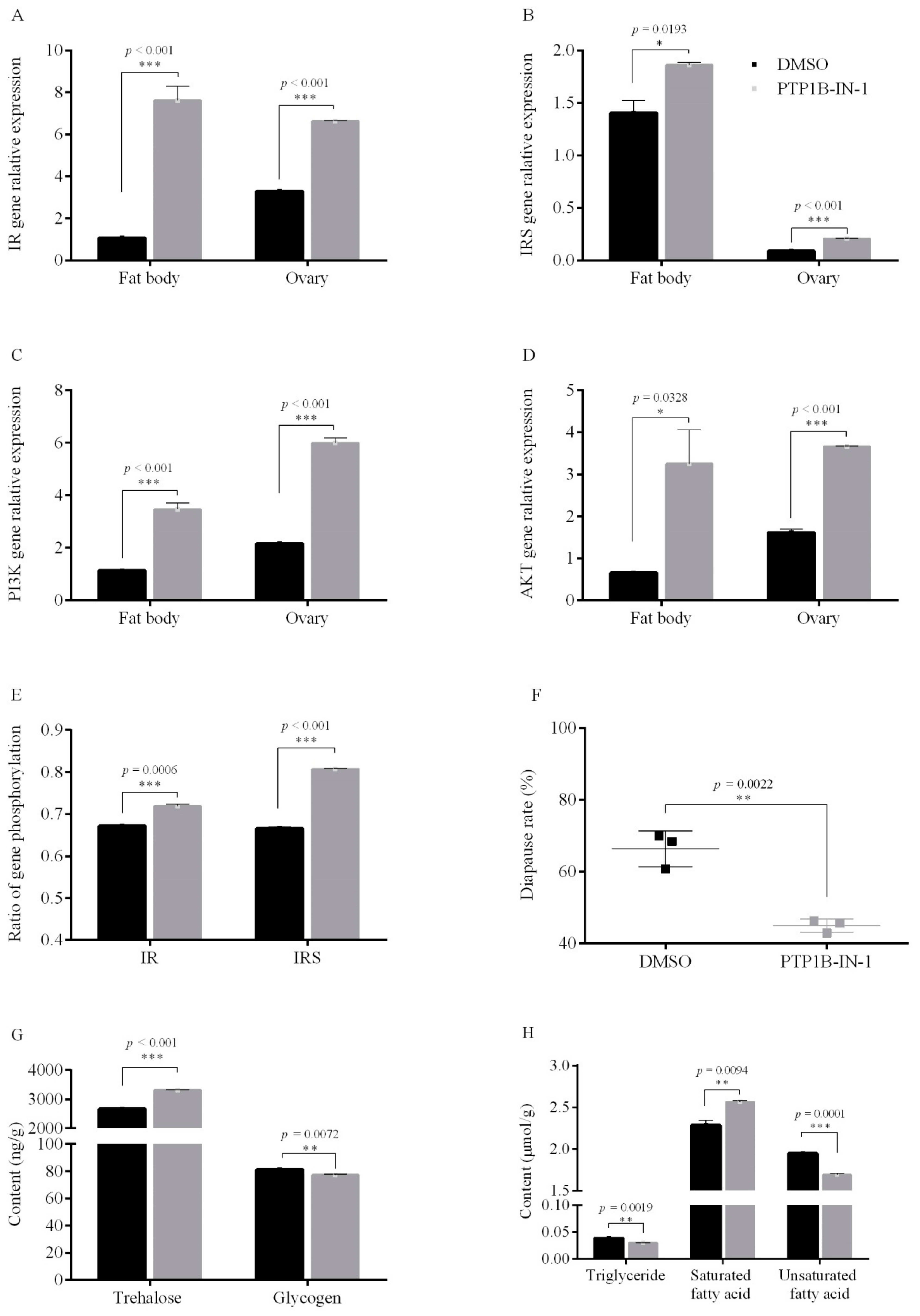
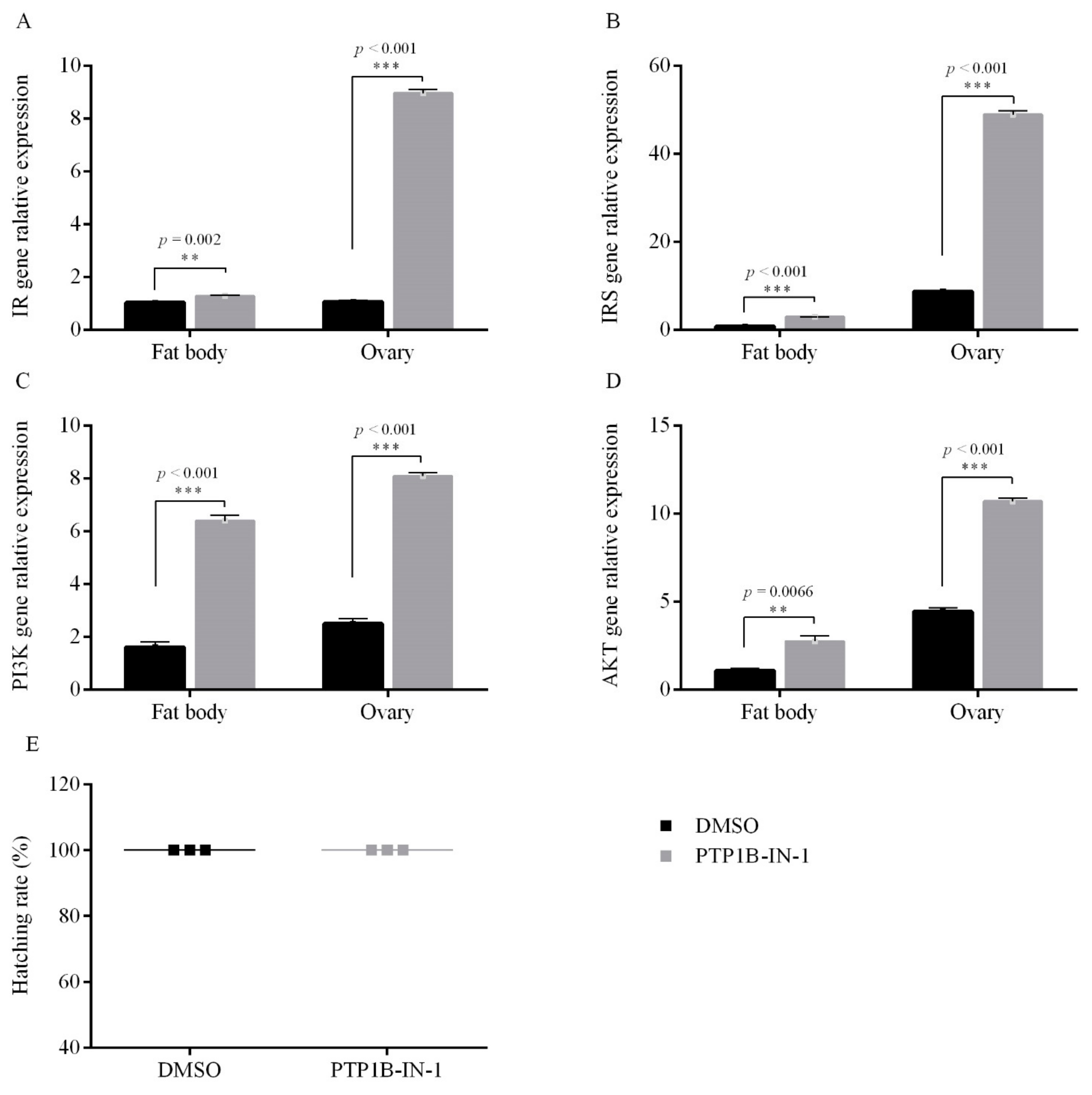
3.1.2. PTP1B Activity on Egg Diapause
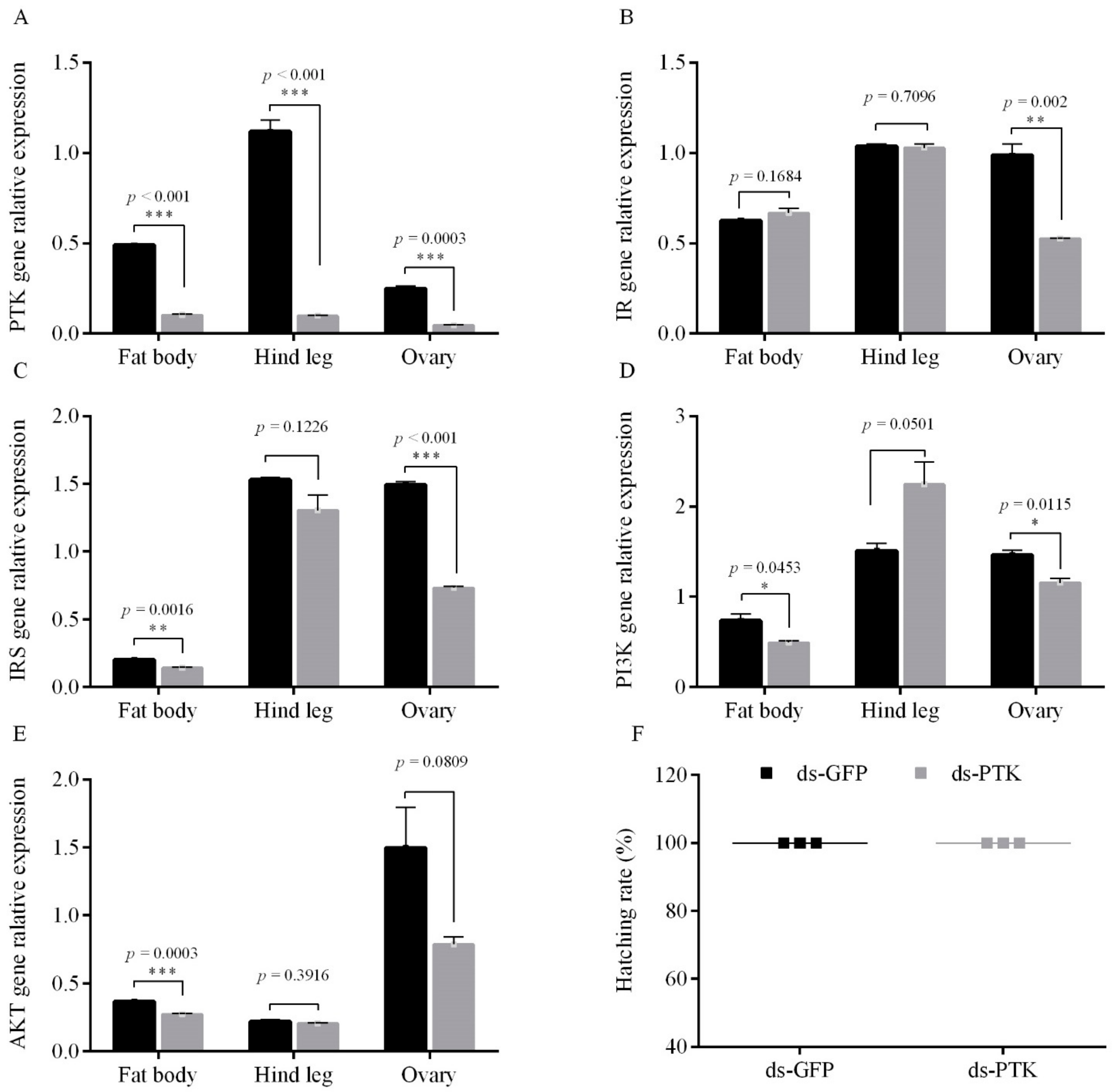
3.2. Female Adult PTK Inhibts Egg Diapause under the Condition of Diapause Induction
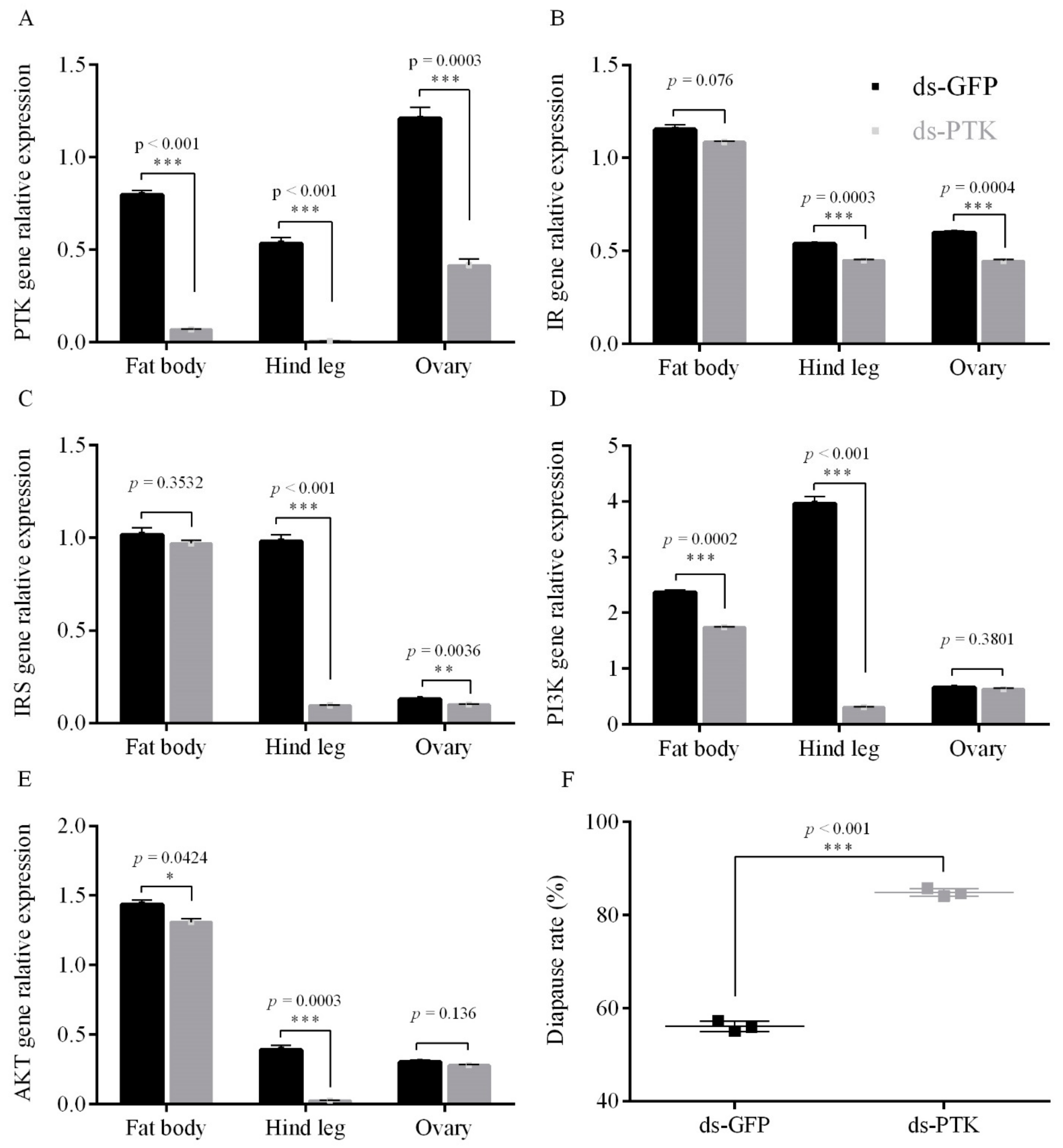
3.2.1. PTK RNAi Decreased the Expression Level of the ISP Genes and Promoting Diapause under Short Photoperiod Conditions
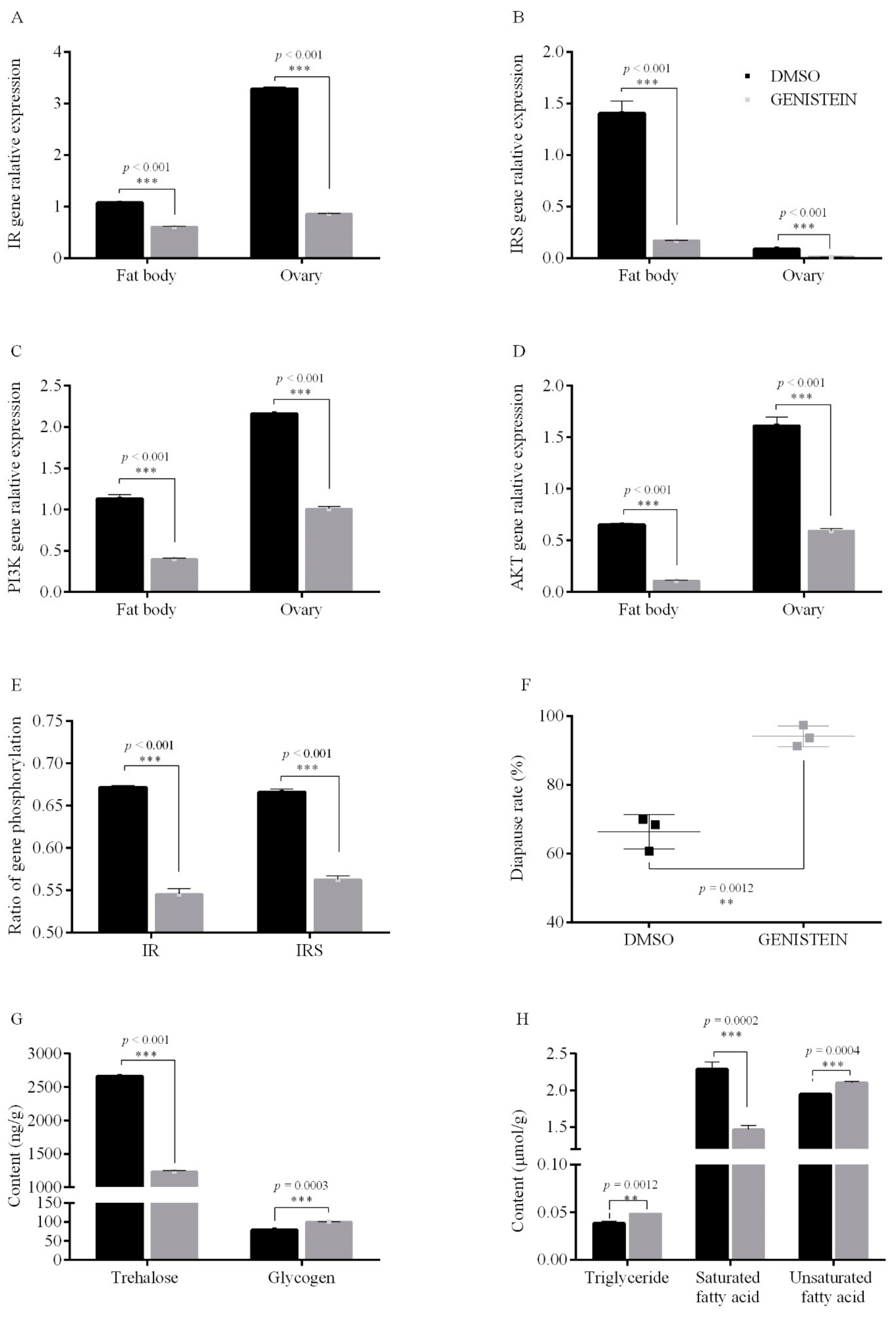
3.2.2. PTK Inhibit on Egg Diapause
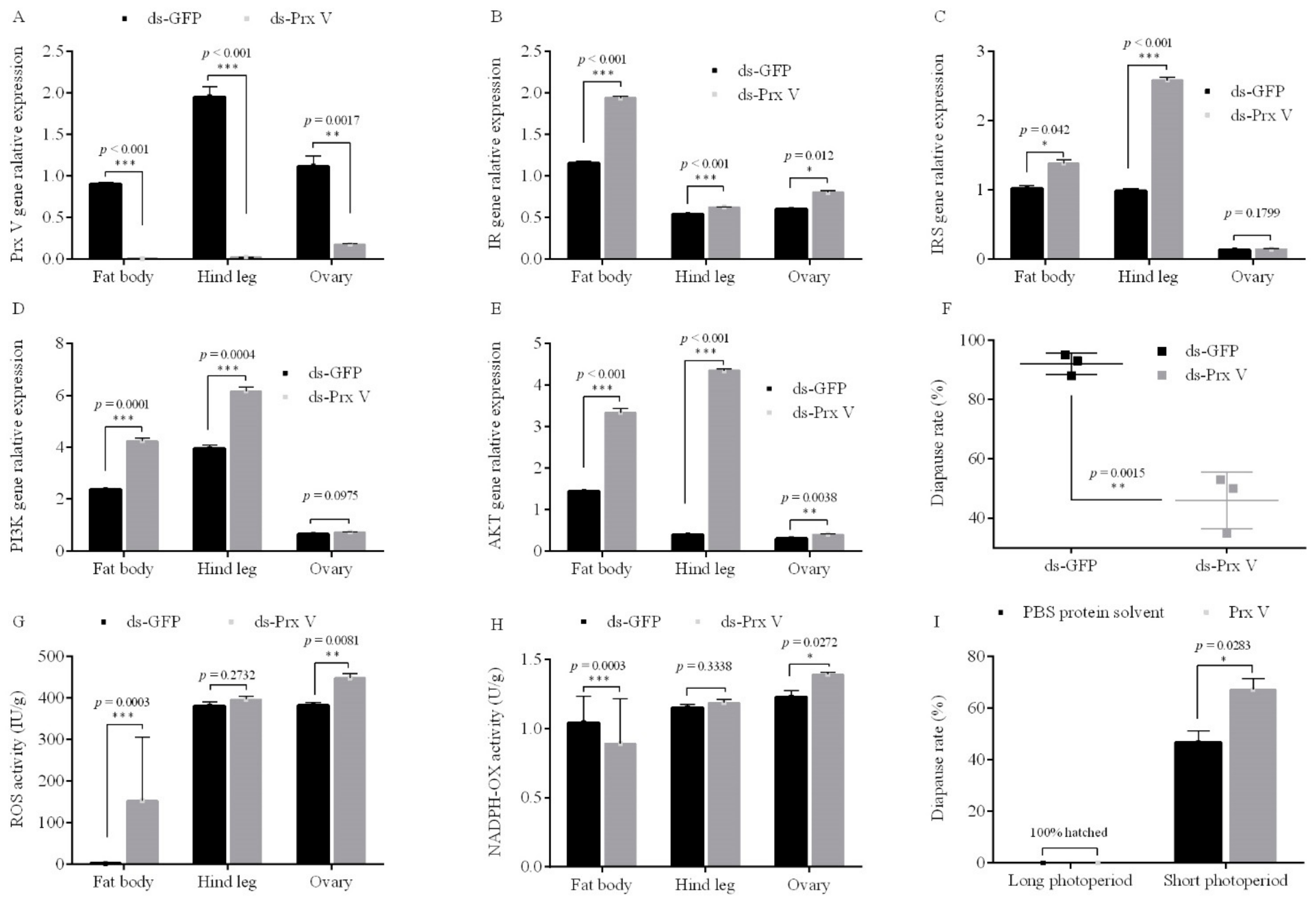
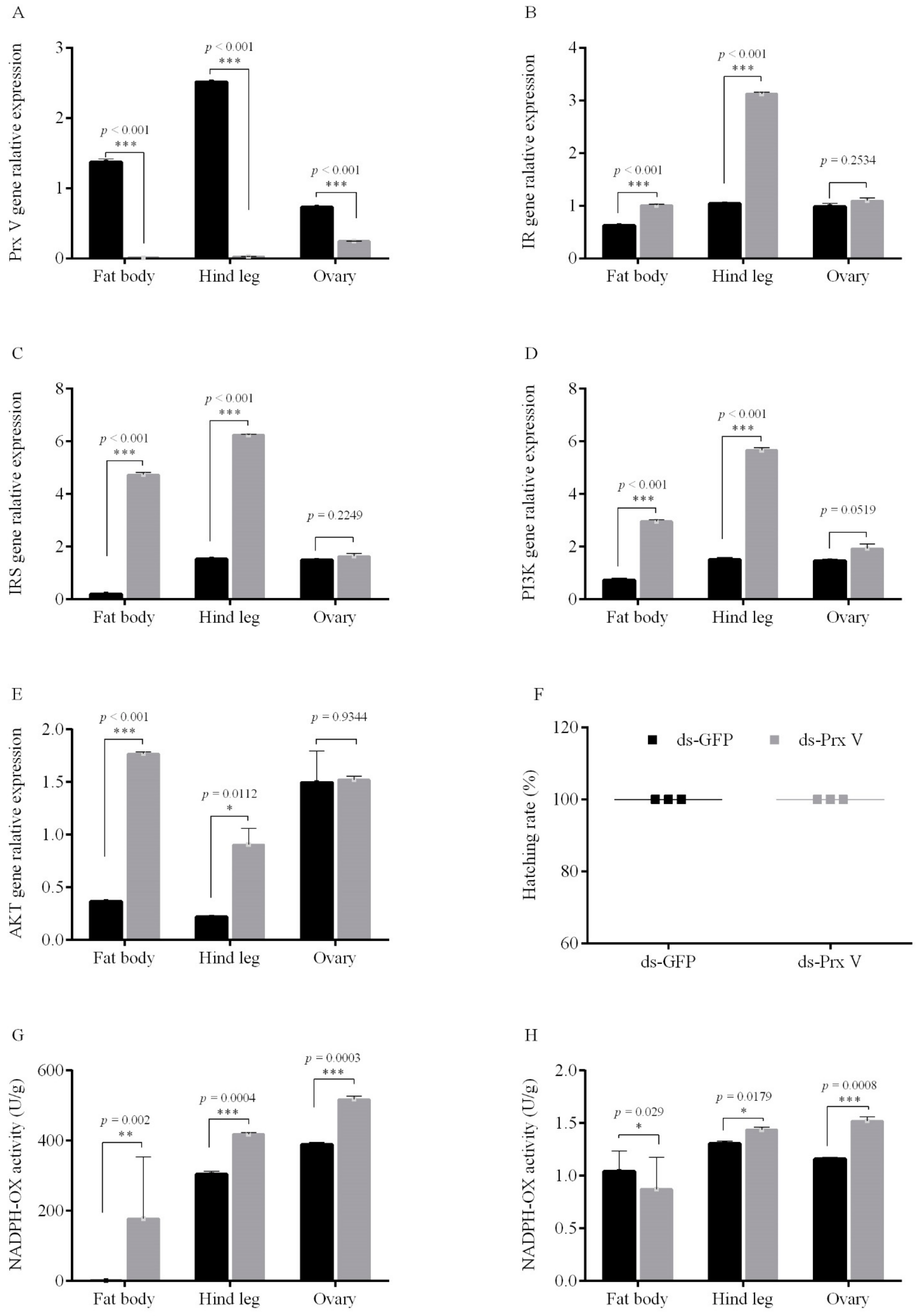
3.3. Female Adult Prx V Promoted Egg Diapause under the Condition of Diapause Induction
3.3.1. RNAi of Prx V Inhibits Eggs’ Diapause Incidence
3.3.2. Injected Prx V Protein Induces Eggs’ Diapause
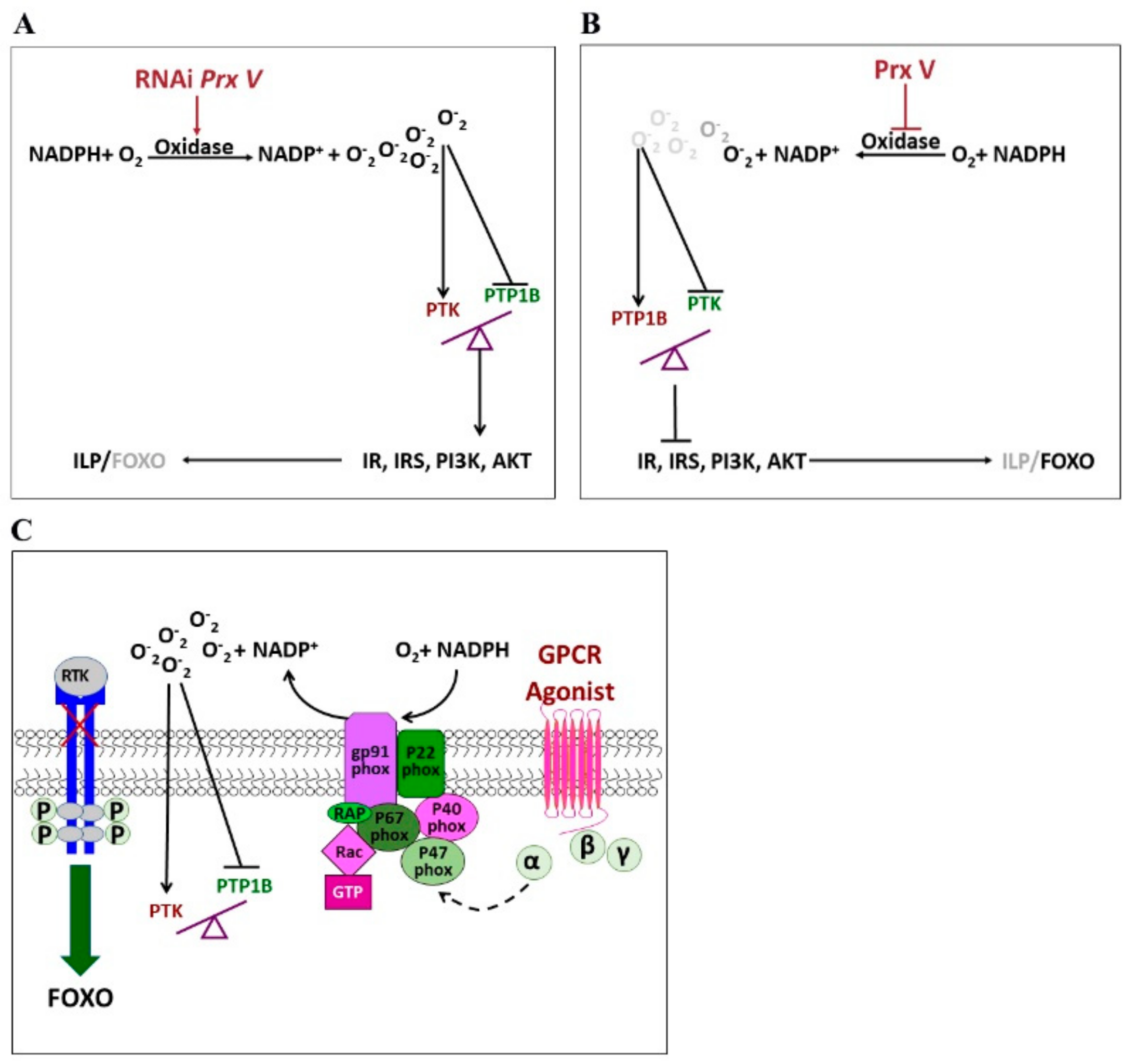
3.4. Antagonism between PTP1B and PTK Mediates in the ISP of Adults Regulates Egg Diapause in Migratory Locust
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| PTP1B | ProteinTyrosine Phosphatase-1B |
| PTK | Protein tyrosine kinase |
| PRX5 | Peroxiredoxin 5 |
| IR | Insulin receptor |
| IRS | Insulin receptor substrate |
| PI3K | Phosphoinositide 3-kinase |
| AKT | Serine/threonine-protein kinase |
Appendix A. Supporting Information Legends





Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (i) PRX V gene sequence |
| ATGCTGCCGATTTACTCTTTGACGGCCGTCAATACTACGAAGACTTTGTTGAAACAGTTTTGTAGATCGTTTCACGCAAGTAGCATCATGGGACTGAAGGTCGGCGACAAATTGCCATCG GTTGAACTGTATGAAAATAATCCTACCAATAAAGTTAATCTTGAAAAATTGCTGAGTAAC AAAAAGGCTGTCGTGTTCGCTGTACCTGGTGCATTTACACCTGGTTGTTCTAAGACACAC CTTCCAGGATACGTAGAAAAGGCAGACGAGTTGAAAAAGAAGGGAATTGATGAAATCATT TGTGTCTCCGTGAATGATCCATTTGTCATGGATGCGTGGGGAAAGGAGCACAAGGCGACC GGAAAGGTCCGGATGCTGGCTGATCCTGATGCATCATTCACAAAAGCATTGGAACTGGAA ACAAATTTACCACCTTTGGGGGGTACTCGTTCAAAACGGTATTCAATGATAATTGAAAAT GGAGTAGTTAAGTCATTAAATGTTGAGCCTGATGGCACAGGCCTATCATGTTCTTTGGCT GACAAACTTCCTTTGTAG |
| (ii) PTP1B gene sequence |
| ATGAGCAGGACTCCTGGTAAAAATAATATTGAAGCAGAATTTGTACAAATAGACTCGATT AGAGCGTGGACGCAACTGTATCAGCGCATAAGGGATGAGAGTTTAAACAATGATTATACA CTCATTGAAGCAAAGAAAGCAGAAAACAAGAATCTTAACAGGTATAGAGATGTCAGTCCA TATGATCATAGCAGAGTTGTTCTCACAAAAGGATCCTGCAACTATATCAATGCAAATCTTATCACGATGGAAAGGGCACATCGTCGGTATATCTTGACACAGGGCCCTTTGCCTCATACA ACAGGTCATTTCTGGTTGATGATTTGGGAACAGAAATGCAAAGCAGTGCTTATGCTAAAC AGAATAATTGAAAAGAACCAGGTTAAATGTCATCAGTACTGGCCTTCAGGTACAAAACCA GGTGTGGATGACAGACTGGAATTGAAAGATGTTGGCCTTGTTGTAGAGTTTGTCTCGGAG ACAGAAGCCTCATACTATACAACAAGAGTTCTGAGGTTGATAGATGAGGTGTCTGGAGAA AGCCGGCAGATTCTGCAGTTCCACTATACTACATGGCCTGACTTTGGTGTTCCAGAGAGC CCAACAGCATTCTTAAATTTCCTGACTGTAGTTAGGCAGAGTGGTGCCCTGGACCAGAAT GTTGGCCCTCCAGTAGTGCATTGCTCAGCTGGAATAGGGCGTTCGGGTACCTTCTGCCTT GTAGATTCTTGCCTTGTTCTTATTGAGGAGAATGGTCTTGACTCCGTGAATGTTGTGGAC GTGCTGCTGGAAATGAGGAAGTTTCGTATGGGCCTCATTCAGACGCCGGATCAGTTAAGA TTTTCTTACTTGGCAATTATTGAAGGTGCAAAGAAACTAATTAACAACAATCCTTTGCAT GACTACAATAGTGTAGAGGACACATCTCTGAATCATCATGATGGTTCAACAGAGGAGACA TCAGCAGATGAGGATGACACAGACGAACCACCGCCACTACCTCCGCCTCGAGGTGATTCG TTGACACGGTCTAAGATGGCTACAAACAATCATGGTATGAATGGGGGATTTGAAGCCAAC AAACCACTCCCTGTGGAACCTGAGGTCTCTGCAGAACAAGAGAGTGTGGAAAGATTGCTT GATTCAGCATTTCAAGATAAGGAAGCAGGTGATGATGATACTCCTCAGCTACGGCATCGA AGGCACGAGCGCCAGGAGCGGATATCAAGGATGAAAGGACATATTCGTGAAATCAAGCGC AAACAAGAAGCTGCTGAACGTTCTGAACAACTGTGGAGGTCCAGCAAGAAGAGGAGGAAGAGCAACACGGAAGAAGCAGGCTCCAGCTGTGACAGTGGTTCTAGTAAGCAGCAGTAA |
| (iii) PTK gene sequence |
| AAGGTGGTGGTCGCTCTTTACCCATTCAAAGCAATTGAAGGTGGTGACATATCATTGGAG AAGGGTGCTGAGTATGAGGTGCTTGACGATTCACAGGAACATTGGTGGAAGGTGAAGGAT GAGCATGGAAACATAGGTTACATCCCTAGTAACTATGTGAAGGAGAAAGAATTACTGGGA CTTCAAAAGTATGAATGGTACGTCGGAGACATGTCCAGGCAGAGAGCAGAGTCACTTCTA AAACAGGAAGACAAGGAAGGTTGCTTTGTGGTTCGGAATTCATCCACAAAAGGTCTCTAT ACACTCTCACTATATACCAAAGTTCCTCACCCCCATGTGAAACATTATCACATTAAGCAG AACTCTCGGGGAGAATTTTTCCTGTCTGAGAAGCACTGCTGCAGCTCCATCCCAGACCTG GTCAACTACCACCGCCACAACAGTGGCGGCCTCGCATCAAGGCTCAAGGCGAGCCCCTGT GAGCCCTGTTCCTGCCACTGCTGGCCTCAGCCATGATAAGTGGGAGATCGACCCACAA GAACTGATGTTGTTGGAAGAGCTCGGTTCTGGGCAGTTCGGTGTCGTGCGACGTGGCAAG TGGCGCGGATCCATAGATGTCGCTGTCAAGATGATGAAGCAGGGTACTATGTCAGAAGAT GACTTCATTGAGGAGGCGAAAGTCATGACACGACTGCAGCATCTCAACTTGGTCCAATTG TATGGTGTGTGCAGCAAGCACCGACCAATCTACATTGTTACGGAGTACATGCGCCACGGC TCTCTGCTCAACTACCTCCGCCGACACGAGGCCTCACTCGGTGGCAACACCGGCCTGCTG CTCGACATGTGCATACAAGTGTGCAATGGTATGGCCTACTTGGAGCGACACAACTATATT CACAGAGACCTGGCAGCACGAAATTGTCTGGTCGGTTCAGAAAATGTTGTGAAAGTTGCA GACTTTGGTTTAGCAAGGTACGTTTTGGATGACCAGTACACGAGCTCTGGCGGCACCAAG TTTCCAATTAAGTGGGCACCACCCGAGGTACTCAACTACACCAGGTTTTCCTCAAAGTCC GACGTCTGGGCTTATGGAGTTCTG |
| (iv) IR gene sequence |
| CATAGGCTTCAGCATTCTTGGGATATTGCTCATAATTGCTTTTATTTTTGTAAAGAGAAG GTATGAGAAGAAAGTTCAGCTGACTAAAAGATACATGGAGCATCTTGGCATTGATT GTATCAGCATCAAATCTGGGAACTCTTGACAAATGGGAGATTCCACGTGATAGAGTAGTCAT CAACCGGAAACTAGGAGAAGGAGCATTTGGTACTGTGTATGGAGGAGAAGCCCACTTTGCTGAGAAAGGATGGGTTGCAGTTGCTGTCAAAACATTAAAAGTT GGCTCTTCAACAGAAGAGAAGCTGGATTTCTTAAGTGAAGCAGAAGTTATGAAACGCTTTGAACATAAGAACATTGT AAAGCTGTTGGGTGTATGTACAAAAAATGAACCTGTCTACACTGTGATGGAATTCATGCTTATGGGGATCTGAAAACATTTCTTCTGGCTAGGCGACATTTAGTAAATGACAGACTTAC TGAAGAGTCAGATGAGGTGTCAAGCAAAAAGTTGACTTCGATGGCTCTGGATGTAGCAAG AGCTCTCAGTTACCTTGCTGATTTGAAATTTGTTCATCGTGATGTTGCATCACGGAACTG TCTGGTCAATGCTGCTCGTGTGGTGAAACTTGGAGATTTTGGAATGACACGACCCATGTT TGAAAATGATTACTACAAATTTAACAGGAAAGGTATGCTCCCTGTACGGTGGATGGCACC CGAGAGTTTGGAACAAAGAAGACATCACT TATGGCCGGACCACAAAGACCTTCTGTGGAACACCAGAGTACCTGGCACCTGAGGTCCTC GAGGACAACGACTACGGCCGCGCCGTCGACTGGTGGGGCATCGGCGTGGTCATGTATGAG ATGA |
References
- Yamashita, O. Diapause hormone of the silkworm, Bombyx mori: Structure, gene expression and function. J. Insect Physiol. 1996, 42, 669–679. [Google Scholar] [CrossRef]
- Yin, C.M.; Chippendale, G.M. Diapause of the southwestern corn borer, Diatraea grandiosella: Further evidence showing juvenile hormone to be the regulator. J. Insect Physiol. 1979, 25, 513–523. [Google Scholar] [CrossRef]
- Goto, S.G.; Denlinger, D.L. Genes encoding two cystatins in the flesh fly Sarcophaga crassipalpis and their distinct expression patterns in relation to pupal diapauses. Gene 2002, 292, 121–127. [Google Scholar] [CrossRef] [Green Version]
- Salminen, T.S.; Hoikkala, A. Effect of temperature on the duration of sensitive period and on the number of photoperiodic cycles required for the induction of reproductive diapause in Drosophila montana. J. Insect Physiol. 2013, 59, 450–457. [Google Scholar] [CrossRef]
- Denlinger, D.L. Regulation of diapause. Annu. Rev. Entomol. 2002, 47, 93–122. [Google Scholar] [CrossRef]
- Fukuda, S. The Production of Diapause Eggs by Transplanting the Subophageal Ganglion in the Silkworm. Proc. Jpn. Acad. B Phys. 1951, 27, 672–676. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, K. The Diapause Hormone of the Silkworm, Bombyx mori. Nature 1957, 179, 1300–1301. [Google Scholar] [CrossRef]
- De Reggi, M.; Gharib, B. Changes in ecdysteroid and juvenile hormone levels in developing eggs of Bombyx mori. J. Insect Physiol. 1983, 29, 871–876. [Google Scholar] [CrossRef]
- Denlinger, D.L. Hormonal Control of Diapause. In Comprehensive Insect Physiology, Biochemistry and Pharmacology; Kerkut, G.A., Gilbert, L.I., Eds.; Pergamon: Oxford, UK, 1985; Volume 8, pp. 353–412. [Google Scholar]
- Tatar, M.; Kopelman, A.; Tu, M.P.; Yin, C.M.; Garofalo, R.S. A mutant Drosophila insulin receptor homolog that extends life-span and impairs neuroendocrine function. Science 2001, 292, 107–110. [Google Scholar] [CrossRef] [Green Version]
- Williams, K.D.; Busto, M.; Suster, M.L.; So, A.K.C.; Ben-Shahar, Y.; Leevers, S.J.; Sokolowski, M.B. Natural variation in Drosophila melanogaster diapause due to the insulin-regulated PI3-kinase. Proc. Natl. Acad. Sci. USA 2006, 103, 15911–15915. [Google Scholar] [CrossRef] [Green Version]
- Antebi, A.; Culotti, J.G.; Hedgecock, E.M. daf-12 regulates developmental age and the dauer alternative in Caenorhabditis elegans. Development 1998, 125, 1191–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Apfeld, J.; Kenyon, C. Cell nonautonomy of C. elegans daf-2 function in the regulation of diapause and lifespan. Cell 1998, 95, 199–210. [Google Scholar] [CrossRef] [Green Version]
- Michaud, M.R.; Denlinger, D.L. Oleic acid is elevated in cell membranes during rapid cold-hardening and pupal diapause in the flesh fly, Sarcophaga crassipalpis. J. Insect Physiol. 2006, 52, 1073–1082. [Google Scholar] [CrossRef] [Green Version]
- Puig, O.; Marr, M.T.; Ruhf, M.L.; Tjian, R. Control of cell number by Drosophila FOXO: Downstream and feedback regulation of the insulin receptor pathway. Genes Dev. 2003, 17, 2006–2020. [Google Scholar] [CrossRef] [Green Version]
- Hahn, D.A.; Denlinger, D.L. Meeting the energetic demands of insect diapause: Nutrient storage and utilization. J. Insect Physiol. 2007, 53, 760–763. [Google Scholar] [CrossRef]
- Sim, C.; Denlinger, D.L. Insulin Signaling and FOXO Regulate the Overwintering Diapause of the Mosquito Culex pipiens. Proc. Natl. Acad. Sci. USA 2008, 105, 6777–6781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gronke, S.; Clarke, D.F.; Broughton, S.; Andrews, T.D.; Partridge, L. Molecular evolution and functional characterization of Drosophila insulin-like peptides. PLoS Genet. 2010, 6, e1000857. [Google Scholar] [CrossRef] [Green Version]
- Ruan, Y.; Chen, C.; Cao, Y.; Garofalo, R.S. The Drosophila insulin receptor contains a novel carboxyl-terminal extension likely to play an important role in signal transduction. J. Biol. Chem. 1995, 270, 4236–4243. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, T.; Fernandez, R.; Roth, R.A. Comparison of the signaling abilities of the drosophila and human insulin receptors in mammalian cells. Biochemistry 1995, 34, 4962–4968. [Google Scholar] [CrossRef]
- Sasaoka, T.; Rose, D.W.; Jhun, B.H.; Saltiel, A.R.; Draznin, B.; Olefsky, J.M. Evidence for a functional role of Shc proteins in mitogenic signaling induced by insulin, insulin-like growth factor-1, and epidermal growth factor. J. Biol. Chem. 1994, 269, 13689–13694. [Google Scholar] [CrossRef]
- Fernandez, R.; Tabarini, D.; Azpiazu, N.; Frasch, M.; Schlessinger, J. The Drosophila insulin receptor homolog: A gene essential for embryonic development encodes two receptor isoforms with different signaling potential. EMBO J. 1995, 14, 3373–3384. [Google Scholar] [CrossRef] [PubMed]
- Fullbright, G.; Lacy, E.R.; Büllesbach, E.E. The Prothoracicotropic Hormone bombyxin has specific receptors on insect ovarian cells. Eur. J. Org. Chem. 1997, 245, 774–780. [Google Scholar] [CrossRef]
- Adrain, C.; Freeman, M. Regulation of receptor tyrosine kinase ligand processing. Cold Spring Harb. Perspect. Biol. 2014, 6, a008995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, N.; Konidaris, K.F.; Gasser, G.; Tonks, N.K. A potent, selective, and orally bioavailable inhibitor of the protein-tyrosine phosphatase PTP1B improves insulin and leptin signaling in animal models. J. Biol. Chem. 2018, 293, 1517–1525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egawa, K.; Maegawa, H.; Shimizu, S.; Morino, K.; Nishio, Y.; Bryer-Ash, M.; Cheung, A.T.; Kolls, J.K.; Kikkawa, R.; Kashiwagi, A. Protein-tyrosine phosphatase-1B negatively regulates insulin signaling in l6 myocytes and Fao hepatoma cells. J. Biol. Chem. 2001, 276, 10207–10211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonks, N.K. Protein tyrosine phosphatases: From genes, to function, to disease. Nat. Rev. Mol. Cell Biol. 2006, 7, 833–846. [Google Scholar] [CrossRef]
- Schlessinger, J. Cell signaling by receptor tyrosine kinases. Cell 2010, 141, 1117–1134. [Google Scholar] [CrossRef] [Green Version]
- Ding, D.; Liu, G.J.; Hou, L.; Gui, W.Y.; Chen, B.; Kang, L. Genetic variation in PTPN1 contributes to metabolic adaptation to high-altitude hypoxia in Tibetan migratory locusts. Nat. Commun. 2018, 9, 4991. [Google Scholar] [CrossRef]
- Tonks, N.K. Redox redux: Revisiting PTPs and the control of cell signaling. Cell 2005, 121, 667–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, D.W.; Baek, K.; Kim, J.R.; Gui, W.Y.; Chen, B.; Kang, L. Resveratrol inhibits foam cell formation via NADPH oxidase 1-mediated reactive oxygen species and monocyte chemotactic protein-1. Exp. Mol. Med. 2009, 41, 171–179. [Google Scholar] [CrossRef]
- Tu, X.B.; Wang, J.; Hao, K.; Whitman, D.W.; Fan, Y.L.; Cao, G.C.; Zhang, Z.H. Transcriptomic and proteomic analysis of pre-diapause and non-diapause eggs of migratory locust, Locusta migratoria L. (Orthoptera: Acridoidea). Sci. Rep. 2015, 5, 11402. [Google Scholar] [CrossRef] [Green Version]
- Yu, L.; Liu, J.P.; Zhuang, Z.X.; Yang, L.Q.; Zhang, R.L.; Ye, X.M.; Cheng, J.Q. Quantitative analysis of real-time PCR expression production by REST and 2~((−ΔΔCT)). J. Trop. Med. 2007, 10, 956–958. (In Chinese) [Google Scholar]
- Radonić, A.; Thulke, S.; Mackay, I.M.; Landt, O.; Siegert, W.; Nitsche, A. Guideline to reference gene selection for quantitative real-time PCR. Biochem. Biophys. Res. 2004, 313, 856–862. [Google Scholar] [CrossRef]
- Hao, K.; Tu, X.B.; Ullah, H.; McNeill, M.R.; Zhang, Z.Z. Novel Lom-dh genes play potential role in promoting egg diapause of Locusta migratoria L. Front. Physiol. 2019, 10, 767. [Google Scholar] [CrossRef] [PubMed]
- Hao, K.; Jarwar, A.R.; Ullah, H.; Tu, X.B.; Nong, X.Q.; Zhang, Z.Z. Transcriptome sequencing reveals potential mechanisms of the maternal effect on egg diapause induction of Locusta migratoria. Int. J. Mol. Sci. 2019, 20, 1974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Zhang, Z.Y. PTP1B as a drug target: Recent developments in PTP1B inhibitor discovery. Drug Discov. Today 2007, 12, 373–381. [Google Scholar] [CrossRef]
- Carlo-Stella, C.; Regazzi, E.; Garau, D.; Mangoni, L.; Rizzo, M.T. Effect of the protein tyrosine kinase inhibitor genistein on normal and leukaemic haemopoietic progenitor cells. Br. J. Haematol. 1996, 93, 551–557. [Google Scholar] [CrossRef] [PubMed]
- Cregg, J.M.; Cereghino, J.L.; Shi, J.; Higgins, D.R. Recombinant protein expression in Pichia pastoris. Mol. Biotech. 2000, 16, 23–52. [Google Scholar] [CrossRef]
- Hawkins, E.; Smith, W.; O’Donnell, M.J. Measurements of ooplasmic pCa in fertilized eggs of the locust, Locusta migratoria, and a role for external calcium in experimental activation of unfertilized eggs. Invertebr. Reprod. Dev. 1998, 34, 165–172. [Google Scholar] [CrossRef]
- Cattaneo, F.; Guerra, G.; Parisi, M.; Marinis, M.D.; Tafuri, D.; Cinelli, M.; Ammendola, R. Cell-surface receptors transactivation mediated by G protein-coupled receptors. Int. J. Mol. Sci. 2014, 15, 19700–19728. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.L. The Main Locust and Ecological Management of Locust Plague in China; Science Press: Beijing, China, 2007; pp. 94–101. [Google Scholar]
- Hao, K.; Wang, J.; Tu, X.B.; Whitman, D.W.; Zhang, Z.H. Transcriptomic and proteomic of Locusta migratoria eggs at different embryonic states: Comparison for diapause and non-diapause regimes. J. Integr. Agric. 2017, 16, 1777–1788. [Google Scholar] [CrossRef] [Green Version]
- Mustelin, T.; Abraham, R.T.; Rudd, C.E.; Alonso, A.; Merlo, J.J. Protein tyrosine phosphorylation in T cell signaling. Front. Biosci. 2002, 7, 918–969. [Google Scholar] [CrossRef]
- Mustelin, T.; Taskén, K. Positive and negative regulation of T-cell activation through kinases and phosphatases. Biochem. J. 2003, 371, 15–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Li, L.; Qian, Z.; Hong, J.; Shen, S.; Huang, W. Reduction of PTP1B by RNAi upregulates the activity of insulin controlled fatty acid synthase promoter. Biochem. Biophys. Res. Commun. 2005, 329, 538–543. [Google Scholar] [CrossRef] [PubMed]
- Homma, T.; Watanabe, K.; Tsurumaru, S.; Kataoka, H.; Imai, K.; Kamba, M.; Niimi, T.; Yamashita, O.; Yaginuma, T. G protein-coupled receptor for diapause hormone, an inducer of Bombyx Embryonic diapause. Biochem. Biophys. Res. Commun. 2006, 344, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Sumimoto, H. Structure, regulation and evolution of Nox-family NADPH oxidases that produce reactive oxygen species. FEBS J. 2008, 275, 3249–3277. [Google Scholar] [CrossRef] [PubMed]
- Putker, M.; Madl, T.; Vos, H.R.; de Ruiter, H.; Visscher, M.; van den Berg, M.C.; Kaplan, M.; Korswagen, H.; Boelens, R.; Vermeulen, M. Redox-dependent control of FOXO/DAF-16 by transportin-1. Mol. Cell. 2013, 49, 730–742. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.S.; Wang, T.; Lin, X.W.; Denlinger, D.L.; Xu, W.H. Reactive oxygen species extend insect life span using components of the insulin-signaling pathway. Proc. Natl. Acad. Sci. USA 2017, 14, 7832–7840. [Google Scholar] [CrossRef] [Green Version]
- Balaban, R.S.; Nemoto, S.; Finkel, T. Mitochondria, oxidants, and aging. Cell 2005, 120, 483–495. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.; Ma, R.; Ding, D.; Wei, L.; Kang, L. Aerobic respiration by haemocyanin in the embryo of the migratory locust. Insect Mol. Biol. 2017, 26, 461–468. [Google Scholar] [CrossRef]
- Sim, C.; Denlinger, D.L. Insulin signaling and the regulation of insect diapause. Front. Physiol. 2013, 4, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Nachman, R.J.; Denlinger, D.L. Diapause hormone in the helicoverpa/heliothis complex: A review of gene expression, peptide structure and activity, analog and antagonist development, and the receptor. Peptides 2015, 72, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Gerisch, B.; Weitzel, C.; Kober-Eisermann, C.; Rottiers, V.; Antebi, A. A hormonal signaling pathway influencing C. elegans metabolism, reproductive development, and life span. Dev. Cell. 2001, 1, 841–851. [Google Scholar] [CrossRef] [Green Version]
- Saito, H.; Takeuchi, Y.; Takeda, R.; Hayashi, Y.; Watanabe, K.; Shin, M.; Imai, K.; Isobe, M.; Yamashita, O. The core and complementary sequence responsible for biological activity of the diapause hormone of the silkworm. Bombyx Mori. Peptides 1994, 15, 1173–1178. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Wang, J.; Cui, D.; Hao, K.; Chen, J.; Tu, X.; Zhang, Z. Antagonism between PTP1B and PTK Mediates Adults’ Insulin-Like Signaling Regulation of Egg Diapause in the Migratory Locust. Insects 2021, 12, 253. https://doi.org/10.3390/insects12030253
Li S, Wang J, Cui D, Hao K, Chen J, Tu X, Zhang Z. Antagonism between PTP1B and PTK Mediates Adults’ Insulin-Like Signaling Regulation of Egg Diapause in the Migratory Locust. Insects. 2021; 12(3):253. https://doi.org/10.3390/insects12030253
Chicago/Turabian StyleLi, Shuang, Jie Wang, Dongnan Cui, Kun Hao, Jun Chen, Xiongbing Tu, and Zehua Zhang. 2021. "Antagonism between PTP1B and PTK Mediates Adults’ Insulin-Like Signaling Regulation of Egg Diapause in the Migratory Locust" Insects 12, no. 3: 253. https://doi.org/10.3390/insects12030253
APA StyleLi, S., Wang, J., Cui, D., Hao, K., Chen, J., Tu, X., & Zhang, Z. (2021). Antagonism between PTP1B and PTK Mediates Adults’ Insulin-Like Signaling Regulation of Egg Diapause in the Migratory Locust. Insects, 12(3), 253. https://doi.org/10.3390/insects12030253