
Characterizing and Quantifying Arbovirus Transmission by Aedes aegypti Using Forced Salivation and Analysis of Bloodmeals

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Virus and Cells
2.2. Mosquito Infections
2.3. Mice Infection
2.4. Mosquito Sample Collections
2.5. Bloodmeal Dissections
2.6. RNA Extractions and qRT-PCR
2.7. Statistical Analyses
3. Results
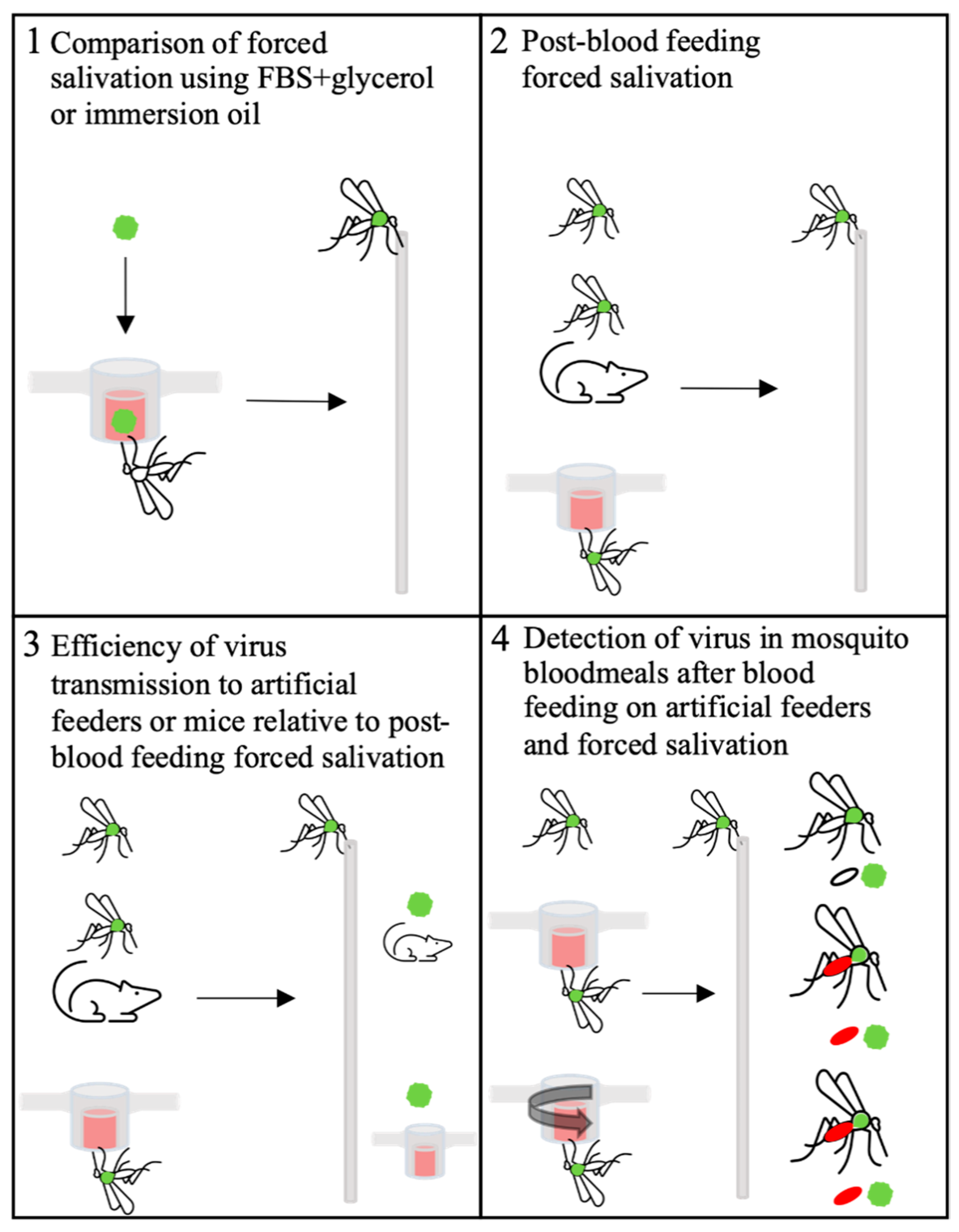
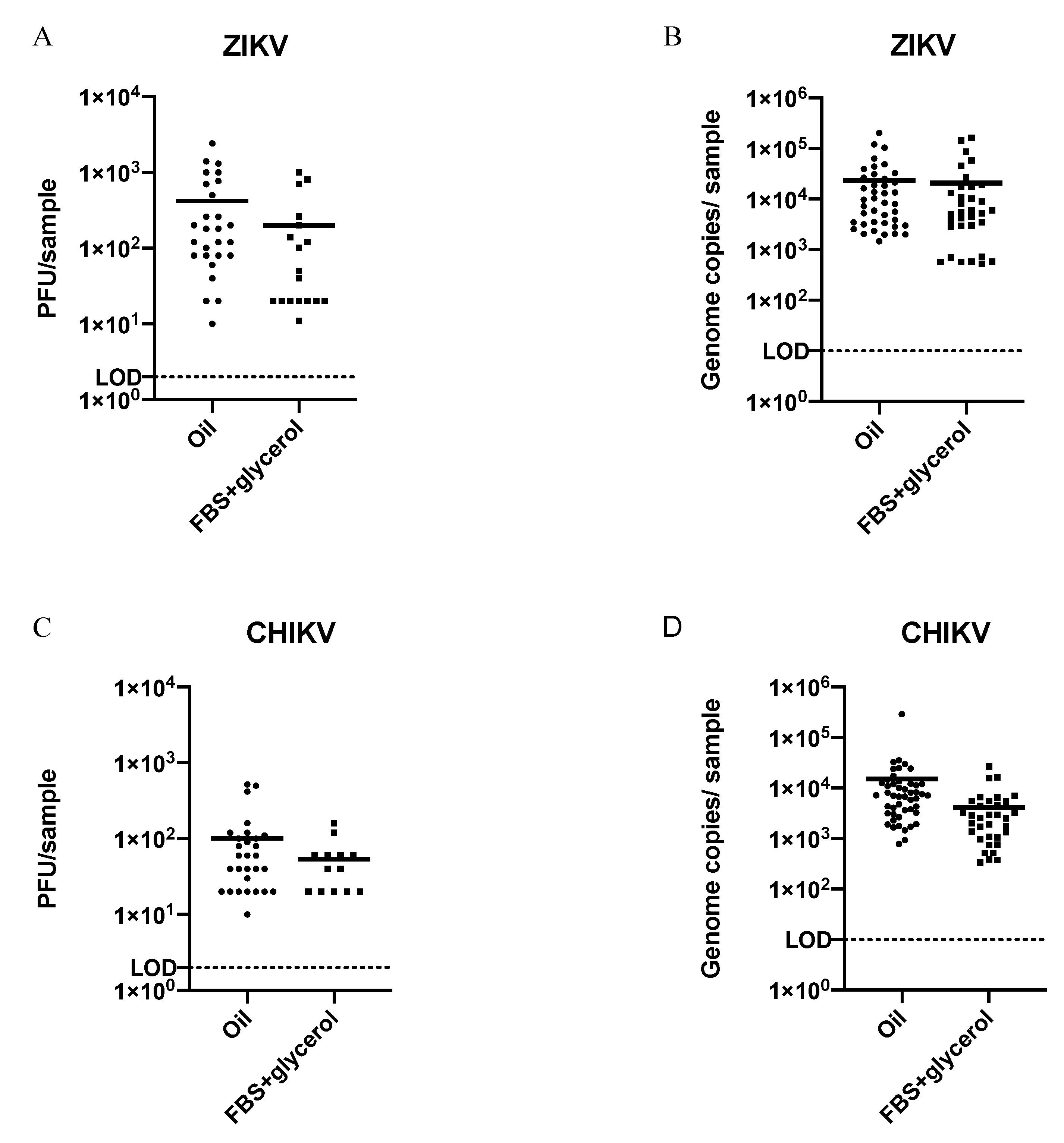
3.1. Comparison of Virus Detection from Saliva Collected in FBS + Glycerol or Immersion Oil
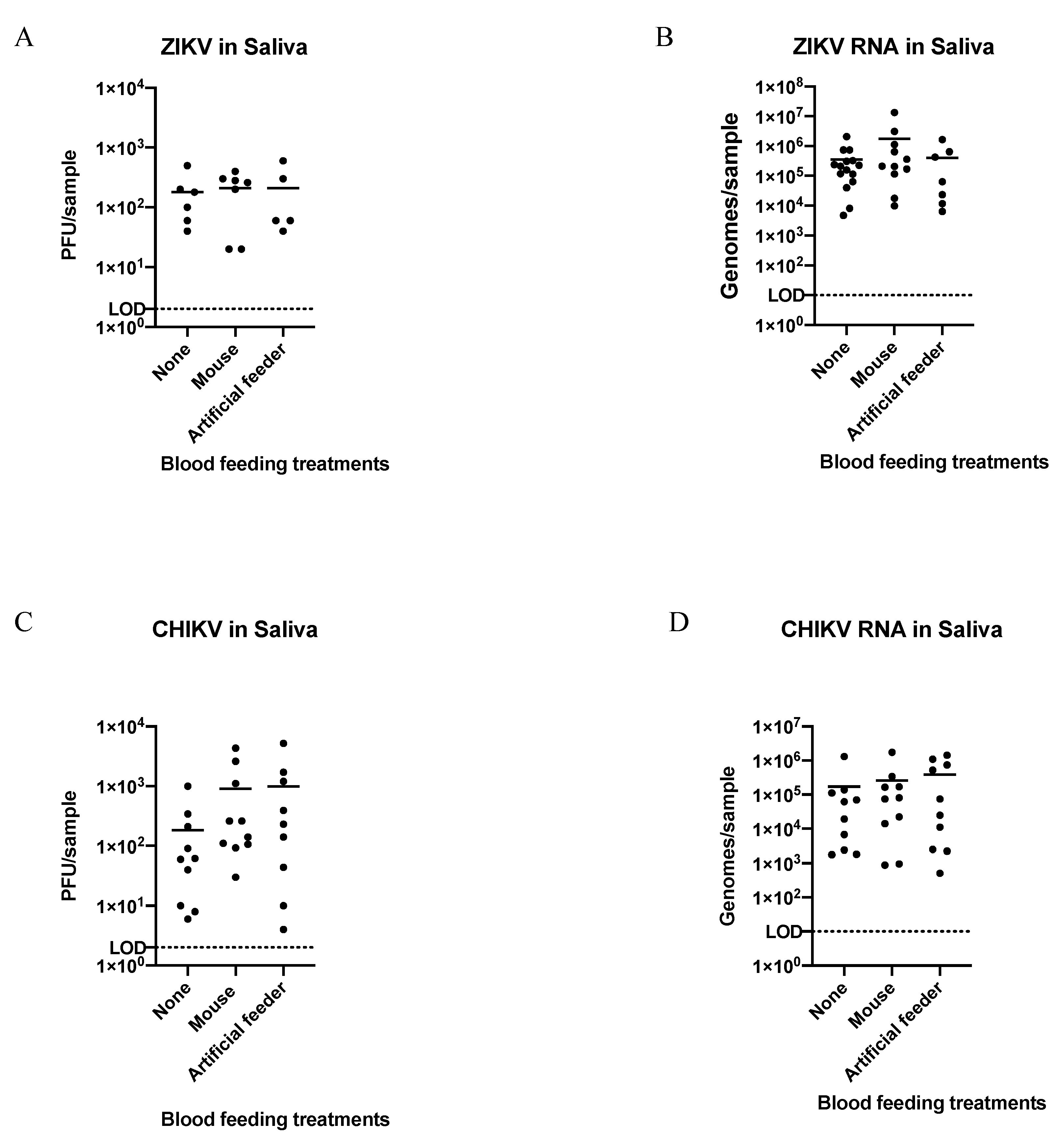
3.2. Detection of ZIKV or CHIKV from Mosquito Saliva Post-Blood Feeding
3.3. Efficiency of ZIKV or CHIKV Transmission to Artificial Feeders or Mice Relative to Post-Blood Feeding Forced Salivation
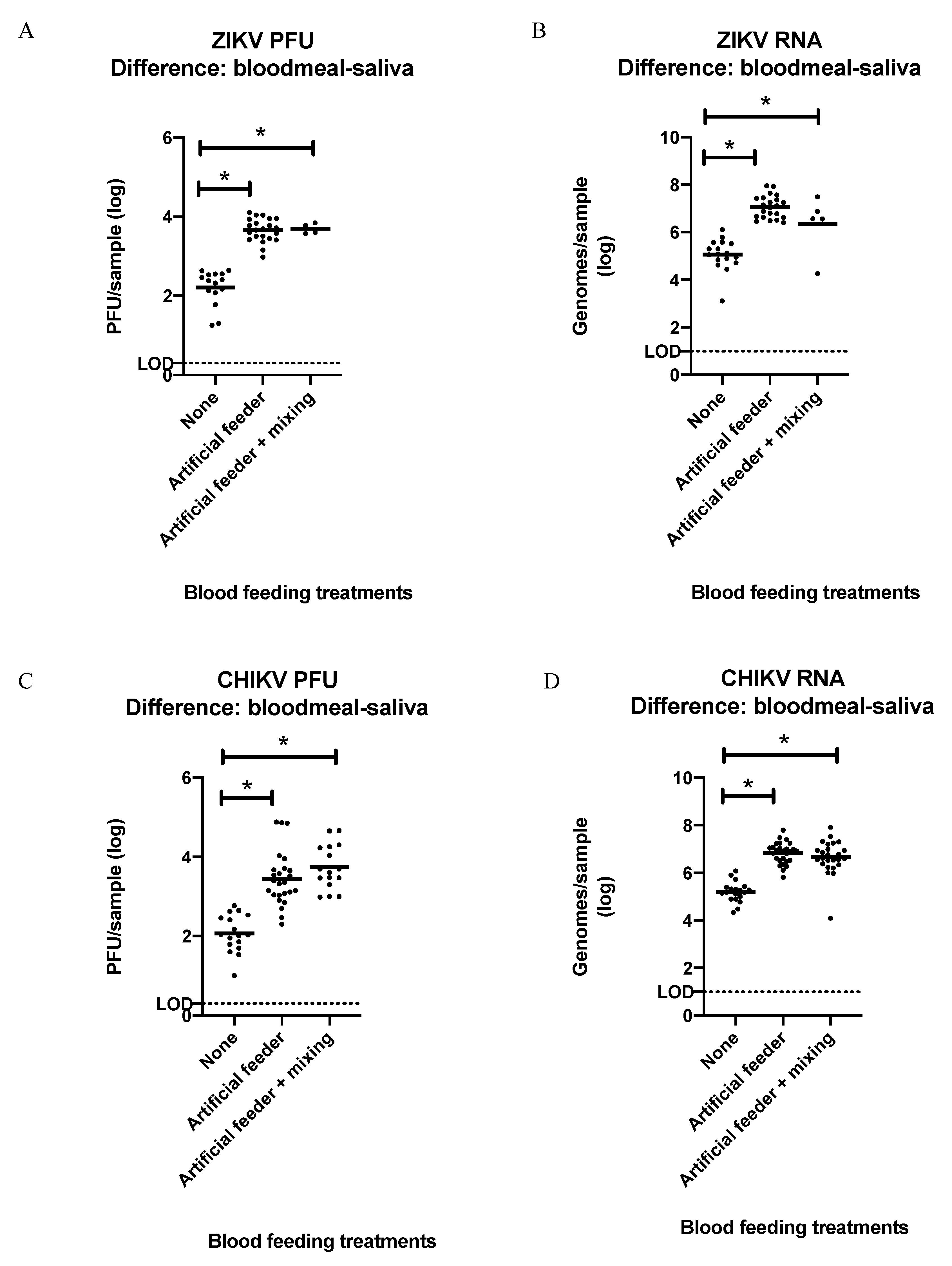
3.4. Detection of ZIKV or CHIKV in Mosquito Bloodmeals
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heitmann, A.; Jansen, S.; Lühken, R.; Leggewie, M.; Schmidt-Chanasit, J.; Tannich, E. Forced Salivation As a Method to Analyze Vector Competence of Mosquitoes. J. Vis. Exp. 2018, e57980. [Google Scholar] [CrossRef] [PubMed]
- Colton, L.; Biggerstaff, B.J.; Johnson, A.; Nasci, R.S. Quantification of West Nile Virus in Vector Mosquito Saliva. J. Am. Mosq. Control Assoc. 2005, 21, 49–53. [Google Scholar] [CrossRef]
- Anderson, S.L.; Richards, S.L.; Smartt, C.T. A simple method for determining arbovirus transmission in mosquitoes. J. Am. Mosq. Control Assoc. 2010, 26, 108–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, D.R.; Carrara, A.-S.; Aguilar, P.V.; Weaver, S.C. Evaluation of methods to assess transmission potential of venezuelan equine encephalitis virus by mosquitoes and estimation of mosquito saliva titers. Am. J. Trop. Med. Hyg. 2005, 73, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Gubler, D.J.; Rosen, L. A Simple Technique for Demonstrating Transmission of Dengue Virus by Mosquitoes without the Use of Vertebrate Hosts. Am. J. Trop. Med. Hyg. 1976, 25, 146–150. [Google Scholar] [CrossRef]
- Hurlbut, H.S. Mosquito Salivation and Virus Transmission. Am. J. Trop. Med. Hyg. 1966, 15, 989–993. [Google Scholar] [CrossRef]
- Danforth, M.E.; Reisen, W.K.; Barker, C.M. Detection of Arbovirus Transmission via Sugar Feeding in a Laboratory Setting. J. Med. Entomol. 2018, 55, 1575–1579. [Google Scholar] [CrossRef]
- Mores, C.N.; Turell, M.J.; Dohm, D.J.; Blow, J.A.; Carranza, M.T.; Quintana, M. Experimental Transmission of West Nile Virus by Culex nigripalpus from Honduras. Vector-Borne Zoonotic Dis. 2007, 7, 279–284. [Google Scholar] [CrossRef]
- Grosz, D.D.; van Geelen, A.; Gallup, J.M.; Hostetter, S.J.; Derscheid, R.J.; Ackermann, M.R. Sucrose stabilization of Respiratory Syncytial Virus (RSV) during nebulization and experimental infection. BMC Res. Notes 2014, 7, 158. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro, J.M.C. Blood-feeding in mosquitoes: Probing time and salivary gland anti-haemostatic activities in representatives of three genera (Aedes, Anopheles, Culex). Med. Vet.Entomol. 2000, 14, 142–148. [Google Scholar] [CrossRef]
- Ohambeblain, E.W.; Kis8ling, B.E.; Sike8, B.K. Studies on the North American Arthropod-Borne Encephalitides vh. Estimation of Amount of Eastern Equine Encephalitis Vxbus Inoculated by Infected Aede8 Aeoypti. Am. J. Epidemiol. 1954, 60, 286–291. [Google Scholar]
- Weaver, S.C.; Scott, T.W.; Lorenz, L.H. Patterns of Eastern Equine Encephalomyelitis Virus Infection in Culiseta melanura (Diptera: Culicidae). J. Med. Entomol. 1990, 27, 878–891. [Google Scholar] [CrossRef]
- Styer, L.M.; Kent, K.A.; Albright, R.G.; Bennett, C.J.; Kramer, L.D.; Bernard, K.A. Mosquitoes Inoculate High Doses of West Nile Virus as They Probe and Feed on Live Hosts. PLoS Pathog. 2007, 3, e132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VanLandingham, D.L.; Beasley, D.; Klingler, K.; Higgs, S.; Huang, J.; Fair, J.; Schneider, B.S.; Hamilton, P. Real-time reverse transcriptase–polymerase chain reaction quantification of west nile virus transmitted by culex pipiens quinquefasciatus. Am. J. Trop. Med. Hyg. 2004, 71, 120–123. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Velez, J.O.; Lambert, A.J.; Johnson, A.J.; Stanfield, S.M.; Duffy, M.R. Genetic and serologic properties of Zika virus associated with an epidemic, Yap State, Micronesia, 2007. Emerg. Infect. Dis. 2008, 14, 1232–1239. [Google Scholar] [CrossRef] [PubMed]
- Goddard, L.B.; Roth, A.E.; Reisen, W.K.; Scott, T.W. Vector Competence of California Mosquitoes for West Nile virus. Emerg. Infect. Dis. 2002, 8, 1385–1391. [Google Scholar] [CrossRef]
- Cornel, A.J.; Jupp, P.G. Comparison of three methods for determining transmission rates in vector competence studies with Culex univittatus and West Nile and Sindbis viruses. J. Am. Mosq. Control Assoc. 1989, 5, 70–72. [Google Scholar]
- Aitken, T.H.G. An In vitro feeding technique for artificially demonstrating virus transmission by mosquitoes. Mosq. News 1977, 37, 130–133. [Google Scholar]
- Beaty, B.J.; Aitken, T.H.G. In vitro transmission of yellow fever virus by geographic strains of Aedes aegypti. Mosq. News 1979, 39, 232–238. [Google Scholar]
- Sanchez-Vargas, I.; Harrington, L.C.; Black, W.C.; Olson, K.E. Analysis of Salivary Glands and Saliva from Aedes albopictus and Aedes aegypti Infected with Chikungunya Viruses. Insects 2019, 10, 39. [Google Scholar] [CrossRef] [Green Version]
- Kebaier, C.; Vanderberg, J.P. Re-ingestion of Plasmodium berghei sporozoites after delivery into the host by mosquitoes. Am. J. Trop. Med. Hyg. 2006, 75, 1200–1204. [Google Scholar] [CrossRef] [PubMed]
- Robison, A.; Young, M.C.; Byas, A.D.; Rückert, C.; Ebel, G.D. Comparison of Chikungunya Virus and Zika Virus Replication and Transmission Dynamics in Aedes aegypti Mosquitoes. Am. J. Trop. Med. Hyg. 2020, 103, 869–875. [Google Scholar] [CrossRef]
- Yamamoto, D.S.; Yokomine, T.; Sumitani, M.; Yagi, K.; Matsuoka, H.; Yoshida, S. Visualization and live imaging analysis of a mosquito saliva protein in host animal skin using a transgenic mosquito with a secreted luciferase reporter system. Insect Mol. Biol. 2013, 22, 685–693. [Google Scholar] [CrossRef]
- Choumet, V.; Attout, T.; Chartier, L.; Khun, H.; Sautereau, J.; Robbe-Vincent, A.; Brey, P.T.; Huerre, M.; Bain, O. Visualizing Non Infectious and Infectious Anopheles gambiae Blood Feedings in Naive and Saliva-Immunized Mice. PLoS ONE 2012, 7, e50464. [Google Scholar] [CrossRef]
- Secundino, N.F.C.; Chaves, B.A.; Orfano, A.S.; Silveira, K.R.D.; Rodrigues, N.B.; Campolina, T.B.; Nacif-Pimenta, R.; Villegas, L.E.M.; Silva, B.; Lacerda, M.V.G.; et al. Zika virus transmission to mouse ear by mosquito bite: A laboratory model that replicates the natural transmission process. Parasites Vectors 2017, 10, 346. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Collection Media | qRT-PCR | p-Value | Plaque Assay | p-Value |
|---|---|---|---|---|---|
| ZIKV | FBS + glycerol | 42% (42/100) | * 0.0170 | 18% (18/100) | * 0.0455 |
| Oil | 65% (62/96) | 31% (30/96) | |||
| CHIKV | FBS + glycerol | 47% (46/98) | * ≤0.0001 | 14% (14/98) | * 0.0002 |
| Oil | 82% (82/100) | 38% (38/100) |
| Blood Feeding Treatments | Group ID | Number of Mosquitoes That Blood Fed | Titer (PFU/Sample) | Genome Copies/Sample |
|---|---|---|---|---|
| None | 1 | 1 | 0 | 4.0 × 104 |
| None | 2 | 1 | 0 | 2.1 × 105 |
| None | 3 | 2 | 1.0 × 102 | 4.7 × 103 |
| None | 4 | 2 | 0 | 8.1 × 103 |
| None | 5 | 3 | 0 | 1.1 × 105 |
| None | 6 | 3 | 4.0 × 101 | 6.3 × 105 |
| None | 7 | 5 | 2.0 × 102 | 3.1 × 105 |
| None | 8 | 5 | 0 | 2.6 × 106 |
| None | 9 | 7 | 1.8 × 102 | 2.3 × 105 |
| None | 10 | 7 | 0 | 1.5 × 105 |
| None | 11 | 10 | 0 | 2.2 × 105 |
| None | 12 | 10 | 0 | 7.3 × 105 |
| None | 13 | 7 | 5.0 × 102 | 7.3 × 105 |
| None | 14 | 10 | 0 | 1.1 × 105 |
| None | 15 | 10 | 6.0 × 101 | 3.2 × 105 |
| Mouse | 1 | 1 | 2.8 × 102 | 1.3 × 107 |
| Mouse | 2 | 1 | 0 | 9.8 × 103 |
| Mouse | 3 | 2 | 4.0 × 102 | 2.0 × 105 |
| Mouse | 4 | 2 | 0 | 2.0 × 105 |
| Mouse | 5 | 3 | 2.6 × 102 | 6.4 × 105 |
| Mouse | 6 | 3 | 2.0 × 101 | 1.7 × 104 |
| Mouse | 7 | 5 | 3.0 × 102 | 3.6 × 105 |
| Mouse | 8 | 5 | 0 | 1.1 × 105 |
| Mouse | 9 | 7 | 0 | 1.7 × 105 |
| Mouse | 10 | 7 | 2.0 × 101 | 3.1 × 106 |
| Mouse | 11 | 7 | 2.0 × 102 | 1.1 × 106 |
| Artificial feeder | 1 | 1 | 3.0 × 102 | 6.4 × 104 |
| Artificial feeder | 2 | 1 | 6.0 × 102 | 1.6 × 106 |
| Artificial feeder | 3 | 3 | 6.0 × 101 | 6.4 × 105 |
| Artificial feeder | 4 | 2 | 0 | 6.3 × 103 |
| Artificial feeder | 5 | 1 | 0 | 1.1 × 104 |
| Artificial feeder | 7 | 1 | 4.0 × 101 | 2.2 × 104 |
| Artificial feeder | 10 | 3 | 6.0 × 101 | 4.2 × 105 |
| Blood Feeding Treatments | Group ID | Number Blood Feed Mosquitoes | Titer (PFU/mL) | Genome Copies |
|---|---|---|---|---|
| None | 1 | 1 | 0 | 0 |
| None | 2 | 1 | 1.0 × 101 | 2.4 × 103 |
| None | 3 | 2 | 6.2 × 101 | 6.7 × 103 |
| None | 4 | 2 | 8 | 1.7 × 103 |
| None | 5 | 3 | 6 | 1.8 × 103 |
| None | 6 | 3 | 3.4 × 102 | 6.2 × 104 |
| None | 7 | 5 | 9.0 × 101 | 1.9 × 104 |
| None | 8 | 5 | 1.0 × 103 | 1.3 × 106 |
| None | 9 | 7 | 3.2 × 101 | 1.3 × 105 |
| None | 10 | 7 | 8.8 × 101 | 6.9 × 104 |
| None | 11 | 4 | 2.01 × 102 | 1.1 × 105 |
| Mouse | 1 | 1 | 0 | 8.7 ×102 |
| Mouse | 2 | 1 | 3.0 × 101 | 9.5 ×102 |
| Mouse | 3 | 2 | 1.0 × 102 | 1.4 × 104 |
| Mouse | 4 | 2 | 1.1 × 103 | 3.4 × 105 |
| Mouse | 5 | 3 | 1.4 × 102 | 0 |
| Mouse | 6 | 3 | 4.3 × 103 | 7.5 × 105 |
| Mouse | 7 | 5 | 2.6 × 102 | 1.6 × 105 |
| Mouse | 8 | 5 | 2.6 × 102 | 1.3 × 105 |
| Mouse | 9 | 7 | 1.1 × 102 | 8.0 × 104 |
| Mouse | 10 | 7 | 2.6 × 103 | 1.7 × 106 |
| Mouse | 11 | 6 | 9.2 × 101 | 2.2 × 104 |
| Artificial feeder | 1 | 1 | 0 | 0 |
| Artificial feeder | 2 | 1 | 4 | 2.2 × 104 |
| Artificial feeder | 3 | 2 | 0 | 5.3 × 102 |
| Artificial feeder | 4 | 2 | 1.4 × 102 | 1.1 × 104 |
| Artificial feeder | 5 | 3 | 1.0 × 101 | 2.4 × 103 |
| Artificial feeder | 6 | 3 | 2.3 × 102 | 7.4 × 104 |
| Artificial feeder | 7 | 5 | 1.2 × 103 | 7.4 × 105 |
| Artificial feeder | 8 | 5 | 5.2 × 103 | 1.4 × 106 |
| Artificial feeder | 9 | 7 | 1.7 × 103 | 1.1 × 106 |
| Artificial feeder | 10 | 7 | 3.9 × 102 | 5.2 × 105 |
| Artificial feeder | 11 | 5 | 4.4 × 101 | 2.4 × 104 |
| Mouse ID | Number of Mosquitoes That Blood Fed | Pooled Saliva-Titer (PFU) | Pooled Saliva-Genome Copies | Mouse Became Infected as Determine by Viral RNA Detection in Tissue/Blood? |
|---|---|---|---|---|
| 080 | 4 | 0 | 2.8 × 105 | Yes |
| 086 | 2 | 0 | 2.0 × 104 | Yes |
| 095 | 1 | 0 | 7.2 × 102 | Yes |
| 109 | 2 | 0 | 2.0 × 104 | Yes |
| 165 | 3 | 0 | 1.3 × 103 | Yes |
| 166 | 1 | 0 | 5.0 × 103 | Yes |
| 169 | 2 | 0 | 6.8 × 104 | Yes |
| 174 | 3 | 0 | 3.3 × 103 | Yes |
| Group Number | Number of Mosquitoes that Blood Fed | Pooled Saliva-Titer (PFU) | Pooled Saliva-Genome Copies | Remaining Blood in the Artificial Feeder-Titer (PFU) | Remaining Blood in the Artificial Feeder-Genome Copies |
|---|---|---|---|---|---|
| 1 | 2 | 8.0 × 101 | 1.1 × 105 | 0 | 2.1 × 103 |
| 2 | 6 | 3.0 × 101 | 1.6 × 104 | 0 | 7.0 × 102 |
| 3 | 4 | 1.8 × 101 | 9.5 × 103 | 0 | 0 |
| 4 | 5 | 1.2 × 102 | 2.2 × 105 | 0 | 3.5 × 103 |
| 5 | 9 | 4.0 × 101 | 1.9 × 105 | 0 | 0 |
| 6 | 2 | 5.0 × 101 | 4.5 × 105 | 0 | 0 |
| 7 | 7 | 2.3 × 102 | 1.9 × 105 | 0 | 0 |
| 8 | 5 | 1.0 × 102 | 1.2 × 106 | 0 | 0 |
| 9 | 3 | 2.3 × 102 | 9.0 × 104 | 0 | 1.7 × 103 |
| Group Number | Number of Mosquitoes That Blood Fed | Pooled Saliva-Titer (PFU) | Pooled Saliva-Genome Copies | Remaining Blood in the Artificial Feeder-Titer (PFU) | Remaining Blood in the Artificial Feeder-Genome Copies |
|---|---|---|---|---|---|
| 1 | 2 | 2.1 × 102 | 1.1 × 105 | 0 | 0 |
| 2 | 11 | 3.7 × 103 | 1.8 × 106 | 0 | 0 |
| 3 | 10 | 2.0 × 102 | 9.5 × 104 | 0 | 0 |
| 4 | 7 | 3.3 × 102 | 5.0 × 104 | 0 | 2.8 × 103 |
| 5 | 6 | 2.1 × 102 | 1.0 × 105 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miller, M.R.; Sorensen, M.R.; Markle, E.D.; Clarkson, T.C.; Knight, A.L.; Savran, M.J.; Foy, B.D. Characterizing and Quantifying Arbovirus Transmission by Aedes aegypti Using Forced Salivation and Analysis of Bloodmeals. Insects 2021, 12, 304. https://doi.org/10.3390/insects12040304
Miller MR, Sorensen MR, Markle ED, Clarkson TC, Knight AL, Savran MJ, Foy BD. Characterizing and Quantifying Arbovirus Transmission by Aedes aegypti Using Forced Salivation and Analysis of Bloodmeals. Insects. 2021; 12(4):304. https://doi.org/10.3390/insects12040304
Chicago/Turabian StyleMiller, Megan R., Madeleine R. Sorensen, Erin D. Markle, Taylor C. Clarkson, Ashley L. Knight, Michelle J. Savran, and Brian D. Foy. 2021. "Characterizing and Quantifying Arbovirus Transmission by Aedes aegypti Using Forced Salivation and Analysis of Bloodmeals" Insects 12, no. 4: 304. https://doi.org/10.3390/insects12040304
APA StyleMiller, M. R., Sorensen, M. R., Markle, E. D., Clarkson, T. C., Knight, A. L., Savran, M. J., & Foy, B. D. (2021). Characterizing and Quantifying Arbovirus Transmission by Aedes aegypti Using Forced Salivation and Analysis of Bloodmeals. Insects, 12(4), 304. https://doi.org/10.3390/insects12040304