
Predation on Drosophila suzukii within Hedges in the Agricultural Landscape

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Sites
2.2. Exposure of D. suzukii Pupae in the Field
2.3. Collection of Arthropod Predators
2.4. Molecular Gut Content Analysis
2.4.1. DNA-Extraction
2.4.2. PCR
2.4.3. Gel Electrophoresis
2.5. Statistical Analyses
3. Results
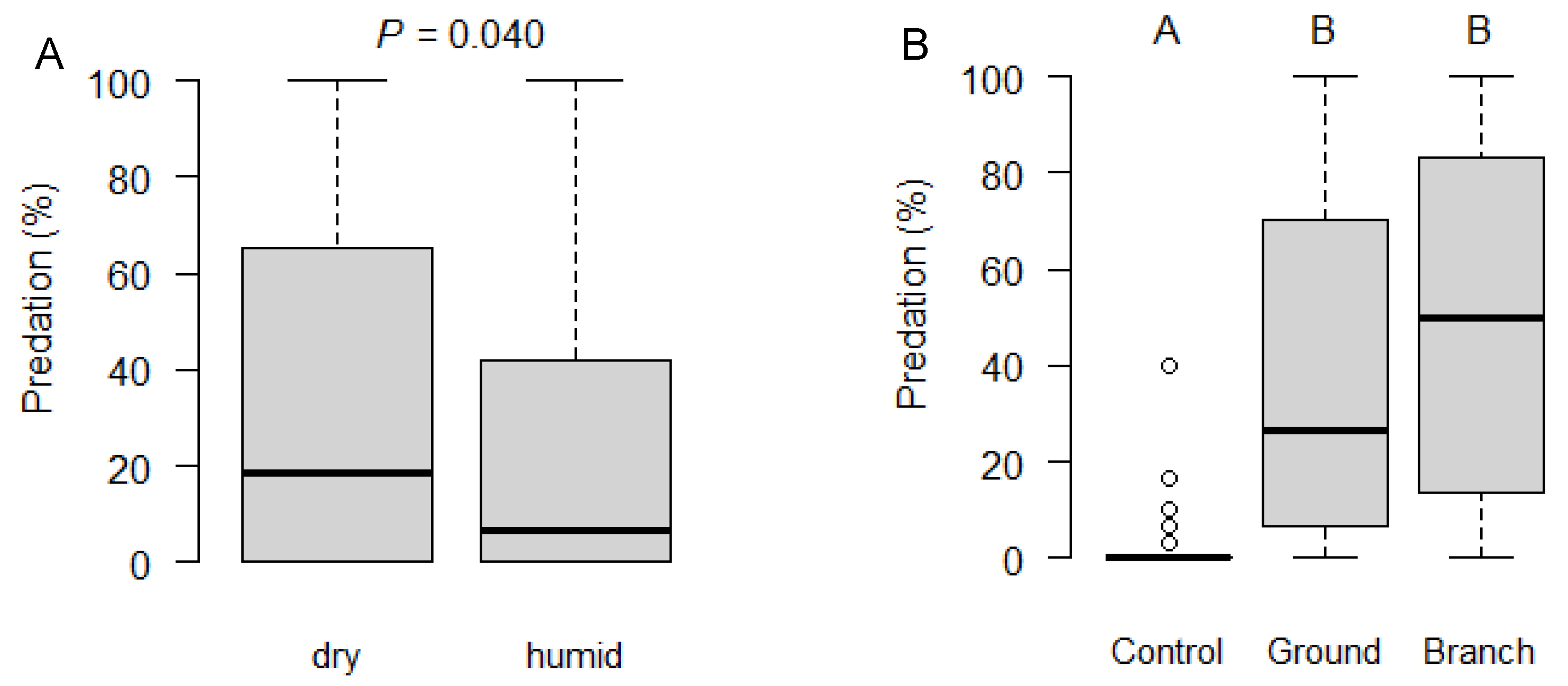
3.1. Exposure of D. suzukii Pupae in the Field
3.2. Collection of Arthropod Predators
3.3. Molecular Gut Content Analyses
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walsh, D.B.; Bolda, M.P.; Goodhue, R.E.; Dreves, A.J.; Lee, J.; Bruck, D.J.; Walton, V.M.; O’Neal, S.D.; Zalom, F.G. Drosophila suzukii (Diptera: Drosophilidae): Invasive Pest of Ripening Soft Fruit Expanding its Geographic Range and Damage Potential. J. Integr. Pest Manag. 2011, 2, G1–G7. [Google Scholar] [CrossRef]
- Mazzi, D.; Bravin, E.; Meraner, M.; Finger, R.; Kuske, S. Economic Impact of the Introduction and Establishment of Drosophila suzukii on Sweet Cherry Production in Switzerland. Insects 2017, 8, 18. [Google Scholar] [CrossRef] [Green Version]
- Poyet, M.; Le Roux, V.; Gibert, P.; Meirland, A.; Prévost, G.; Eslin, P.; Chabrerie, O. The Wide Potential Trophic Niche of the Asiatic Fruit Fly Drosophila suzukii: The Key of its Invasion Success in Temperate Europe? PLoS ONE 2015, 10, e0142785. [Google Scholar] [CrossRef] [Green Version]
- Kenis, M.; Tonina, L.; Eschen, R.; Van Der Sluis, B.; Sancassani, M.; Mori, N.; Haye, T.; Helsen, H. Non-crop plants used as hosts by Drosophila suzukii in Europe. J. Pest Sci. 2016, 89, 735–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, S.; Boycheva-Woltering, S.; Romeis, J.; Collatz, J. Trichopria drosophilae parasitizes Drosophila suzukii in seven common non-crop fruits. J. Pest Sci. 2019, 93, 627–638. [Google Scholar] [CrossRef]
- Santoiemma, G.; Trivellato, F.; Caloi, V.; Mori, N.; Marini, L. Habitat preference of Drosophila suzukii across heterogeneous landscapes. J. Pest Sci. 2018, 92, 485–494. [Google Scholar] [CrossRef]
- Rossi-Stacconi, M.V.; Kaur, R.; Mazzoni, V.; Ometto, L.; Grassi, A.; Gottardello, A.; Rota-Stabelli, O.; Anfora, G. Multiple lines of evidence for reproductive winter diapause in the invasive pest Drosophila suzukii: Useful clues for control strategies. J. Pest Sci. 2016, 89, 689–700. [Google Scholar] [CrossRef]
- Holland, J.M.; Bianchi, F.J.; Entling, M.H.; Moonen, A.-C.; Smith, B.M.; Jeanneret, P. Structure, function and management of semi-natural habitats for conservation biological control: A review of European studies. Pest Manag. Sci. 2016, 72, 1638–1651. [Google Scholar] [CrossRef]
- Tscharntke, T.; Karp, D.S.; Chaplin-Kramer, R.; Batáry, P.; DeClerck, F.; Gratton, C.; Hunt, L.; Ives, A.; Jonsson, M.; Larsen, A.; et al. When natural habitat fails to enhance biological pest control—Five hypotheses. Biol. Conserv. 2016, 204, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Cahenzli, F.; Bühlmann, I.; Daniel, C.; Fahrentrapp, J. The distance between forests and crops affects the abundance of Dro-sophila suzukii during fruit ripening, but not during harvest. Environ. Entomol. 2018, 47, 1274–1279. [Google Scholar] [CrossRef]
- Hennig, E.I.; Mazzi, D. Spotted Wing Drosophila in Sweet Cherry Orchards in Relation to Forest Characteristics, Bycatch and Resource Availability. Insects 2018, 9, 118. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Saona, C.; Firbas, N.; Hernández-Cumplido, J.; Holdcraft, R.; Michel, C.; Palacios-Castro, S.; Silva, D.B. Interpreting Temporal and Spatial Variation in Spotted-Wing Drosophila (Diptera: Drosophilidae) Trap Captures in Highbush Blueberries. J. Econ. Entomol. 2020, 113, 2362–2371. [Google Scholar] [CrossRef]
- Schmidt, J.M.; Whitehouse, T.S.; Green, K.; Krehenwinkel, H.; Schmidt-Jeffris, R.; Sial, A.A. Local and landscape-scale heterogeneity shape spotted wing drosophila (Drosophila suzukii) activity and natural enemy abundance: Implications for trophic interactions. Agr. Ecosyst. Environ. 2019, 272, 86–94. [Google Scholar] [CrossRef]
- Chaplin-Kramer, R.; O’Rourke, M.E.; Blitzer, E.J.; Kremen, C. A meta-analysis of crop pest and natural enemy response to landscape complexity. Ecol. Lett. 2011, 14, 922–932. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, F.; Booij, C.; Tscharntke, T. Sustainable pest regulation in agricultural landscapes: A review on landscape composition, biodiversity and natural pest control. Proc. R. Soc. B Biol. Sci. 2006, 273, 1715–1727. [Google Scholar] [CrossRef] [Green Version]
- Fischer, C.; Schlinkert, H.; Ludwig, M.; Holzschuh, A.; Gallé, R.; Tscharntke, T.; Batáry, P. The impact of hedge-forest con-nectivity and microhabitat conditions on spider and carabid beetle assemblages in agricultural landscapes. J. Ins. Cons. 2013, 17, 1027–1038. [Google Scholar] [CrossRef]
- Holland, J.M. Contribution of hedgerows to biological control. In The Ecology of Hedgerows and Field Margins, 1st ed.; Dover, J.W., Ed.; Routledge: Oxford, UK, 2019; pp. 123–146. [Google Scholar]
- Gillespie, M.A.; Gurr, G.M.; Wratten, S.D. Beyond nectar provision: The other resource requirements of parasitoid biological control agents. Entomol. Exp. Appl. 2016, 159, 207–221. [Google Scholar] [CrossRef]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat Management to Conserve Natural Enemies of Arthropod Pests in Agriculture. Annu. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef] [PubMed]
- Gurr, G.M.; Wratten, S.D.; Landis, D.A.; You, M. Habitat Management to Suppress Pest Populations: Progress and Prospects. Annu. Rev. Entomol. 2017, 62, 91–109. [Google Scholar] [CrossRef] [PubMed]
- Rusch, A.; Bommarco, R.; Ekbom, B. Conservation Biological Control in Agricultural Landscapes. Adv. Bot. Res. 2017, 81, 333–360. [Google Scholar] [CrossRef]
- Wolf, S.; Zeisler, C.; Sint, D.; Romeis, J.; Traugott, M.; Collatz, J. A simple and cost-effective molecular method to track predation on Drosophila suzukii in the field. J. Pest. Sci. 2018, 91, 927–935. [Google Scholar] [CrossRef]
- Symondson, W.O.C.; Sunderland, K.D.; Greenstone, M.H. Can generalist predators be effective biocontrol agents? Annu. Rev. Entomol. 2002, 47, 561–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bourne, A.; Fountain, M.T.; Wijnen, H.; Shaw, B. Potential of the European earwig (Forficula auricularia) as a biocontrol agent of the soft and stone fruit pest Drosophila suzukii. Pest Manag. Sci. 2019, 75, 3340–3345. [Google Scholar] [CrossRef] [PubMed]
- Englert, C.; Herz, A. Acceptability of Drosophila suzukii as prey for common predators occurring in cherries and berries. J. Appl. Entomol. 2018, 143, 387–396. [Google Scholar] [CrossRef]
- Woltering, S.B.; Romeis, J.; Collatz, J. Influence of the Rearing Host on Biological Parameters of Trichopria drosophilae, a Potential Biological Control Agent of Drosophila suzukii. Insects 2019, 10, 183. [Google Scholar] [CrossRef] [Green Version]
- Greenstone, M.H.; Weber, D.C.; Coudron, T.A.; Payton, M.E.; Hu, J.S. Removing external DNA contamination from arthropod predators destined for molecular gut-content analysis. Mol. Ecol. Resour. 2012, 12, 464–469. [Google Scholar] [CrossRef] [PubMed]
- Wallinger, C.; Staudacher, K.; Schallhart, N.; Peter, E.; Dresch, P.; Juen, A.; Traugott, M. The effect of plant identity and the level of plant decay on molecular gut content analysis in a herbivorous soil insect. Mol. Ecol. Resour. 2012, 13, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- R Version 4.0.1 (2020-06-06)—“See Things Now”, Copyright (C) 2020 The R Foundation for Statistical Computing, Platform: i386-w64-mingw32/i386 (32-Bit). Index of /bin/windows/base/old/4.0.1 (r-project.org). Available online: https://cran.r-project.org/bin/windows/base/old/4.0.1/ (accessed on 29 March 2021).
- Lenth, R.V. Least-Squares Means: The R Package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Wagner, H. Vegan: Community Ecology Package. 2019. Available online: https://CRAN.R-project.org/package=vegan (accessed on 29 March 2021).
- Lee, J.C.; Wang, X.; Daane, K.M.; Hoelmer, K.A.; Isaacs, R.; Sial, A.A.; Walton, V.M. Biological control of spotted-wing dro-sophila (Diptera: Drosophilidae)-current and pending tactics. J. Integr. Pest Manag. 2019, 10, 13. [Google Scholar] [CrossRef]
- Van Lenteren, J.C.; Sexsmith, W. Guidance for information requirements for regulation of invertebrates as biological control agents (IBCAs). OECD Ser. Pestic. 2004, 21, 1–22. [Google Scholar]
- Woltz, J.M.; Lee, J.C. Pupation behavior and larval and pupal biocontrol of Drosophila suzukii in the field. Biol. Control. 2017, 110, 62–69. [Google Scholar] [CrossRef] [Green Version]
- Ballman, E.S.A.; Collins, J.A.; Drummond, F. Pupation Behavior and Predation on Drosophila suzukii (Diptera: Drosophilidae) Pupae in Maine Wild Blueberry Fields. J. Econ. Entomol. 2017, 110, 2308–2317. [Google Scholar] [CrossRef] [PubMed]
- Rossi Stacconi, V.M.; Ouantar, M.; Grassi, A.; Baser, N.; Loni, A.; Walton, V.V.; Ioriatti, C.; Anfora, G. Field collection and laboratory efficacy of parasitoids adapted to Drosophila suzukii in newly invaded areas of Trentino, Italy. IOBC WPRS Bull. 2015, 109, 177–179. [Google Scholar]
- Miller, B.; Anfora, G.; Buffington, M.; Daane, K.M.; Dalton, D.T.; Hoelmer, K.M.; Rossi Stacconi, M.V.; Grassi, A.; Ioriatti, C.; Loni, A.; et al. Seasonal occurrence of resident parasitoids associated with Drosophila suzukii in two small fruit production re-gions of Italy and the USA. Bull. Insectol. 2015, 68, 255–263. [Google Scholar]
- Chabert, S.; Allemand, R.; Poyet, M.; Eslin, P.; Gibert, P. Ability of European parasitoids (Hymenoptera) to control a new invasive Asiatic pest, Drosophila suzukii. Biol. Control. 2012, 63, 40–47. [Google Scholar] [CrossRef]
- Knoll, V.; Ellenbroek, T.; Romeis, J.; Collatz, J. Seasonal and regional presence of hymenopteran parasitoids of Drosophila in Switzerland and their ability to parasitize the invasive Drosophila suzukii. Sci. Rep. 2017, 7, 40697. [Google Scholar] [CrossRef] [PubMed]
- Gabarra, R.; Riudavets, J.; Rodríguez, G.A.; Pujade-Villar, J.; Arnó, J. Prospects for the biological control of Drosophila suzukii. BioControl 2015, 60, 331–339. [Google Scholar] [CrossRef]
- King, R.A.; Read, D.S.; Traugott, M.; Symondson, W.O.C. Molecular analysis of predation: A review of best practice for DNA-based approaches. Mol. Ecol. 2008, 17, 947–963. [Google Scholar] [CrossRef]
- Kraft, K. Molekular Biologischer Nachweis von Drosophila suzukii Matsumura (Diptera: Drosophilidae) in räuberisch lebenden Arthropoden. Ph. D. Thesis, Julius Kühn Institut, Darmstadt, Germany, 2017. [Google Scholar]
- Schetelig, M.F.; Lee, K.-Z.; Otto, S.; Talmann, L.; Stökl, J.; Degenkolb, T.; Vilcinskas, A.; Halitschke, R. Environmentally sustainable pest control options for Drosophila suzukii. J. Appl. Entomol. 2018, 142, 3–17. [Google Scholar] [CrossRef]
- Huth, C.; Schirra, K.; Seitz, A.; Louis, F. A beneficial species becomes a pest—The common earwig Forficula auricularia (Linnaeus 1758). IOBC WPRS Bull. 2011, 67, 249–256. [Google Scholar]
- Orpet, R.J.; Crowder, D.W.; Jones, V.P. Biology and Management of European Earwig in Orchards and Vineyards. J. Integr. Pest Manag. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Quarrell, S.R.; Corkrey, R.; Allen, G.R. Cherry damage and the spatial distribution of European earwigs, (Forficula auricularia L.) in sweet cherry trees. Pest Manag. Sci. 2021, 77, 159–167. [Google Scholar] [CrossRef]
- Nyffeler, M.; Benz, G. Spiders in natural pest control: A review. J. Appl. Entomol. 1987, 103, 321–339. [Google Scholar] [CrossRef] [Green Version]
- Tonina, L.; Mori, N.; Sancassani, M.; Dall’Ara, P.; Marini, L. Spillover of Drosophila suzukii between noncrop and crop areas: Implications for pest management. Agric. For. Entomol. 2018, 20, 575–581. [Google Scholar] [CrossRef]
- Weißinger, L.; Schrieber, K.; Breuer, M.; Müller, C. Influences of blackberry margins on population dynamics of Drosophila suzukii and grape infestation in adjacent vineyards. J. Appl. Entomol. 2019, 143, 802–812. [Google Scholar] [CrossRef]
- Shelton, A.; Badenes-Perez, F. Concepts and applications of trap cropping in pest management. Annu. Rev. Entomol. 2006, 51, 285–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alhmedi, A.; Clymans, R.; Van Kerckvoorde, V.; Bylemans, D.; Beliën, T. Preference and performance of Drosophila suzukii on Prunus species: A potential eco-friendly pest management tool. Crop. Prot. 2019, 122, 35–41. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Location (Canton) | Exposure Period | Sampling Predators | Hedge Type | Lat. [N]/ Lon. [E] | Host Plant Species |
|---|---|---|---|---|---|
| Changins (VD) | 1.7–4.7 (1) 22.7–25.7 (2) 27.8–30.8 (3) 3.10–6.10 (4) | 1.7–4.7 (1) 22.7 (2) 26.8–27.8 (3) 30.9 (4) 3.10–4.10 (4) | dry | 46°24′09.1″/ 6°13′42.4″ | Cornus sanguinea, Euonymus europaeus, Hedera helix, Lonicera xylosteum, Prunus avium, P. spinosa, Rosa spp., Rubus spp., Sambucus nigra, Viburnum lantana, V. opulus |
| humid | 46°23′47.3″/ 6°13′42.0″ | Bryonia dioica, Cornus sanguinea, Rubus spp., Sambucus nigra | |||
| Conthey (VS) | 16.7–18.7 (2) 12.8–15.8 (3) 4.10–7.10 (4) | 16.7 (2) 12.8–13.8 (3) 27.8 (3) 2.10–4.10 (4) | dry | 46°12′37.6″/ 7°18′02.6″ | Cornus spp., Sambucus nigra, Viburnum lantana, V. opulus |
| humid | 46°14′07.5″/ 7°18′35.7″ | Hipophae rhamnoides, Prunus avium, P. cerasifera, P. mahaleb, Rosa spp., Sambucus nigra | |||
| Frick (AG) | 24.6–28.6 (1) * 21.7–26.7 (2) 13.8–16.8 (3) 17.9–20.9 (4) 15.10–18.10 (4) | 25.6 (1) 23.7–25.7 (2) 12.8–14.8 (3) 17.9, 18.9 (4) 16.10 (4) | dry | 47°30′58.3″/ 8°1′26.6″ | Amelanchier ovalis, Cornus spp., Crataegus spp., Euonymus europaeus, Ligustrum spp., Lonicera xylosteum, P. spinosa, Rhamnus frangula, Rosa spp., Rubus spp., Sambucus nigra, Viburnum opulus |
| humid | 47°30′58.28″/ 8°1′26.62″ | Cornus spp., Crataegus spp., Euonymus europaeus, Ligustrum spp., Prunus spinosa, Rhamnus frangula, Rosa spp., Rubus spp., Sambucus nigra, Viburnum opulus | |||
| Reckenholz (ZH) | 25.6–28.6 (1) 8.7–12.7 (2) 22.8–26.8 (3) 4.10–8.10 (4) | 17.6. (1) 8.7–9.7 (2) 22.8 (3) 4.10 (4) | dry | 47°25′34.9″/ 8°31′02.4″ | Cornus sanguinea, Crataegus spp., Ligustrum vulgare, Lonicera xylosteum, Rosa canina, Rhamnus cathartica |
| humid | 47°25′32.9″/ 8°31′18.8″ | Crataegus spp., Prunus spinosa | |||
| Wädenswil (ZH) | 5.7–8.7 (2) 30.8–2.9 (3) * 1.10–4.10 (4) | 9.7 (2) 4.9 (3) 3.10–4.10 (4) | dry | 47°21′8.11″/ 8°68′24.2″ | Amelanchier ovalis, Cornus spp., Crataegus spp., Euonymus europaeus, Hedera helix, Ligustrum vulgare, Prunus padus, P. spinosa, Rhamnus cathartica, Rosa spp., Rubus spp., Sambucus nigra, S. racemosa, Taxus baccata, Viburnum opulus |
| humid | 47°22′37.5″/ 8°67′59.3″ | Cornus spp., Hedera helix, Ligustrum vulgare, Rubus spp., Sambucus nigra |
| Changins | Conthey | Frick | Reckenholz | Wädenswil | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| D | H | D | H | D | H | D | H | D | H | |
| Number exposed | 210 | 243 | 180 | 190 | 270 | 270 | 240 | 240 | 150 | 120 |
| Number predated | 114 | 27 | 77 | 87 | 109 | 124 | 152 | 144 | 78 | 17 |
| Mean predation [%] | 54.3 | 10.7 | 42.8 | 47.5 | 40.3 | 46.0 | 63.3 | 60.0 | 52.2 | 12.7 |
| SE predation | 16.6 | 6.4 | 15.4 | 12.8 | 10.7 | 10.5 | 6.3 | 9.9 | 21.4 | 8.1 |
| Mean unaccounted [%] | 18.1 | 25.6 | 12.2 | 20.8 | 45.0 | 46.3 | 15.4 | 8.8 | 6.7 | 7.3 |
| SE unaccounted | 10.7 | 6.4 | 5.0 | 6.0 | 11.5 | 11.4 | 3.0 | 1.8 | 4.8 | 3.1 |
| Samples | 7 | 8 | 6 | 6 | 10 | 10 | 8 | 8 | 6 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siffert, A.; Cahenzli, F.; Kehrli, P.; Daniel, C.; Dekumbis, V.; Egger, B.; Furtwengler, J.; Minguely, C.; Stäheli, N.; Widmer, F.; et al. Predation on Drosophila suzukii within Hedges in the Agricultural Landscape. Insects 2021, 12, 305. https://doi.org/10.3390/insects12040305
Siffert A, Cahenzli F, Kehrli P, Daniel C, Dekumbis V, Egger B, Furtwengler J, Minguely C, Stäheli N, Widmer F, et al. Predation on Drosophila suzukii within Hedges in the Agricultural Landscape. Insects. 2021; 12(4):305. https://doi.org/10.3390/insects12040305
Chicago/Turabian StyleSiffert, Alexandra, Fabian Cahenzli, Patrik Kehrli, Claudia Daniel, Virginie Dekumbis, Barbara Egger, Jana Furtwengler, Camille Minguely, Nicola Stäheli, Franco Widmer, and et al. 2021. "Predation on Drosophila suzukii within Hedges in the Agricultural Landscape" Insects 12, no. 4: 305. https://doi.org/10.3390/insects12040305
APA StyleSiffert, A., Cahenzli, F., Kehrli, P., Daniel, C., Dekumbis, V., Egger, B., Furtwengler, J., Minguely, C., Stäheli, N., Widmer, F., Mazzi, D., & Collatz, J. (2021). Predation on Drosophila suzukii within Hedges in the Agricultural Landscape. Insects, 12(4), 305. https://doi.org/10.3390/insects12040305