
A Method for Repeated, Longitudinal Sampling of Individual Aedes aegypti for Transmission Potential of Arboviruses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
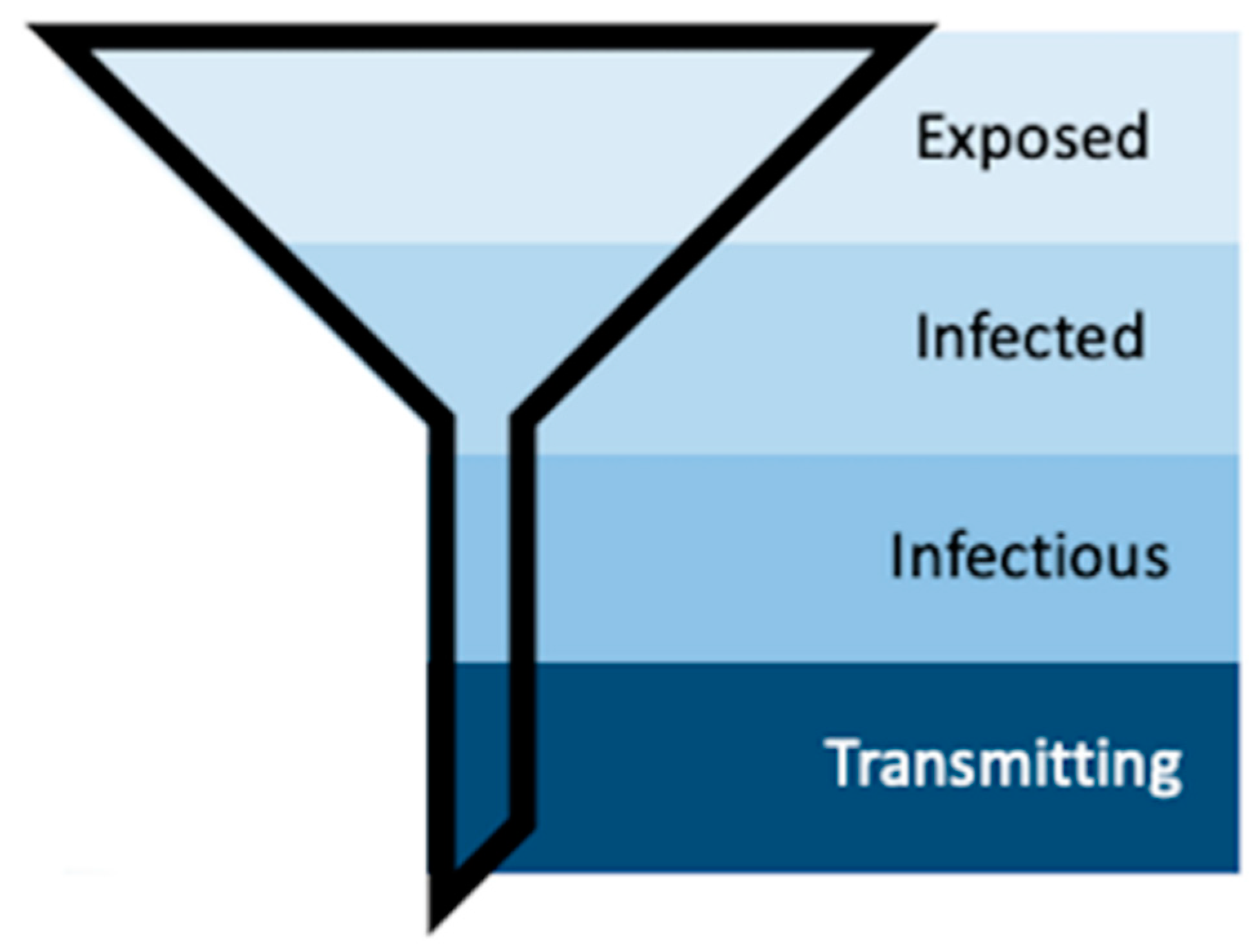
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Virus
2.2. Viral Quantification and Testing
2.3. Mosquito Exposure and Maintenance
2.4. Traditional Vector Competence Assay
2.5. Limit of Detection
2.6. Longitudinal Sampling
3. Results
3.1. Model System Development
3.1.1. In Vitro Limit of Detection
3.1.2. Vector Competence by Traditional Measures
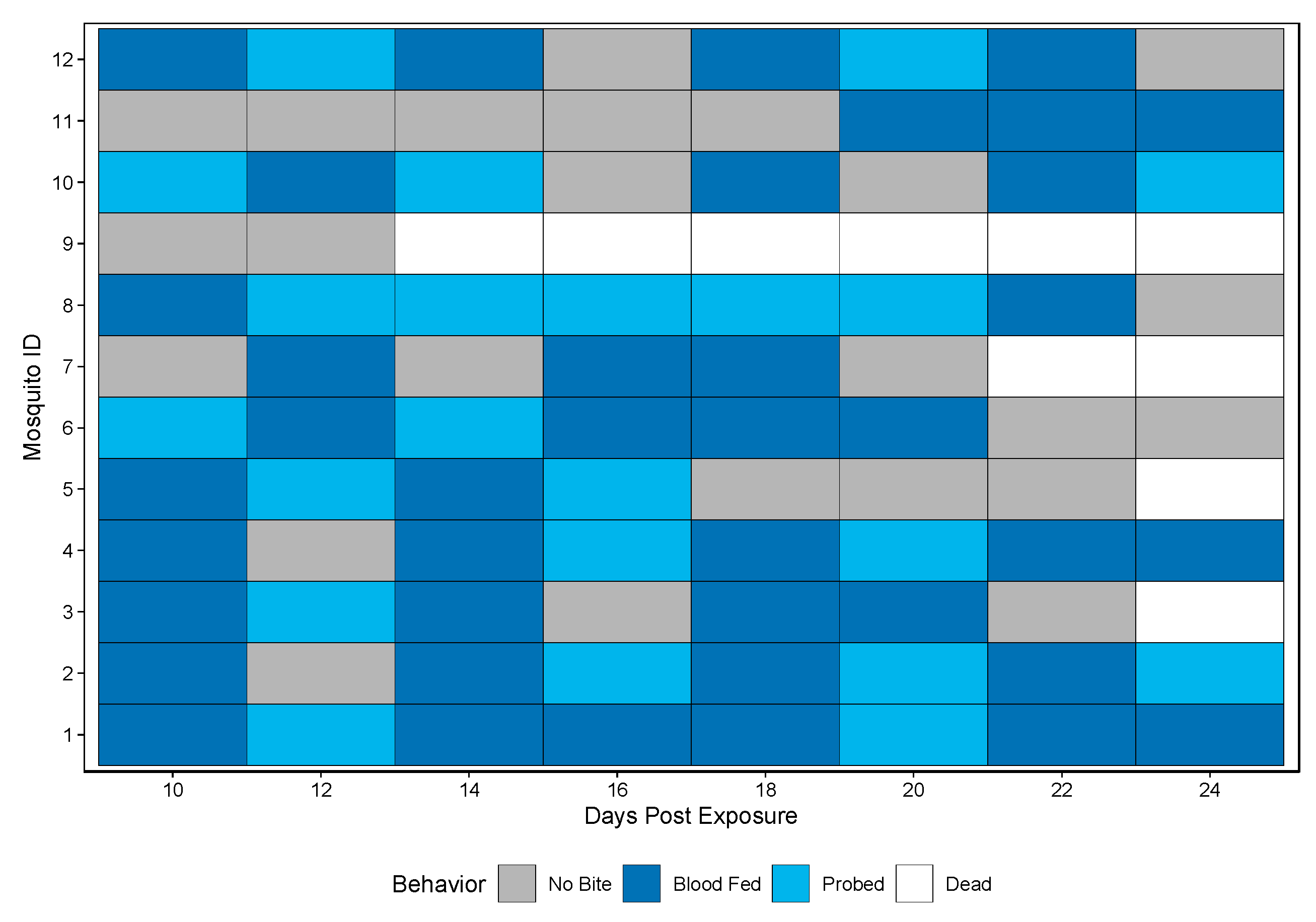
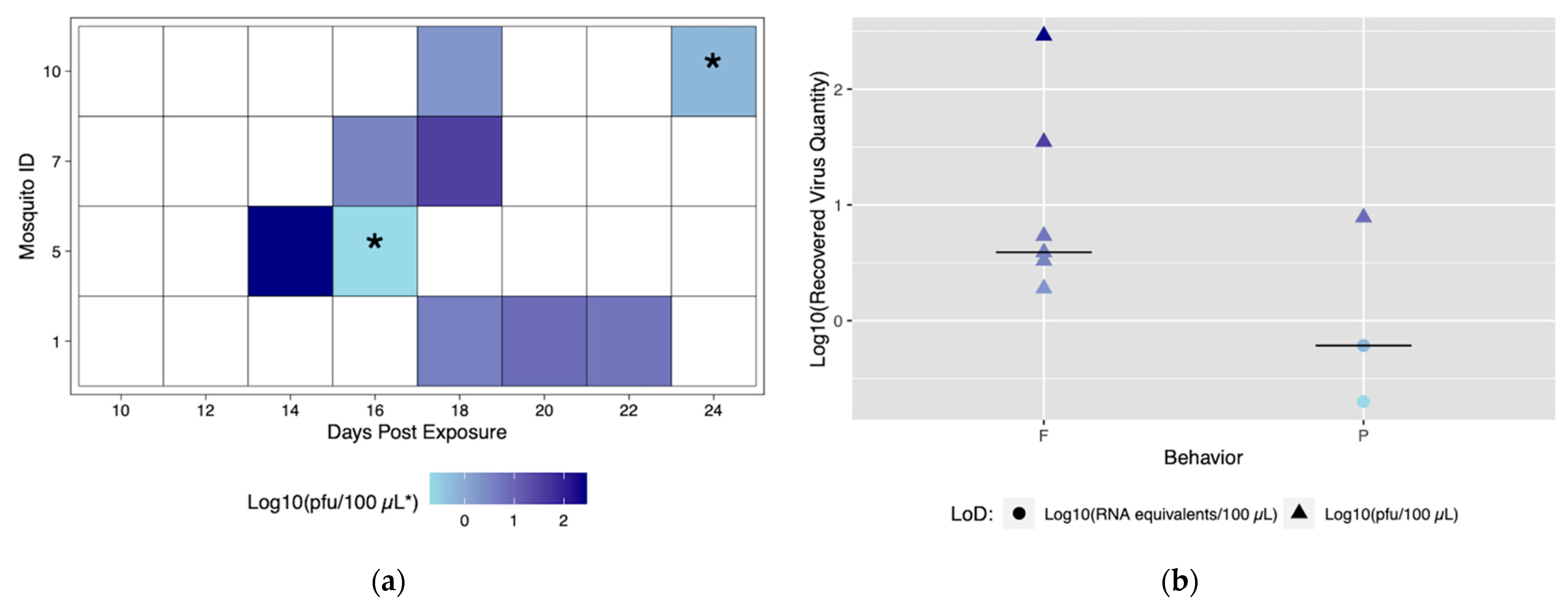
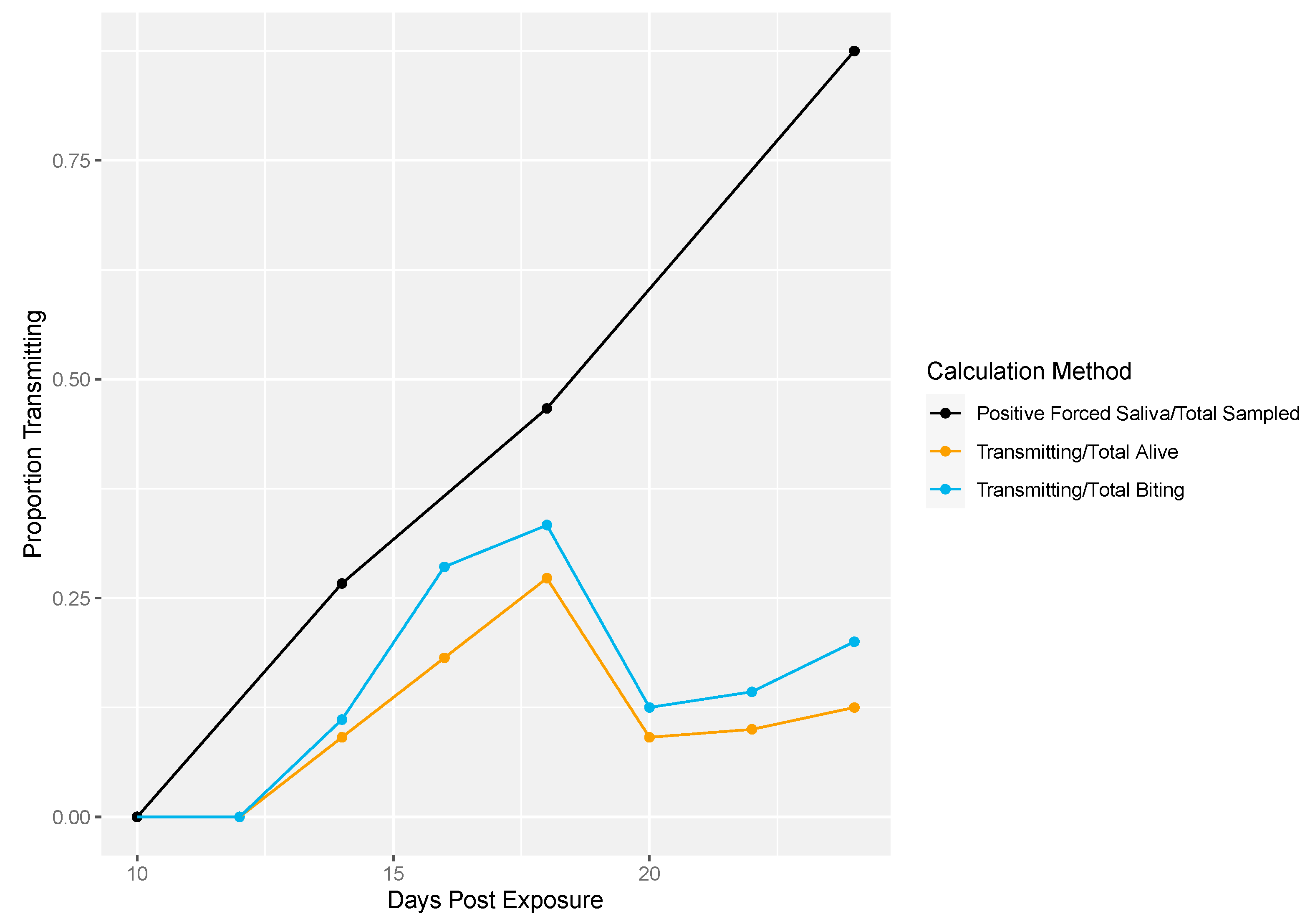
3.1.3. Individual, Longitudinal Vector Competence Method
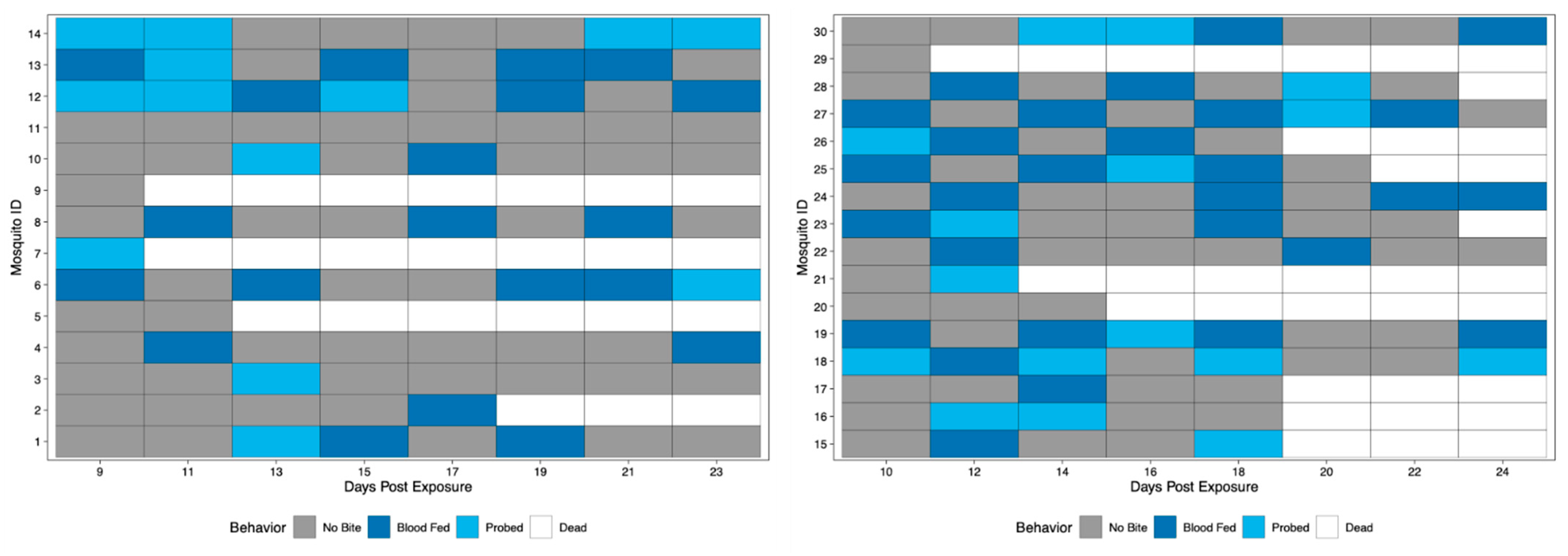
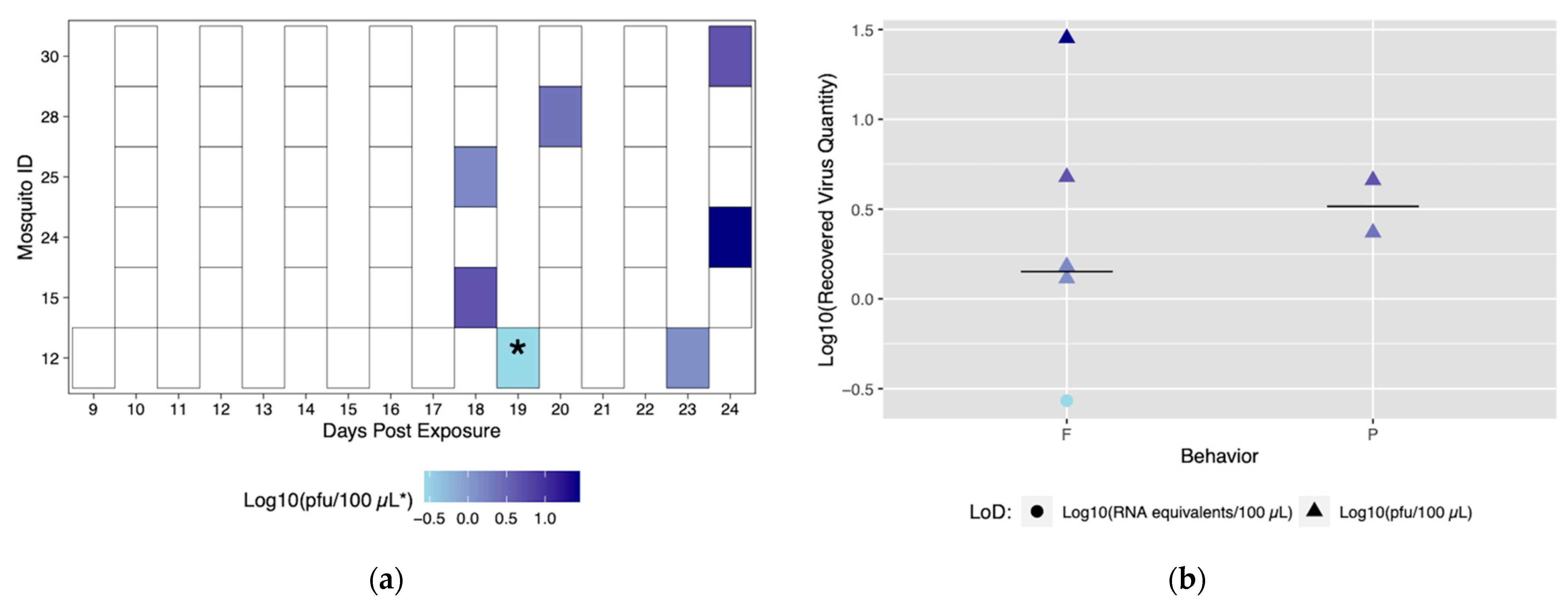
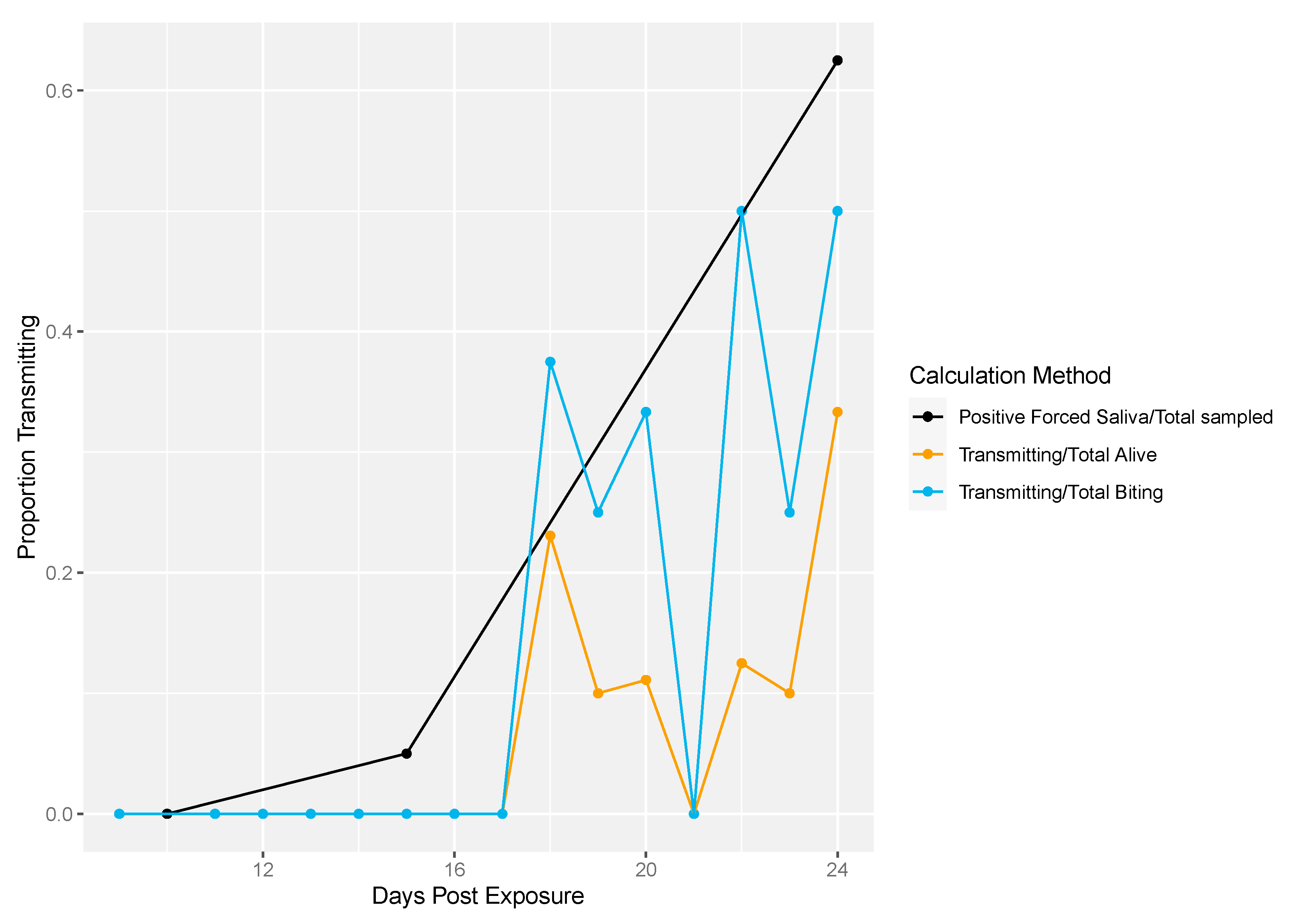
3.2. Application of Method to Field-Derived Mosquitoes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pietsch, C.; Michalski, D.; Munch, J.; Petros, S.; Bergs, S.; Trawinski, H.; Lubbert, C.; Liebert, U.G. Autochthonous West Nile virus infection outbreak in humans, Leipzig, Germany, August to September 2020. Eurosurveillance 2020, 25, 2001786. [Google Scholar] [CrossRef]
- Aubry, F.; Dabo, S.; Manet, C.; Filipovic, I.; Rose, N.H.; Miot, E.F.; Martynow, D.; Baidaliuk, A.; Merkling, S.H.; Dickson, L.B.; et al. Enhanced Zika virus susceptibility of globally invasive Aedes aegypti populations. Science 2020, 370, 991–996. [Google Scholar] [CrossRef]
- Vermeulen, T.D.; Reimerink, J.; Reusken, C.; Giron, S.; de Vries, P.J. Autochthonous dengue in two Dutch tourists visiting Departement Var, southern France, July 2020. Eurosurveillance 2020, 25, 2001670. [Google Scholar] [CrossRef]
- Brady, O.J.; Johansson, M.A.; Guerra, C.A.; Bhatt, S.; Golding, N.; Pigott, D.M.; Delatte, H.; Grech, M.G.; Leisnham, P.T.; Maciel-de-Freitas, R.; et al. Modelling adult Aedes aegypti and Aedes albopictus survival at different temperatures in laboratory and field settings. Parasites Vectors 2013, 6, 351. [Google Scholar] [CrossRef] [Green Version]
- Kraemer, M.U.G.; Reiner, R.C., Jr.; Brady, O.J.; Messina, J.P.; Gilbert, M.; Pigott, D.M.; Yi, D.; Johnson, K.; Earl, L.; Marczak, L.B.; et al. Past and future spread of the arbovirus vectors Aedes aegypti and Aedes albopictus. Nat. Microbiol. 2019, 4, 854–863. [Google Scholar] [CrossRef]
- Messina, J.P.; Brady, O.J.; Scott, T.W.; Zou, C.; Pigott, D.M.; Duda, K.A.; Bhatt, S.; Katzelnick, L.; Howes, R.E.; Battle, K.E.; et al. Global spread of dengue virus types: Mapping the 70 year history. Trends Microbiol. 2014, 22, 138–146. [Google Scholar] [CrossRef] [Green Version]
- Kraemer, M.U.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. Elife 2015, 4, e08347. [Google Scholar] [CrossRef]
- Lounibos, L.P.; Kramer, L.D. Invasiveness of Aedes aegypti and Aedes albopictus and Vectorial Capacity for Chikungunya Virus. J. Infect. Dis. 2016, 214, S453–S458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valderrama, A.; Diaz, Y.; Lopez-Verges, S. Interaction of Flavivirus with their mosquito vectors and their impact on the human health in the Americas. Biochem. Biophys. Res. Commun. 2017, 492, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Dick, O.B.; San Martin, J.L.; Montoya, R.H.; del Diego, J.; Zambrano, B.; Dayan, G.H. The history of dengue outbreaks in the Americas. Am. J. Trop. Med. Hyg. 2012, 87, 584–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marano, G.; Pupella, S.; Pati, I.; Masiello, F.; Franchini, M.; Vaglio, S.; Velati, C.; Liumbruno, G.M. Ten years since the last Chikungunya virus outbreak in Italy: History repeats itself. High Speed Blood Transfus. Equip. 2017, 15, 489–490. [Google Scholar]
- Braack, L.; de Almeida, A.P.G.; Cornel, A.J.; Swanepoel, R.; de Jager, C. Mosquito-borne arboviruses of African origin: Review of key viruses and vectors. Parasites Vectors 2018, 11, 29. [Google Scholar] [CrossRef]
- Dengue/DHF Update (12): Americas, in ProMED-Mail. 2020. Available online: www.promedmail.org (accessed on 30 November 2020).
- Campos, M.; Ward, D.; Morales, R.F.; Gomes, A.R.; Silva, K.; Sepulveda, N.; Gomez, L.F.; Clark, T.G.; Campino, S. Surveillance of Aedes aegypti populations in the city of Praia, Cape Verde: Zika virus infection, insecticide resistance and genetic diversity. Parasites Vectors 2020, 13, 481. [Google Scholar] [CrossRef]
- Ciota, A.T.; Kramer, L.D. Vector-virus interactions and transmission dynamics of West Nile virus. Viruses 2013, 5, 3021–3047. [Google Scholar] [CrossRef]
- Kramer, L.D.; Ciota, A.T. Dissecting vectorial capacity for mosquito-borne viruses. Curr. Opin. Virol. 2015, 15, 112–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christofferson, R.C.; Mores, C.N. Estimating the magnitude and direction of altered arbovirus transmission due to viral phenotype. PLoS ONE 2011, 6, e16298. [Google Scholar] [CrossRef] [PubMed]
- Richardson, J.; Molina-Cruz, A.; Salazar, M.I.; Black, W. Quantitative analysis of dengue-2 virus RNA during the extrinsic incubation period in individual Aedes aegypti. Am. J. Trop. Med. Hyg. 2006, 74, 132–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrett-Jones, C. Prognosis for Interruption of Malaria Transmission through Assessment of the Mosquito’s Vectorial Capacity. Nature 1964, 204, 1173–1175. [Google Scholar] [CrossRef]
- Smith, D.R.; Carrara, A.S.; Aguilar, P.V.; Weaver, S.C. Evaluation of methods to assess transmission potential of Venezuelan equine encephalitis virus by mosquitoes and estimation of mosquito saliva titers. Am. J. Trop. Med. Hyg. 2005, 73, 33–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christofferson, R.C.; Mores, C.N.; Wearing, H.J. Bridging the Gap Between Experimental Data and Model Parameterization for Chikungunya Virus Transmission Predictions. J. Infect. Dis. 2016, 214, S466–S470. [Google Scholar] [CrossRef] [Green Version]
- Bellan, S.E. The importance of age dependent mortality and the extrinsic incubation period in models of mosquito-borne disease transmission and control. PLoS ONE 2010, 5, e10165. [Google Scholar] [CrossRef]
- Mayton, E.H.; Tramonte, A.R.; Wearing, H.J.; Christofferson, R.C. Age-structured vectorial capacity reveals timing, not magnitude of within-mosquito dynamics is critical for arbovirus fitness assessment. Parasites Vectors 2020, 13, 310. [Google Scholar] [CrossRef]
- Christofferson, R.C.; Chisenhall, D.M.; Wearing, H.J.; Mores, C.N. Chikungunya viral fitness measures within the vector and subsequent transmission potential. PLoS ONE 2014, 9, e110538. [Google Scholar] [CrossRef] [Green Version]
- Christofferson, R.C.; Mores, C.N.; Wearing, H.J. Characterizing the likelihood of dengue emergence and detection in naive populations. Parasites Vectors 2014, 7, 282. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, P.M.; Ehrlich, H.Y.; Magalhaes, T.; Miller, M.R.; Conway, P.J.; Bransfield, A.; Misencik, M.J.; Gloria-Soria, A.; Warren, J.L.; Andreadis, T.G.; et al. Successive blood meals enhance virus dissemination within mosquitoes and increase transmission potential. Nat. Microbiol. 2020, 5, 239–247. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.L.; Houk, E.J.; Kramer, L.D.; Reeves, W.C. Intrinsic factors affecting vector competence of mosquitoes for arboviruses. Annu. Rev. Entomol. 1983, 28, 229–262. [Google Scholar] [CrossRef] [PubMed]
- Cator, L.J.; Johnson, L.R.; Mordecai, E.A.; Moustaid, F.E.; Smallwood, T.R.C.; LaDeau, S.L.; Johansson, M.A.; Hudson, P.J.; Boots, M.; Thomas, M.B.; et al. The Role of Vector Trait Variation in Vector-Borne Disease Dynamics. Front. Ecol. Evol. 2020, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Kawiecki, A.B.; Mayton, E.H.; Dutuze, M.F.; Goupil, B.A.; Langohr, I.M.; Del Piero, F.; Christofferson, R.C. Tissue tropisms, infection kinetics, histologic lesions, and antibody response of the MR766 strain of Zika virus in a murine model. Virol. J. 2017, 14, 82. [Google Scholar] [CrossRef] [PubMed]
- Weger-Lucarelli, J.; Ruckert, C.; Chotiwan, N.; Nguyen, C.; Luna, S.M.G.; Fauver, J.R.; Foy, B.D.; Perera, R.; Black, W.C.; Kading, R.C.; et al. Vector Competence of American Mosquitoes for Three Strains of Zika Virus. PLoS Negl. Trop. Dis. 2016, 10, e0005101. [Google Scholar] [CrossRef] [Green Version]
- Ciota, A.T.; Bialosuknia, S.M.; Zink, S.D.; Brecher, M.; Ehrbar, D.J.; Morrissette, M.N.; Kramer, L.D. Effects of Zika Virus Strain and Aedes Mosquito Species on Vector Competence. Emerg. Infect. Dis. 2017, 23, 1110–1117. [Google Scholar] [CrossRef] [Green Version]
- Faye, O.; Faye, O.; Dupressoir, A.; Weidmann, M.; Ndiaye, M.; Sall, A.A. One-step RT-PCR for detection of Zika virus. J. Clin. Virol. 2008, 43, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Tramonte, A.R.; Christofferson, R.C. Investigating the probability of establishment of Zika virus and detection through mosquito surveillance under different temperature conditions. PLoS ONE 2019, 14, e0214306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tesla, B.; Demakovsky, L.R.; Mordecai, E.A.; Ryan, S.J.; Bonds, M.H.; Ngonghala, C.N.; Brindley, M.A.; Murdock, C.C. Temperature drives Zika virus transmission: Evidence from empirical and mathematical models. Proc. R. Soc. B 2018, 285, 20180795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, T.W.; Clark, G.G.; Lorenz, L.H.; Amerasinghe, P.H.; Reiter, P.; Edman, J.D. Detection of multiple blood feeding in Aedes aegypti (Diptera: Culicidae) during a single gonotrophic cycle using a histologic technique. J. Med. Entomol. 1993, 30, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Cebrián-Camisón, S.; La Puente, J.M.-D.; Figuerola, J. A Literature Review of Host Feeding Patterns of Invasive Aedes Mosquitoes in Europe. Insects 2020, 11, 848. [Google Scholar] [CrossRef] [PubMed]
- Scott, T.W.; Chow, E.; Strickman, D.; Kittayapong, P.; Wirtz, R.A.; Lorenz, L.H.; Edman, J.D. Blood-feeding patterns of Aedes aegypti (Diptera: Culicidae) collected in a rural Thai village. J. Med. Entomol. 1993, 30, 922–927. [Google Scholar] [CrossRef] [PubMed]
- Stoddard, S.T.; Forshey, B.M.; Morrison, A.C.; Paz-Soldan, V.A.; Vazquez-Prokopec, G.M.; Astete, H.; Reiner, R.C., Jr.; Vilcarromero, S.; Elder, J.P.; Halsey, E.S.; et al. House-to-house human movement drives dengue virus transmission. Proc. Natl. Acad. Sci. USA 2013, 110, 994–999. [Google Scholar] [CrossRef] [Green Version]
- Styer, L.M.; Bernard, K.A.; Kramer, L.D. Enhanced early West Nile virus infection in young chickens infected by mosquito bite: Effect of viral dose. Am. J. Trop. Med. Hyg. 2006, 75, 337–345. [Google Scholar] [CrossRef]
- Ross, P.A.; Endersby-Harshman, N.M.; Hoffmann, A.A. A comprehensive assessment of inbreeding and laboratory adaptation in Aedes aegypti mosquitoes. Evol. Appl. 2019, 12, 572–586. [Google Scholar] [CrossRef] [Green Version]
- Hol, F.J.; Lambrechts, L.; Prakash, M. BiteOscope, an open platform to study mosquito biting behavior. Elife 2020, 9, 9. [Google Scholar] [CrossRef]
- Kumar, S.; Hol, F.J.; Pujhari, S.; Ellington, C.; Narayanan, H.V.; Li, H.; Rasgon, J.L.; Prakash, M. Vectorchip: Microfluidic platform for highly parallel bite by bite profiling of mosquito-borne pathogen transmission. bioRxiv 2020. [Google Scholar] [CrossRef]
- Zermoglio, P.F.; Robuchon, E.; Leonardi, M.S.; Chandre, F.; Lazzari, C.R. What does heat tell a mosquito? Characterization of the orientation behaviour of Aedes aegypti towards heat sources. J. Insect Physiol. 2017, 100, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Dekker, T.; Carde, R.T. Moment-to-moment flight manoeuvres of the female yellow fever mosquito (Aedes aegypti L.) in response to plumes of carbon dioxide and human skin odour. J. Exp. Biol. 2011, 214, 3480–3494. [Google Scholar] [CrossRef] [Green Version]
- Healy, T.P.; Copland, M.J.W.; Cork, A.; Przyborowska, A.; Halket, J.M. Landing responses of Anopheles gambiae elicited by oxocarboxylic acids. Med. Vet. Entomol. 2002, 16, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Aubry, F.; Jacobs, S.; Darmuzey, M.; Lequime, S.; Delang, L.; Fontaine, A.; Jupatanakul, N.; Miot, E.F.; Dabo, S.; Manet, C.; et al. Recent African strains of Zika virus display higher transmissibility and fetal pathogenicity than Asian strains. Nat. Commun. 2021, 12, 916. [Google Scholar] [CrossRef]
- Azar, S.R.; Weaver, S.C. Vector Competence: What Has Zika Virus Taught Us? Viruses 2019, 11, 867. [Google Scholar] [CrossRef] [Green Version]
- Alto, B.W.; Wiggins, K.; Eastmond, B.; Ortiz, S.; Zirbel, K.; Lounibos, L.P. Diurnal Temperature Range and Chikungunya Virus Infection in Invasive Mosquito Vectors. J. Med. Entomol. 2018, 55, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Christofferson, R.C.; Mores, C.N. Potential for Extrinsic Incubation Temperature to Alter Interplay Between Transmission Potential and Mortality of Dengue-Infected Aedes aegypti. Environ. Health Insights 2016, 10, 119–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muttis, E.; Balsalobre, A.; Chuchuy, A.; Mangudo, C.; Ciota, A.T.; Kramer, L.D.; Micieli, M.V. Factors Related to Aedes aegypti (Diptera: Culicidae) Populations and Temperature Determine Differences on Life-History Traits with Regional Implications in Disease Transmission. J. Med. Entomol. 2018, 55, 1105–1112. [Google Scholar] [CrossRef]
- Danet, L.; Beauclair, G.; Berthet, M.; Moratorio, G.; Gracias, S.; Tangy, F.; Choumet, V.; Jouvenet, N. Midgut barriers prevent the replication and dissemination of the yellow fever vaccine in Aedes aegypti. PLoS Negl. Trop. Dis. 2019, 13, e0007299. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayton, E.H.; Hernandez, H.M.; Vitek, C.J.; Christofferson, R.C. A Method for Repeated, Longitudinal Sampling of Individual Aedes aegypti for Transmission Potential of Arboviruses. Insects 2021, 12, 292. https://doi.org/10.3390/insects12040292
Mayton EH, Hernandez HM, Vitek CJ, Christofferson RC. A Method for Repeated, Longitudinal Sampling of Individual Aedes aegypti for Transmission Potential of Arboviruses. Insects. 2021; 12(4):292. https://doi.org/10.3390/insects12040292
Chicago/Turabian StyleMayton, E. Handly, Heather M. Hernandez, Christopher J. Vitek, and Rebecca C. Christofferson. 2021. "A Method for Repeated, Longitudinal Sampling of Individual Aedes aegypti for Transmission Potential of Arboviruses" Insects 12, no. 4: 292. https://doi.org/10.3390/insects12040292
APA StyleMayton, E. H., Hernandez, H. M., Vitek, C. J., & Christofferson, R. C. (2021). A Method for Repeated, Longitudinal Sampling of Individual Aedes aegypti for Transmission Potential of Arboviruses. Insects, 12(4), 292. https://doi.org/10.3390/insects12040292