
Assessing the Distribution of Exotic Egg Parasitoids of Halyomorpha halys in Europe with a Large-Scale Monitoring Program

 ,
,  ,
,  ,
,  ,
,  , , , , , , , ,
, , , , , , , ,  add
Show full author list
add
Show full author list
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Surveys
2.2. Laboratory Handling of Collected Egg Masses
2.3. Taxonomic Identification
2.4. Data Analysis
3. Results
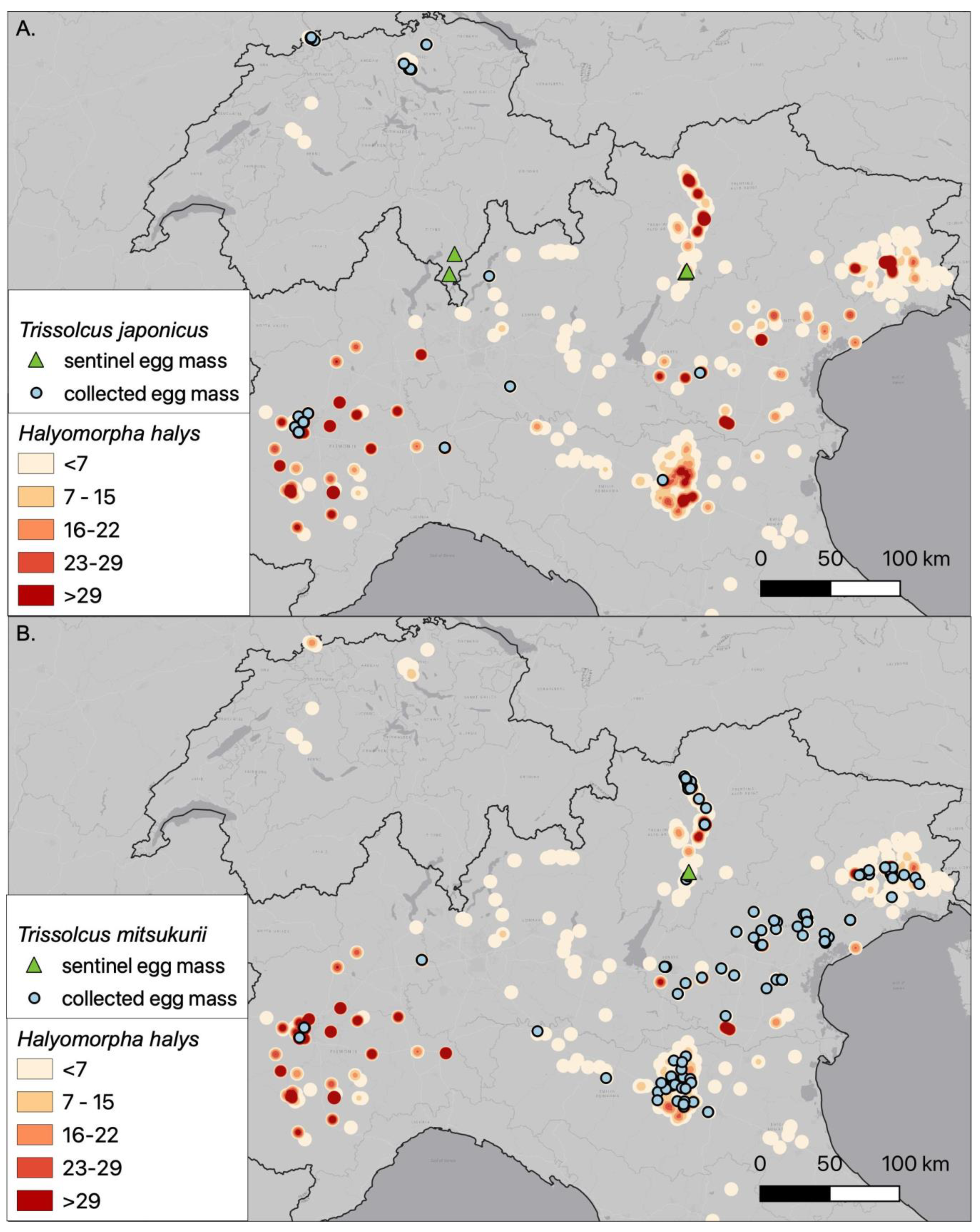
3.1. Field Survey
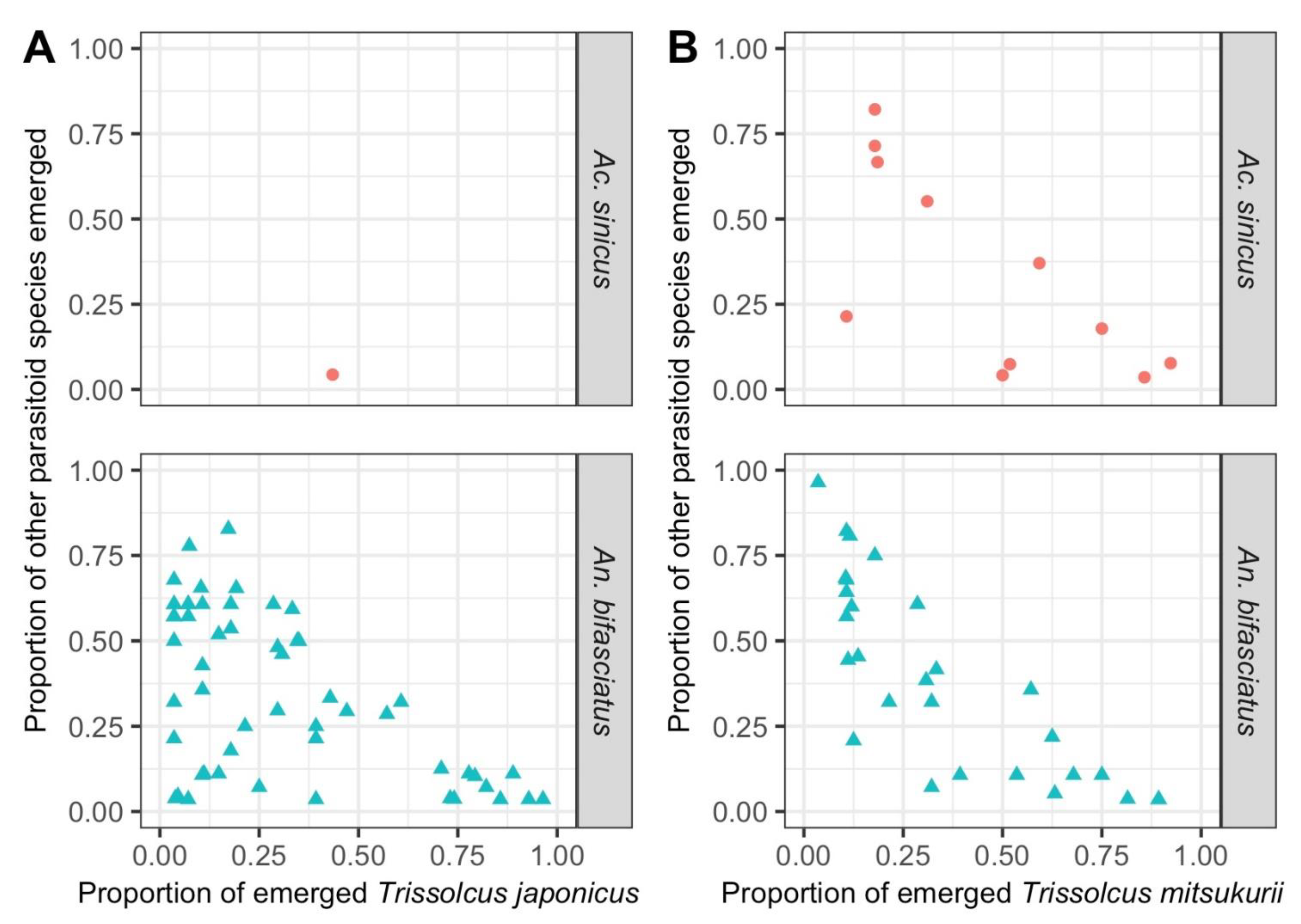
3.2. Parasitoid Species Composition
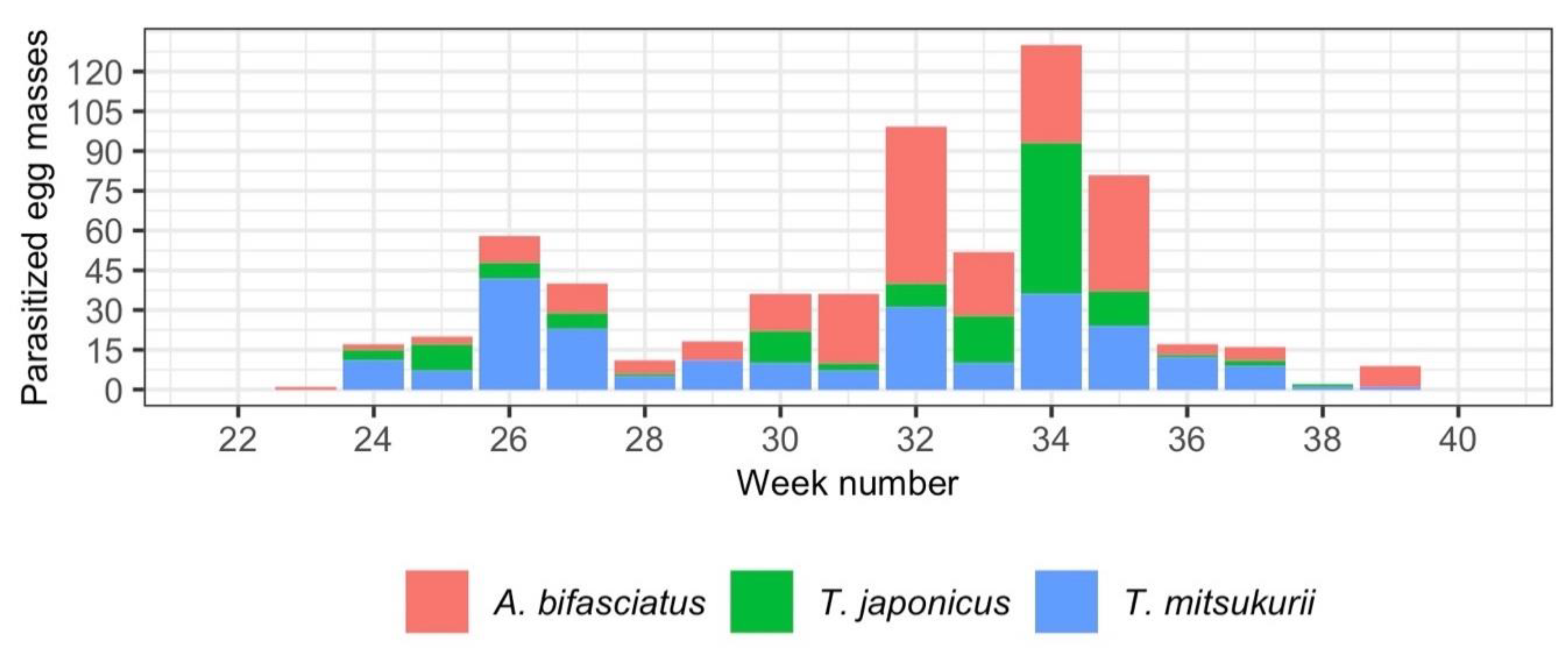
3.3. Parasitism Rate and Phenology
3.4. Habitat, Host Species and Distribution
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Faúndez, E.I.; Rider, D.A. The Brown Marmorated Stink Bug Halyomorpha halys (Stål, 1855) (Heteroptera: Pentatomidae) in Chile. Arq. Entomolóxicos 2017, 17, 305–307. [Google Scholar]
- Xu, J.; Fonseca, D.M.; Hamilton, G.C.; Hoelmer, K.A.; Nielsen, A.L. Tracing the Origin of US Brown Marmorated Stink Bugs, Halyomorpha halys. Biol. Invasions 2014, 16, 153–166. [Google Scholar] [CrossRef]
- Maistrello, L.; Dioli, P.; Dutto, M.; Volani, S.; Pasquali, S.; Gilioli, G. Tracking the Spread of Sneaking Aliens by Integrating Crowdsourcing and Spatial Modeling: The Italian Invasion of Halyomorpha halys. BioScience 2018, 68, 979–989. [Google Scholar] [CrossRef] [Green Version]
- Musolin, D.L.; Konjević, A.; Karpun, N.N.; Protsenko, V.Y.; Ayba, L.Y.; Saulich, A.K. Invasive Brown Marmorated Stink Bug Halyomorpha halys (Stål) (Heteroptera: Pentatomidae) in Russia, Abkhazia, and Serbia: History of Invasion, Range Expansion, Early Stages of Establishment, and First Records of Damage to Local Crops. Arthropod-Plant Interact. 2018, 12, 517–529. [Google Scholar] [CrossRef]
- Leskey, T.C.; Nielsen, A.L. Impact of the Invasive Brown Marmorated Stink Bug in North America and Europe: History, Biology, Ecology, and Management. Annu. Rev. Entomol. 2018, 63, 599–618. [Google Scholar] [CrossRef] [Green Version]
- Abram, P.K.; Hoelmer, K.A.; Acebes-Doria, A.; Andrews, H.; Beers, E.H.; Bergh, J.C.; Bessin, R.; Biddinger, D.; Botch, P.; Buffington, M.L.; et al. Indigenous Arthropod Natural Enemies of the Invasive Brown Marmorated Stink Bug in North America and Europe. J. Pest Sci. 2017, 90, 1009–1020. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, F.; Gariepy, T.; Mason, P.; Gillespie, D.; Talamas, E.; Haye, T. Seasonal Parasitism and Host Specificity of Trissolcus japonicus in Northern China. J. Pest Sci. 2017, 90, 1127–1141. [Google Scholar] [CrossRef] [Green Version]
- Conti, E.; Avila, G.; Barratt, B.; Cingolani, F.; Colazza, S.; Guarino, S.; Hoelmer, K.; Laumann, R.A.; Maistrello, L.; Martel, G.; et al. Biological Control of Invasive Stink Bugs: Review of Global State and Future Prospects. Entomol. Exp. Et Appl. 2020, eea.12967. [Google Scholar] [CrossRef]
- van Lenteren, J.C.; Bolckmans, K.; Köhl, J.; Ravensberg, W.J.; Urbaneja, A. Biological Control Using Invertebrates and Microorganisms: Plenty of New Opportunities. BioControl 2018, 63, 39–59. [Google Scholar] [CrossRef] [Green Version]
- EPPO. PM 6/2 (3) Import and Release of Non-Indigenous Biological Control Agents. EPPO Bull. 2014, 44, 320–329. [Google Scholar] [CrossRef]
- Food Agriculture Organization of the United Nations (FAO); International Plant Protection Convention (I.P.P.C.) (Eds.) Pest Risk Analysis for Quarantine Pests; International Standard for Phytosanitary Measures 11. Available online: https://www.ippc.int/en/publications/639/ (accessed on 26 June 2019).
- Hajek, A.E.; Hurley, B.P.; Kenis, M.; Garnas, J.R.; Bush, S.J.; Wingfield, M.J.; van Lenteren, J.C.; Cock, M.J.W. Exotic Biological Control Agents: A Solution or Contribution to Arthropod Invasions? Biol. Invasions 2016, 18, 953–969. [Google Scholar] [CrossRef] [Green Version]
- Van Lenteren, J.C.; Bale, J.; Bigler, F.; Hokkanen, H.M.T.; Loomans, A.J.M. Assessing risks of releasing exotic biological control agents of arthropod pests. Annu. Rev. Entomol. 2006, 51, 609–634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haye, T.; Fischer, S.; Zhang, J.; Gariepy, T. Can Native Egg Parasitoids Adopt the Invasive Brown Marmorated Stink Bug, Halyomorpha halys (Heteroptera: Pentatomidae), in Europe? J. Pest Sci. 2015, 88, 693–705. [Google Scholar] [CrossRef]
- Costi, E.; Haye, T.; Maistrello, L. Surveying Native Egg Parasitoids and Predators of the Invasive Halyomorpha halys in Northern Italy. J. Appl. Entomol. 2019, 143, 299–307. [Google Scholar] [CrossRef]
- Stahl, J.; Babendreier, D.; Marazzi, C.; Caruso, S.; Costi, E.; Maistrello, L.; Haye, T. Can Anastatus bifasciatus Be Used for Augmentative Biological Control of the Brown Marmorated Stink Bug in Fruit Orchards? Insects 2019, 10, 108. [Google Scholar] [CrossRef] [Green Version]
- Maistrello, L.; Costi, E.; Di Bella, E.; Vaccari, G.; Caruso, S.; Casoli, L.; Preti, M.; Landi, M.; Bombardini, E.; Moretti, C.; et al. Inundative releases of the native parasitoid Anastatus bifasciatus for the control of Halyomorpha halys in Italy. In Proceedings of the Atti Giornate Fitopatologiche 2020, Bologna, Italy, 3 November 2020; pp. 167–176. [Google Scholar]
- Talamas, E.J.; Herlihy, M.; Dieckhoff, C.; Hoelmer, K.A.; Buffington, M.; Bon, M.-C.; Weber, D.C. Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae) Emerges in North America. J. Hymenopt. Res. 2015, 43, 119–128. [Google Scholar] [CrossRef] [Green Version]
- Sabbatini Peverieri, G.; Talamas, E.; Bon, M.C.; Marianelli, L.; Bernardinelli, I.; Malossini, G.; Benvenuto, L.; Roversi, P.F.; Hoelmer, K. Two Asian Egg Parasitoids of Halyomorpha halys (Stål) (Hemiptera, Pentatomidae) Emerge in Northern Italy: Trissolcus mitsukurii (Ashmead) and Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae). J. Hymenopt. Res. 2018, 67, 37–53. [Google Scholar] [CrossRef]
- Moraglio, S.T.; Tortorici, F.; Pansa, M.G.; Castelli, G.; Pontini, M.; Scovero, S.; Visentin, S.; Tavella, L. A 3-Year Survey on Parasitism of Halyomorpha halys by Egg Parasitoids in Northern Italy. J. Pest Sci. 2020, 93, 183–194. [Google Scholar] [CrossRef]
- Stahl, J.; Tortorici, F.; Pontini, M.; Bon, M.C.; Hoelmer, K.; Marazzi, C.; Tavella, L.; Haye, T. First Discovery of Adventive Populations of Trissolcus japonicus in Europe. J. Pest Sci. 2019, 92, 371–379. [Google Scholar] [CrossRef] [Green Version]
- Scaccini, D.; Falagiarda, M.; Tortorici, F.; Martinez-Sañudo, I.; Tirello, P.; Reyes-Domínguez, Y.; Gallmetzer, A.; Tavella, L.; Zandigiacomo, P.; Duso, C.; et al. An Insight into the Role of Trissolcus mitsukurii as Biological Control Agent of Halyomorpha Halys in Northeastern Italy. Insects 2020, 11, 306. [Google Scholar] [CrossRef]
- Zapponi, L.; Bon, M.C.; Fouani, J.M.; Anfora, G.; Schmidt, S.; Falagiarda, M. Assemblage of the Egg Parasitoids of the Invasive Stink Bug Halyomorpha halys: Insights on Plant Host Associations. Insects 2020, 11, 588. [Google Scholar] [CrossRef]
- Milnes, J.M.; Wiman, N.G.; Talamas, E.J.; Brunner, J.F.; Hoelmer, K.A.; Buffington, M.L.; Beers, E.H. Discovery of an Exotic Egg Parasitoid of the Brown Marmorated Stink Ug, Halyomorpha halys (Stål) in the Pacific Northwest. Proc. Entomol. Soc. Wash. 2016, 118, 466–470. [Google Scholar] [CrossRef]
- Moraglio, S.T.; Tortorici, F.; Giromini, D.; Pansa, M.G.; Visentin, S.; Tavella, L. Field collection of egg parasitoids of Pentatomidae and Scutelleridae in Northwest Italy and their efficacy in parasitizing Halyomorpha halys under laboratory conditions. Entomol. Exp. Et Appl. 2021, 169, 52–63. [Google Scholar] [CrossRef]
- Bin, F.; Vinson, S.B. Efficacy Assesment in Egg Parasitoids (Hymenoptera): Proposal for a Unified Terminology. In Proceedings of the Trichogramma and Other Egg Parasitoids 3rd International Symposium on Le Colloques de l’INRA, San Antonio, TX, USA, 23–27 September 1990; Wajnberg, E., Vinson, S.B., Eds.; Institut National de la Recherche Agronomique: Paris, France, 1991; Volume 56, p. 246. [Google Scholar]
- Askew, R.R.; Nieves-Aldrey, J.L. Further Observations on Eupelminae (Hymenoptera, Chalcidoidea, Eupelmidae) in the Iberian Peninsula and Canary Islands, Including Descriptions of New Species. Graellsia 2004, 60, 27–39. [Google Scholar]
- Sabbatini Peverieri, G.; Mitroiu, M.-D.; Bon, M.-C.; Balusu, R.; Benvenuto, L.; Bernardinelli, I.; Fadamiro, H.; Falagiarda, M.; Fusu, L.; Grove, E.; et al. Surveys of Stink Bug Egg Parasitism in Asia, Europe and North America, Morphological Taxonomy, and Molecular Analysis Reveal the Holarctic Distribution of Acroclisoides sinicus (Huang & Liao) (Hymenoptera, Pteromalidae). J. Hymenopt. Res. 2019, 74, 123–151. [Google Scholar] [CrossRef] [Green Version]
- Talamas, E.J.; Buffington, M.L.; Hoelmer, K. Revision of Palearctic Trissolcus Ashmead (Hymenoptera, Scelionidae). J. Hymenopt. Res. 2017, 56, 79–261. [Google Scholar] [CrossRef] [Green Version]
- Tortorici, F.; Talamas, E.J.; Moraglio, S.T.; Pansa, M.G.; Asadi-Farfar, M.; Tavella, L.; Caleca, V. A Morphological, Biological and Molecular Approach Reveals Four Cryptic Species of Trissolcus Ashmead (Hymenoptera, Scelionidae), Egg Parasitoids of Pentatomidae (Hemiptera). J. Hymenopt. Res. 2019, 73, 153–200. [Google Scholar] [CrossRef] [Green Version]
- Kozlov, M.A.; Kononova, S.V. Telenominae of the Fauna of the USSR (Hymenoptera, Scelionidae, Telenominae); “Nauka”, Leningradskoe Otdelenie: Leningrad, Russia, 1983. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.r-project.org/ (accessed on 1 February 2021).
- Viggiani, G. Lotta Biologica e Integrata Nella Difesa Fitosanitaria; Liguori Editore: Napoli, Italy, 1994; pp. 1–517. [Google Scholar]
- Konopka, J.K.; Haye, T.; Gariepy, T.D.; McNeil, J.N. Possible coexistence of native and exotic parasitoids and their impact on control of Halyomorpha halys. J. Pest Sci. 2017, 90, 1119–1125. [Google Scholar] [CrossRef]
- Konopka, J.K.; Gariepy, T.D.; Haye, T.; Zhang, J.; Rubin, B.D.; McNeil, J.N. Exploitation of Pentatomids by Native Egg Parasitoids in the Native and Introduced Ranges of Halyomorpha halys: A Molecular Approach Using Sentinel Egg Masses. J. Pest Sci. 2019, 92, 609–619. [Google Scholar] [CrossRef]
- Costi, E.; Wong, W.H.L.; Cossentine, J.; Acheampong, S.; Maistrello, L.; Haye, T.; Talamas, E.J.; Abram, P.K. Variation in Levels of Acceptance, Developmental Success, and Abortion of Halyomorpha halys Eggs by Native North American Parasitoids. Biol. Control 2020, 151, 104396. [Google Scholar] [CrossRef]
- Cornell, H.V.; Hawkins, B.A. Accumulation of Native Parasitoid Species on Introduced Herbivores: A Comparison of Hosts as Natives and Hosts as Invaders. Am. Nat. 1993, 141, 847–865. [Google Scholar] [CrossRef] [PubMed]
- Arakawa, R.; Namura, Y. Effects of Temperature on Development of Three Trissolcus Spp. (Hymenoptera: Scelionidae), Egg Parasitoids of the Brown Marmorated Stink Bug, Halyomorpha halys (Hemiptera: Pentatomidae). Entomol. Sci. 2002, 5, 215–218. [Google Scholar]
- Jones, A.L.; Jennings, D.E.; Hooks, C.R.R.; Shrewsbury, P.M. Sentinel eggs underestimate rates of parasitism of the exotic brown marmorated stink bug, Halyomorpha halys. Biol. Control 2014, 78, 61–66. [Google Scholar] [CrossRef]
- Hepler, J.R.; Athey, K.; Enicks, D.; Abram, P.K.; Gariepy, T.D.; Talamas, E.J.; Beers, E. Hidden Host Mortality from an Introduced Parasitoid: Conventional and Molecular Evaluation of Non-Target Risk. Insects 2020, 11, 822. [Google Scholar] [CrossRef] [PubMed]
- Haye, T.; Moraglio, S.T.; Stahl, J.; Visentin, S.; Gregorio, T.; Tavella, L. Fundamental Host Range of Trissolcus japonicus in Europe. J. Pest Sci. 2020, 93, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Sabbatini Peverieri, G.; Giovannini, L.; Benvenuti, C.; Madonni, L.; Hoelmer, K.; Roversi, P.F. Characteristics of the Meconia of European Egg Parasitoids of Halyomorpha halys. J. Hymenopt. Res. 2020, 77, 187–201. [Google Scholar] [CrossRef]
- Colazza, S.; Bin, F. Efficiency of Trissolcus basalis (Hymenoptera: Scelionidae) as an Egg Parasitoid of Nezara viridula (Heteroptera: Pentatomidae) in Central Italy. Environ. Entomol. 1995, 24, 1703–1707. [Google Scholar] [CrossRef]
- McEvoy, P.B. Theoretical Contributions to Biological Control Success. BioControl 2018, 63, 87–103. [Google Scholar] [CrossRef]
- Barratt, B.I.P.; Moran, V.C.; Bigler, F.; van Lenteren, J.C. The Status of Biological Control and Recommendations for Improving Uptake for the Future. BioControl 2018, 63, 155–167. [Google Scholar] [CrossRef] [Green Version]
- Sheppard, A.W.; Shaw, R.H.; Sforza, R. Top 20 Environmental Weeds for Classical Biological Control in Europe: A Review of Opportunities, Regulations and Other Barriers to Adoption. Weed Res. 2006, 46, 93–117. [Google Scholar] [CrossRef]
- Buffington, M.L.; Talamas, E.J.; Hoelmer, K.A. Team Trissolcus: Integrating Taxonomy and Biological Control to Combat the Brown Marmorated Stink Bug. Am. Entomol. 2018, 64, 224–232. [Google Scholar] [CrossRef]
- Peixoto, L.; Allen, G.R.; Ridenbaugh, R.D.; Quarrell, S.R.; Withers, T.M.; Sharanowski, B.J. When Taxonomy and Biological Control Researchers Unite: Species Delimitation of Eadya Parasitoids (Braconidae) and Consequences for Classical Biological Control of Invasive Paropsine Pests of Eucalyptus. PLoS ONE 2018, 13, e0201276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Liu, Y.; Shi, M.; Huang, J.; Chen, X. Parasitoid Wasps as Effective Biological Control Agents. J. Integr. Agric. 2019, 18, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Avila, G.A.; Charles, J.G. Modelling the Potential Geographic Distribution of Trissolcus japonicus: A Biological Control Agent of the Brown Marmorated Stink Bug, Halyomorpha Halys. BioControl 2018, 63, 505–518. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Species | N. EM | N. PEM | Species of Parasitoids | Species of Hyperparasitoids |
|---|---|---|---|---|
| Carpocoris spp. | 8 | 4 | Telenomus sp. (4) | |
| Dolycoris baccarum | 22 | 7 | Anastatus bifasciatus (1), Telenomus sp. (3), Trissolcus belenus (1), Trissolcus mitsukurii (2) | |
| Eurydema spp. | 4 | 2 | Trissolcus viktorovi (2) | |
| Halyomorpha halys | 4348 | 974 | Anastatus bifasciatus (624), Telenomus sp. (25), Trissolcus basalis (20), Trissolcus belenus (4), Trissolcus cultratus (14), Trissolcus japonicus (152), Trissolcus mitsukurii (325), Trissolcus semistriatus (4), Undet. (20) | Acroclisoides sinicus (69) |
| Nezara viridula | 153 | 31 | Anastatus bifasciatus (17), Trissolcus basalis (11), Trissolcus mitsukurii (3) | Acroclisoides sinicus (1) |
| Palomena prasina | 74 | 38 | Anastatus bifasciatus (11), Telenomus sp. (7), Telenomus turesis (2), Trissolcus cultratus (11), Trissolcus japonicus (5), Trissolcus mitsukurii (2) | |
| Pentatoma rufipes | 6 | 2 | Trissolcus cultratus (2) | |
| Piezodorus lituratus | 7 | 1 | Trissolcus semistriatus (1) | |
| Rhaphigaster nebulosa | 11 | 2 | Anastatus bifasciatus (1), Telenomus sp. (1) |
| A. Trissolcus japonicus | ||||||
| Country | Region/Canton | N. EM | N. E | PEM (%) | PE (%) | PR (Mean ± SE) |
| Italy | Piedmont | 1788 | 46,824 | 4.03 | 2.98 | 89.43 ± 2.97 |
| Veneto | 669 | 16,966 | 0.45 | 0.29 | 100.00 ± 0.00 | |
| Switzerland | Basel | 26 | 625 | 53.85 | 50.88 | 80.25 ± 6.95 |
| Zürich | 25 | 673 | 32.00 | 25.41 | 71.43 ± 10.19 | |
| B. Trissolcus mitsukurii | ||||||
| Country | Region/Canton | N. EM | N. E | PEM (%) | PE (%) | PR (Mean ± SE) |
| Italy | Emilia Romagna | 629 | 16,707 | 3.66 | 2.75 | 97.40 ± 1.68 |
| Friuli-Venezia Giulia | 589 | 13,874 | 20.20 | 14.49 | 96.21 ± 1.38 | |
| Piedmont | 1788 | 46,824 | 0.17 | 0.09 | 91.23 ± 8.77 | |
| Trentino-Alto Adige | 585 | 15,506 | 5.98 | 4.15 | 96.70 ± 2.92 | |
| Veneto | 669 | 16,966 | 6.88 | 4.66 | 88.59 ± 3.27 | |
| C. Anastatus bifasciatus | ||||||
| Country | Region/Canton | N. EM | N. E | PEM (%) | PE (%) | PR (Mean ± SE) |
| Italy | Emilia Romagna | 629 | 16,707 | 19.55 | 2.36 | 66.06 ± 2.78 |
| Friuli-Venezia Giulia | 589 | 13,874 | 0.85 | 0.35 | 44.89 ± 19.54 | |
| Piedmont | 1788 | 46,824 | 15.72 | 7.79 | 73.97 ± 1.92 | |
| Trentino-Alto Adige | 585 | 15,506 | 11.97 | 5.62 | 76.09 ± 3.69 | |
| Veneto | 669 | 16,966 | 1.05 | 0.62 | 68.32 ± 15.15 | |
| Switzerland | Basel | 26 | 625 | 3.85 | 3.36 | 75.00 |
| Zürich | 25 | 673 | 4.00 | 0.15 | 3.57 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zapponi, L.; Tortorici, F.; Anfora, G.; Bardella, S.; Bariselli, M.; Benvenuto, L.; Bernardinelli, I.; Butturini, A.; Caruso, S.; Colla, R.; et al. Assessing the Distribution of Exotic Egg Parasitoids of Halyomorpha halys in Europe with a Large-Scale Monitoring Program. Insects 2021, 12, 316. https://doi.org/10.3390/insects12040316
Zapponi L, Tortorici F, Anfora G, Bardella S, Bariselli M, Benvenuto L, Bernardinelli I, Butturini A, Caruso S, Colla R, et al. Assessing the Distribution of Exotic Egg Parasitoids of Halyomorpha halys in Europe with a Large-Scale Monitoring Program. Insects. 2021; 12(4):316. https://doi.org/10.3390/insects12040316
Chicago/Turabian StyleZapponi, Livia, Francesco Tortorici, Gianfranco Anfora, Simone Bardella, Massimo Bariselli, Luca Benvenuto, Iris Bernardinelli, Alda Butturini, Stefano Caruso, Ruggero Colla, and et al. 2021. "Assessing the Distribution of Exotic Egg Parasitoids of Halyomorpha halys in Europe with a Large-Scale Monitoring Program" Insects 12, no. 4: 316. https://doi.org/10.3390/insects12040316
APA StyleZapponi, L., Tortorici, F., Anfora, G., Bardella, S., Bariselli, M., Benvenuto, L., Bernardinelli, I., Butturini, A., Caruso, S., Colla, R., Costi, E., Culatti, P., Di Bella, E., Falagiarda, M., Giovannini, L., Haye, T., Maistrello, L., Malossini, G., Marazzi, C., ... Sabbatini-Peverieri, G. (2021). Assessing the Distribution of Exotic Egg Parasitoids of Halyomorpha halys in Europe with a Large-Scale Monitoring Program. Insects, 12(4), 316. https://doi.org/10.3390/insects12040316