
Optimal Conditions for Diapause Survival of Aprostocetus fukutai, an Egg Parasitoid for Biological Control of Anoplophora chinensis


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Plants
2.2. Diapause Survival on Potted Trees under Semi-Natural Conditions
2.3. Optimal Humidity
2.4. Optimal Temperature Regimes
2.5. Data Analysis
3. Results
3.1. Diapause Survival on Potted Trees under Semi-Natural Conditions
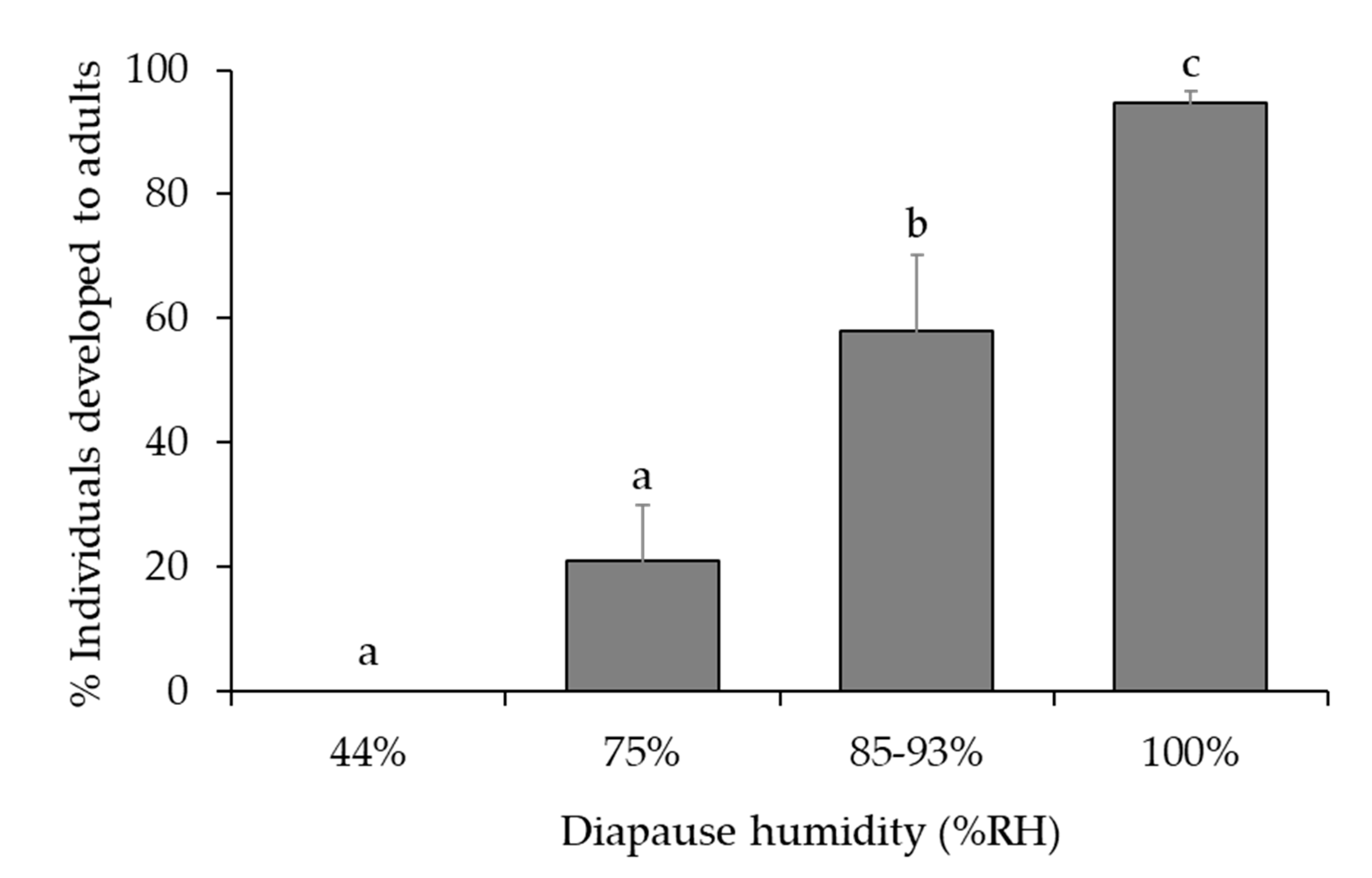
3.2. Optimal Humidity
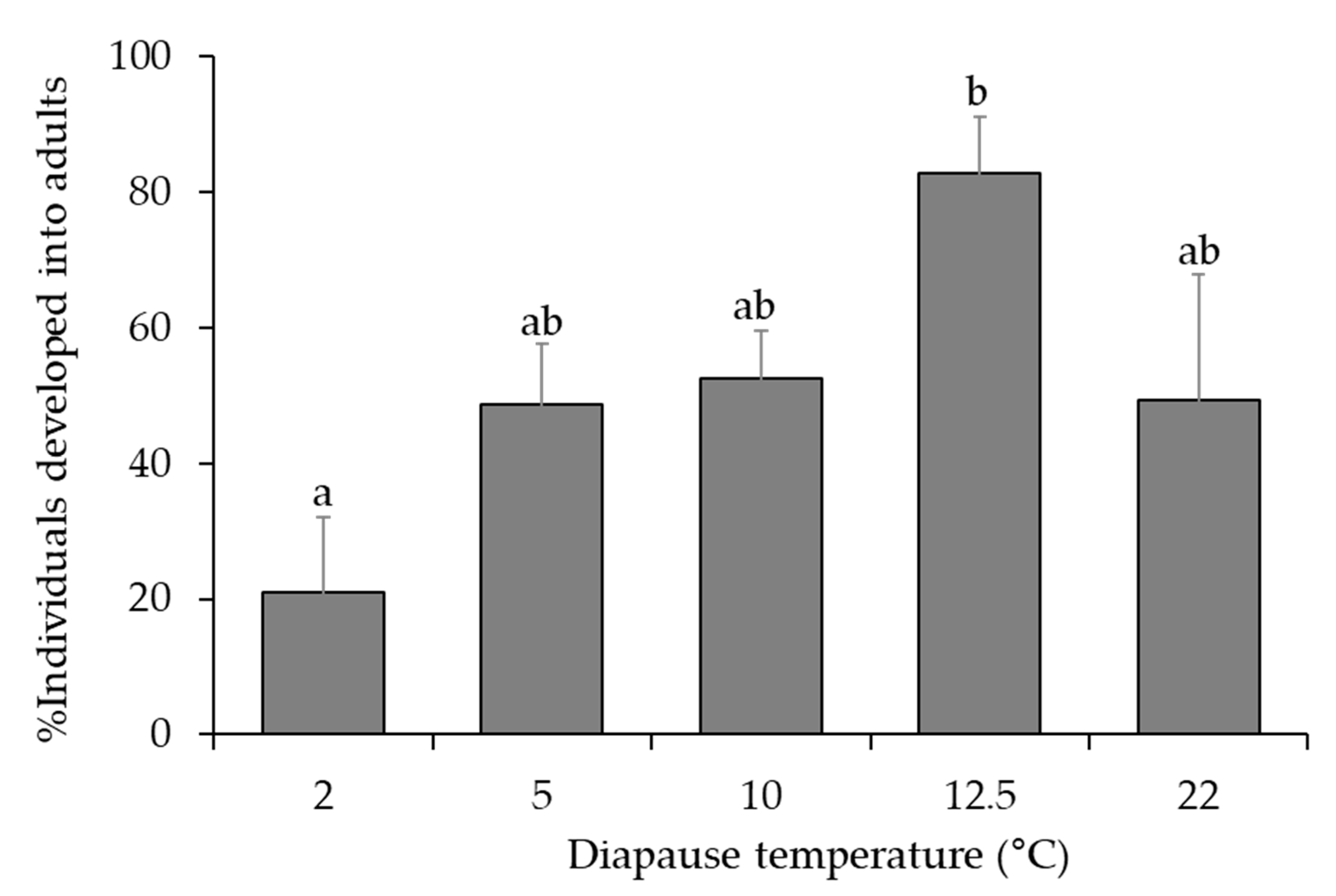
3.3. Optimal Temperature Regimes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tauber, M.J.; Tauber, C.A. Insect seasonality: Diapause maintenance, termination, and post diapause development. Annu. Rev. Entomol. 1976, 21, 81–107. [Google Scholar] [CrossRef]
- Tauber, M.J.; Tauber, C.A.; Masaki, S. Seasonal Adaptations of Insects; Oxford University Press: Oxford, UK, 1986. [Google Scholar]
- Hopper, K.R. Risk-spreading and bet-hedging in insect population biology. Annu. Rev. Entomol. 1999, 44, 535–560. [Google Scholar] [CrossRef]
- Koštal, V.; Stětina, T.; Poupardin, R.; Korbelova, J.; Bruce, A.W. Conceptual framework of the eco-physiological phases of insect diapause development justified by transcriptomic profiling. Proc. Natl. Acad. Sci. USA 2017, 114, 8532–8537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, J.J.; Larson, K.M. Effects of chilling on diapause development and reproductive fitness of two congeneric species of encyrtid parasitoids (Hymenoptera: Encyrtidae) of the emerald ash borer. Biol. Control 2019, 134, 163–169. [Google Scholar] [CrossRef]
- Duan, J.J.; Bauer, L.S.; van Driesche, R.; Schmude, J.M.; Petrice, T.; Chandler, J.L.; Elkinton, J. Effects of extreme low winter temperatures on the overwintering survival of the introduced larval parasitoids Spathius galinae and Tetrastichus planipennisi: Implications for biological control of emerald ash borer in North America. J. Econ. Entomol. 2020, 113, 1145–1151. [Google Scholar] [CrossRef] [Green Version]
- Denlinger, D.L. Regulation of diapause. Annu. Rev. Entomol. 2002, 47, 93–122. [Google Scholar] [CrossRef] [PubMed]
- Hahn, D.A.; Denlinger, D.L. Energetics and insect diapause. Annu. Rev. Entomol. 2011, 56, 103–121. [Google Scholar] [CrossRef] [PubMed]
- Hance, T.; van Baaren, J.; Vernon, P.; Boivin, G. Impact of extreme temperatures on parasitoids in a climate change perspective. Annu. Rev. Entomol. 2007, 52, 107–126. [Google Scholar] [CrossRef]
- Koštal, V. Eco-physiological phases of insect diapause. J. Insect Physiol. 2006, 52, 113–127. [Google Scholar] [CrossRef]
- Hodek, I.; Hodkova, M. Multiple roles of temperature during insect diapause: A review. Entomol. Exp. Appl. 1988, 49, 153–165. [Google Scholar] [CrossRef]
- Haack, R.A.; Hérard, F.; Sun, J.; Turgeon, J.J. Managing invasive populations of Asian longhorned beetle and citrus longhorned beetle: A worldwide perspective. Annu. Rev. Entomol. 2010, 55, 521–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hérard, F.; Maspero, M. History of discoveries and management of the citrus longhorned beetle, Anoplophora chinensis, in Europe. J. Pest. Sci. 2019, 92, 117–130. [Google Scholar] [CrossRef]
- Aukema, J.E.; McCullough, D.G.; Von Holle, B.; Liebhold, A.M.; Britton, K.; Frankel, S.J. Historical accumulation of non-indigenous forest pests in the continental United States. BioScience 2010, 60, 886–897. [Google Scholar] [CrossRef]
- Haack, R.A. Exotic bark- and wood-boring Coleoptera in the United States: Recent establishments and interceptions. Can. J. For. Res. 2006, 36, 269–288. [Google Scholar] [CrossRef]
- Pschorn-Walcher, H. Biological control of forest insects. Annu. Rev. Entomol. 1977, 22, 1–22. [Google Scholar] [CrossRef]
- Kenis, M.; Hurley, B.P.; Hajek, A.E.; Cock, M.J.W. Classical biological control of insect pests of trees: Facts and figures. Biol. Invasions 2017, 19, 3401–3417. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.G.; Wang, X.; Kenis, M.; Cao, L.M.; Duan, J.J.; Gould, J.; Hoelmer, K.M. Exploring the potential for novel association biological control of invasive woodboring beetles using generalist parasitoids. BioControl 2021, 66, 97–112. [Google Scholar] [CrossRef]
- Gould, J.R.; Aflague, B.; Murphy, T.C.; McCartin, L.; Elkinton, J.S.; Rim, K.; Duan, J.J. Collecting nontarget wood-boring insects for host-specificity testing of natural enemies of Cerambycids: A case study of Dastarcus helophoroides (Coleoptera: Bothrideridae), a parasitoid of the Asian longhorned beetle (Coleoptera: Cerambycidae). Environ. Entomol. 2018, 47, 1440–1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brabbs, T.; Collins, D.; Hérard, F.; Maspero, M.; Eyre, D. Prospects for the use of biological control agents against Anoplophora in Europe. Pest. Manag. Sci. 2015, 71, 7–14. [Google Scholar] [CrossRef]
- Golec, J.R.; Aparicio, E.M.; Wang, X.G.; Duan, J.J.; Fuester, R.W.; Tatman, D.; Kula, R.R. Cerambycid communities and their associated hymenopteran parasitoids from major hardwood trees in Delaware, USA: Implications for biocontrol of invasive longhorned beetles. Environ. Entomol. 2020, 49, 370–382. [Google Scholar] [CrossRef]
- Wang, X.G.; Aparicio, E.M.; Murphy, T.C.; Duan, J.J.; Elkinton, J.S.; Gould, J.R. Assessing the host range of the North American parasitoid Ontsira mellipes: Potential for biological control of Asian longhorned beetle. Biol. Control 2019, 137, 104028. [Google Scholar] [CrossRef]
- Duan, J.J.; Aparicio, E.M.; Tatman, D.; Smith, M.T.; Luster, D.G. Potential new associations of North American parasitoids with the invasive Asian longhorned beetle (Coleoptera: Cerambycidae) for biological control. J. Econ. Entomol. 2015, 109, 669–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.G.; Aparicio, E.M. Reproductive traits of Ontsira mellipes (Hymenoptera: Braconidae), a North American parasitoid as a novel biological control agent for exotic Anoplophora glabripennis (Coleoptera: Cerambycidae). J. Econ. Entomol. 2020, 113, 2112–2119. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.G.; Aparicio, E.M.; Duan, J.J.; Gould, J.; Hoelmer, K.A. Optimizing parasitoid and host densities for efficient rearing of Ontsira mellipes (Hymenoptera: Braconidae) on Asian longhorned beetle (Coleoptera: Cerambycidae). Environ. Entomol. 2020, 49, 1041–1048. [Google Scholar] [CrossRef]
- Hanks, L.M.; Paine, T.D.; Millar, J.G. Tiny wasp helps protect eucalypts from eucalyptus longhorned borer. Cali. Agri. 1996, 50, 14–16. [Google Scholar] [CrossRef]
- Delvare, G.; Bon, M.-C.; Hérard, F.; Cocquempot, C.; Maspero, M.; Colombo, M. Description of Aprostocetus anoplophorae n. sp. (Hymenoptera: Eulophidae), a new egg parasitoid of the invasive pest Anoplophora chinensis (Förster) (Coleoptera: Cerambycidae). Ann. Soc. Entomol. Fr. 2004, 40, 227–233. [Google Scholar] [CrossRef]
- Colombo, M.; Limonta, L. Anoplophora malasiaca Thomson (Coleoptera: Cerambycidae: Lamiinae: Lamiini) in Europe. Boll. Zool. Agrar. Bachic. 2001, 33, 65–68. [Google Scholar]
- LaSalle, J.; Huang, D.W. Two new Eulophidae (Hymenoptera: Chalcidoidea) of economic importance from China. Bull. Entomol. Res. 1994, 84, 51–56. [Google Scholar] [CrossRef]
- He, X.; Huang, J.; Zeng, G.; Ye, J.; Gao, M. Bionomics of Aprostocetus fukutai Miwa et Sonan. For. Pest. Dis. 2000, 19, 11–13. [Google Scholar]
- Li, J.Q.; Wang, S.X.; Yang, Y.; Huang, D.Z.; Jin, Y.J.; Bai, Y. The study on oviposition, host recognition and discrimination behavior of Aprostocetus fukutai (Miwa et Sonan). For. Sci. 2006, 32, 447–452. [Google Scholar]
- Hérard, F.; Maspero, M.; Bon, M.-C. Accidental introduction into Italy and establishment of Aprostocetus fukutai (Hymenoptera: Eulophidae) in citrus longhorned beetle infestations. In Proceedings of the 5th International Symposium on Biological Control of Arthropods, Longkawi, Malaysia, 11–15 September 2017; Mason, P.G., Gillespie, D.R., Vincent, C., Eds.; CAB International: Wallingford, UK, 2017; pp. 15–17. [Google Scholar]
- Adachi, I. Reproductive biology of the white-spotted longicorn beetle, Anoplophora malasiaca Thomson (Coleoptera: Cerambycidae), in citrus trees. Appl. Entomol. Zool. 1988, 23, 256–264. [Google Scholar] [CrossRef] [Green Version]
- Keena, M.A. Laboratory rearing and handling of Cerambycids. In Cerambycidae of the World: Biology and Pest Management; Wang, Q., Ed.; Taylor & Francis Group: Boca Raton, FL, USA, 2017; pp. 253–284. [Google Scholar]
- Greenspan, L. Humidity fixed points of binary saturated aqueous solutions. J. Res. Natl. Bur. Stand. 1977, 81A, 89–96. [Google Scholar] [CrossRef]
- Ge, X.; Zong, S.; He, S.; Liu, Y.; Kong, X. Areas of China predicted to have a suitable climate for Anoplophora chinensis under a climate-warming scenario. Entomol. Exp. Appl. 2014, 153, 256–265. [Google Scholar] [CrossRef]
- Wang, X.G.; Messing, R.H. Fitness consequence of body size-dependent host species selection in a generalist ectoparasitoid. Behav. Ecol. Sociobiol. 2004, 56, 513–522. [Google Scholar] [CrossRef]
- Larson, K.M.; Duan, J.J. Differences in the reproductive biology and diapause of two congeneric species of egg parasitoids (Hymenoptera: Encyrtidae) from northeast Asia: Implications for biological control of the invasive emerald ash borer (Coleoptera: Buprestidae). Biol. Control 2016, 103, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Hoban, J.; Duan, J.J.; Hough-Goldstein, J. Effects of temperature and photoperiod on the reproductive biology and diapause of Oobius agrili (Hymenoptera: Encyrtidae), an egg parasitoid of emerald ash borer (Coleoptera: Buprestidae). Environ. Entomol. 2016, 45, 726–731. [Google Scholar] [CrossRef] [Green Version]
- Fatouros, N.E.; Cusumano, A.; Bin, F.; Polaszek, A.; van Lenteren, J.C. How to escape from insect egg parasitoids: A review of potential factors explaining parasitoid absence across the Insecta. Proc. R. Soc. B 2020, 287, 20200344. [Google Scholar] [CrossRef]
- Haack, R.A.; Keena, M.A.; Eyre, D. Life history and population dynamics of Cerambycidae. In Cerambycidae of the World, Biology and Management; Wang, Q., Ed.; Taylor & Francis Group: Boca Raton, FL, USA, 2017; pp. 71–94. [Google Scholar]
- Wang, J.; Zhou, Z. Studies on resistance mechanisms of poplar clones to Anoplophora glabripennis. J. Beijing For. Univ. 1994, 16, 49–53. [Google Scholar]
- Desurmont, G.A.; Donoghue, M.J.; Clement, W.L.; Agrawal, A.A. Evolutionary history predicts plant defense against an invasive pest. Proc. Natl. Acad. Sci. USA 2011, 108, 7070–7074. [Google Scholar] [CrossRef] [Green Version]
- Strangi, A.; Binazzi, F.; Peverieri, G.S.; Roversi, P.F. The Anoplophora chinensis (Forster) (Coleoptera: Cerambycidae: Lamiinae) outbreaks in Italy: A possible geographical origin. Redia 2017, 100, 175–179. [Google Scholar]
- Hougardy, E.; Hogg, B.N.; Wang, X.G.; Daane, K.M. Comparison of thermal performances of two Asian larval parasitoids of Drosophila suzukii. Biol. Control 2019, 136, 104000. [Google Scholar] [CrossRef]
- Keena, M.A.; Moore, P.M.; Bradford, G. Effects of temperature on Anoplophora chinensis (Coleoptera: Cerambycidae) adult survival, reproduction, and egg hatch. Forests 2021, 12, 432. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Humidity Conditions at 11 °C and 10L:14D 1,2 | ||||
| Humidity (% RH) | Humidity Control | No. of Parasitized Host Eggs | ||
| 44 | Saturated K2CO3 solution | 24 | ||
| 75 | Saturated NaCl solution | 54 | ||
| 85–93 3 | Saturated KCl or KNO3 solution | 59 | ||
| 100 | Distilled water | 72 | ||
| Temperature Regimes at 100% RH and Dark 1,2 | ||||
| First Temperature (°C) | Duration (month) | Second Temperature (°C) | Duration (month) | No. of Parasitized Host Eggs |
| 2 | 5 | 12.5 | 1 | 25 |
| 10 | 1 | 5 | 6 | 71 |
| 10 | 6 | 10 | 0 | 83 |
| 12.5 | 6 | 12.5 | 0 | 30 |
| 22 | All time | 22 | All time | 11 |
| Temperature Regime (°C) | Realized Clutch Size 1,2 | % Female Offspring 1,2 | Post-Diapause Developmental Time (days) 1,2 | Longevity (Days) 1,3 | Lifetime Fecundity 1,3 |
|---|---|---|---|---|---|
| 2 | 12.8 ± 1.0 a | 89.0 ± 2.5 a | 72.5 ± 1.9 a | 9.6 ± 1.1 a | 3.0 ± 0.5 a |
| 5 | 10.7 ± 0.8 a | 80.8 ± 3.0 a | 71.8 ± 1.0 a | ||
| 10 | 10.0 ± 0.6 a | 82.3 ± 2.6 a | 51.5 ± 0.8 b | ||
| 12.5 | 11.6 ± 0.8 a | 87.3 ± 2.5 a | 34.9 ± 0.5 c | 15.1 ± 2.2 b | 3.1 ± 0.4 a |
| 22 | 11.7 ± 0.5 a | 85.4 ± 1.4 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.; Ramualde, N.; Aparicio, E.M.; Maspero, M.; Duan, J.J.; Smith, L. Optimal Conditions for Diapause Survival of Aprostocetus fukutai, an Egg Parasitoid for Biological Control of Anoplophora chinensis. Insects 2021, 12, 535. https://doi.org/10.3390/insects12060535
Wang X, Ramualde N, Aparicio EM, Maspero M, Duan JJ, Smith L. Optimal Conditions for Diapause Survival of Aprostocetus fukutai, an Egg Parasitoid for Biological Control of Anoplophora chinensis. Insects. 2021; 12(6):535. https://doi.org/10.3390/insects12060535
Chicago/Turabian StyleWang, Xingeng, Nathalie Ramualde, Ellen M. Aparicio, Matteo Maspero, Jian J. Duan, and Lincoln Smith. 2021. "Optimal Conditions for Diapause Survival of Aprostocetus fukutai, an Egg Parasitoid for Biological Control of Anoplophora chinensis" Insects 12, no. 6: 535. https://doi.org/10.3390/insects12060535
APA StyleWang, X., Ramualde, N., Aparicio, E. M., Maspero, M., Duan, J. J., & Smith, L. (2021). Optimal Conditions for Diapause Survival of Aprostocetus fukutai, an Egg Parasitoid for Biological Control of Anoplophora chinensis. Insects, 12(6), 535. https://doi.org/10.3390/insects12060535