
Fermenting Bread Dough as a Cheap, Effective, Nontoxic, and Generic Attractant for Pest Snails and Slugs




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Dough Formulation
2.2. Laboratory Choice Bioassays with Cornu aspersum to Test the Attractiveness of Bread Dough
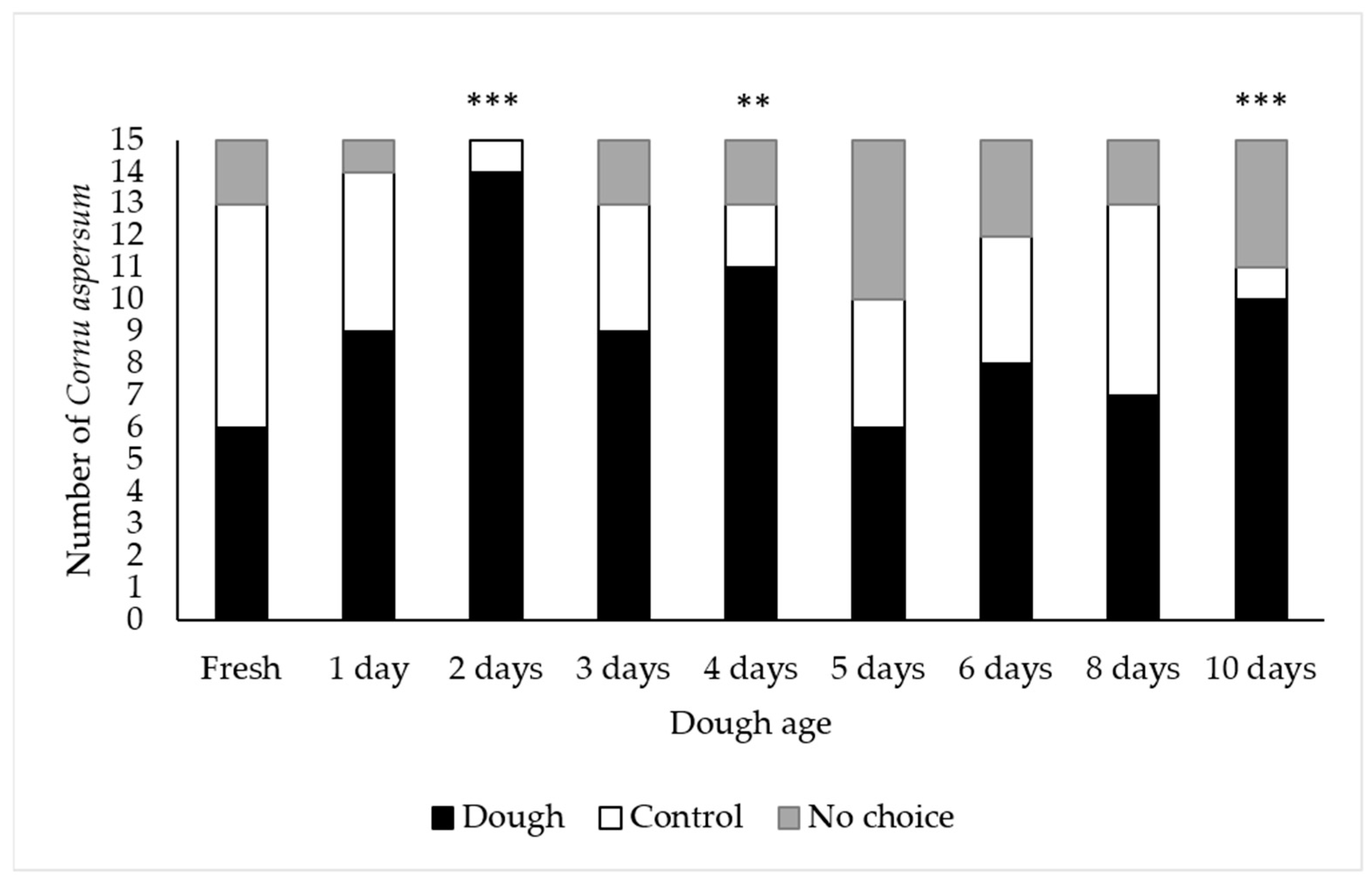
2.2.1. Attractiveness of Dough of Various Ages
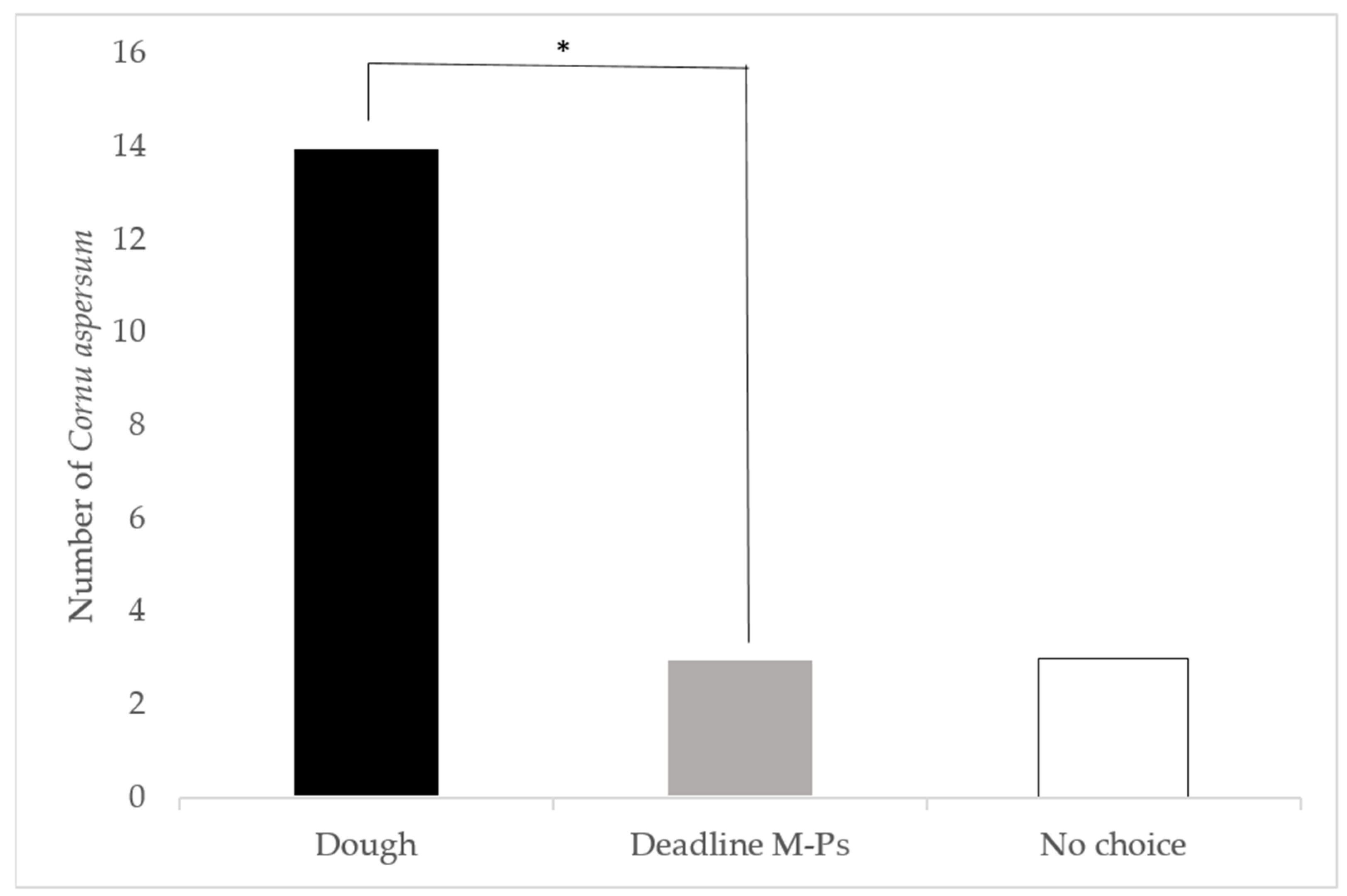
2.2.2. Attractiveness of Dough Aged for 2 Days versus Deadline® M-Ps™
2.3. Field Trials with Dough Aged for 2 Days
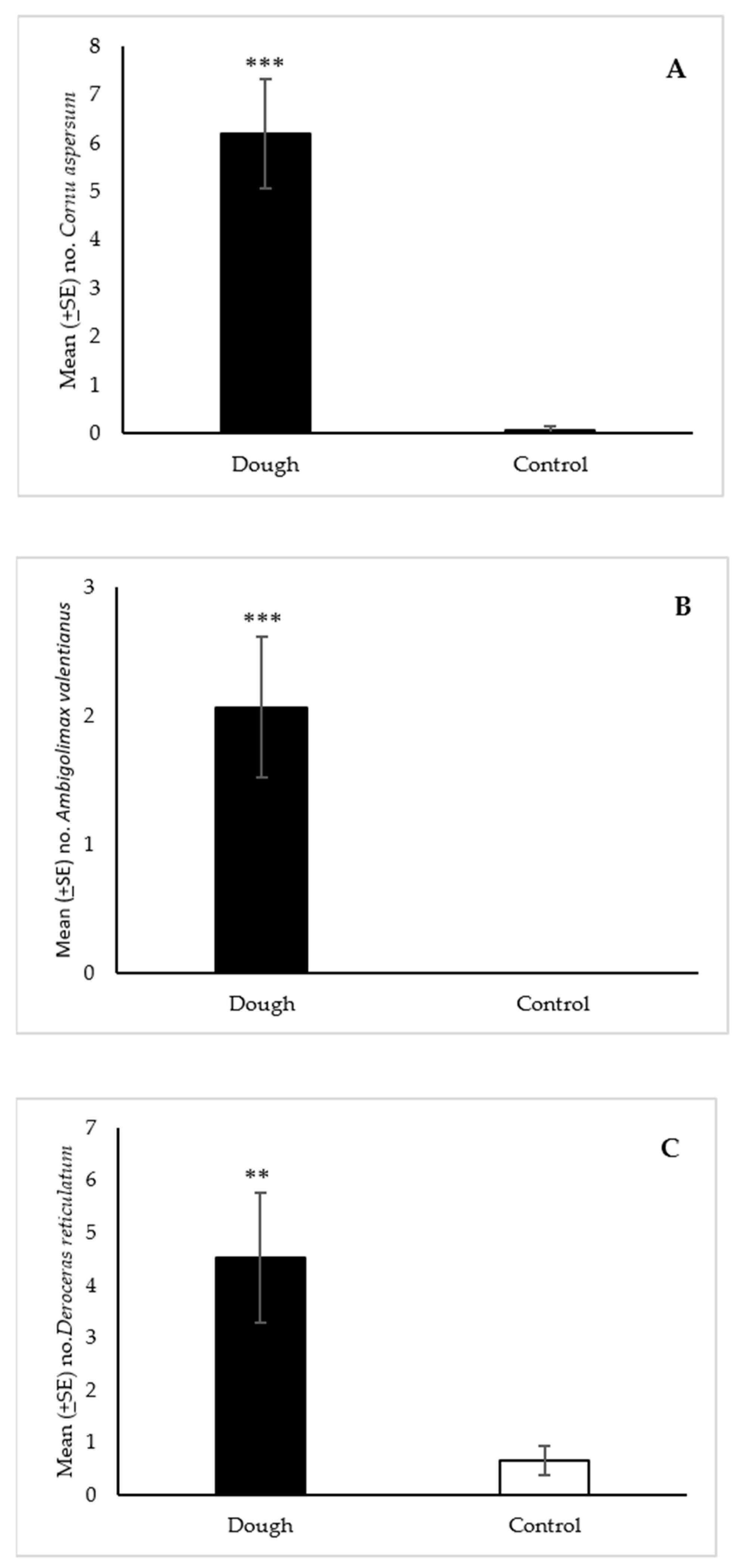
2.3.1. Field Trials with Cornu aspersum, Ambigolimax valentianus, and Deroceras reticulatum
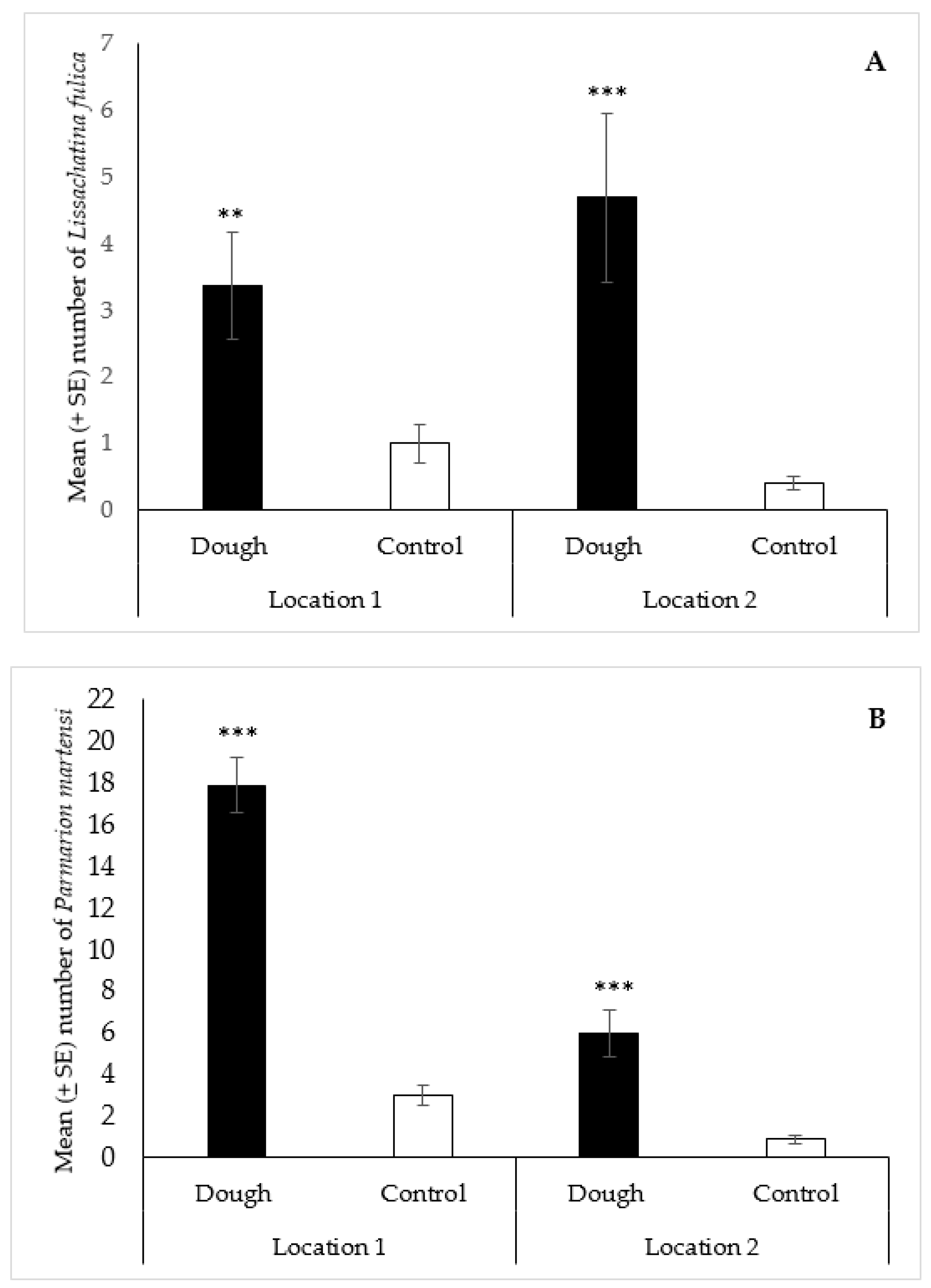
2.3.2. Lissachatina fulica and Parmarion martensi
2.3.3. Xerolenta obvia
Snailer® Traps
Petri Dish Bioassays
2.4. Effect of Aging on Dough Performance in the Field
2.5. Statistical Analysis
3. Results
3.1. Laboratory Choice Bioassays with Cornu aspersum to Test the Attractiveness of Bread Dough
3.1.1. Attractiveness of Dough of Various Ages
3.1.2. Attractiveness of Dough Aged for 2 Days versus Deadline® M-Ps™
3.2. Field Trials with Dough Aged for 2 Days
3.2.1. Cornu aspersum and Ambigolimax valentianus
3.2.2. Deroceras reticulatum
3.2.3. Lissachatina fulica and Parmarion martensi
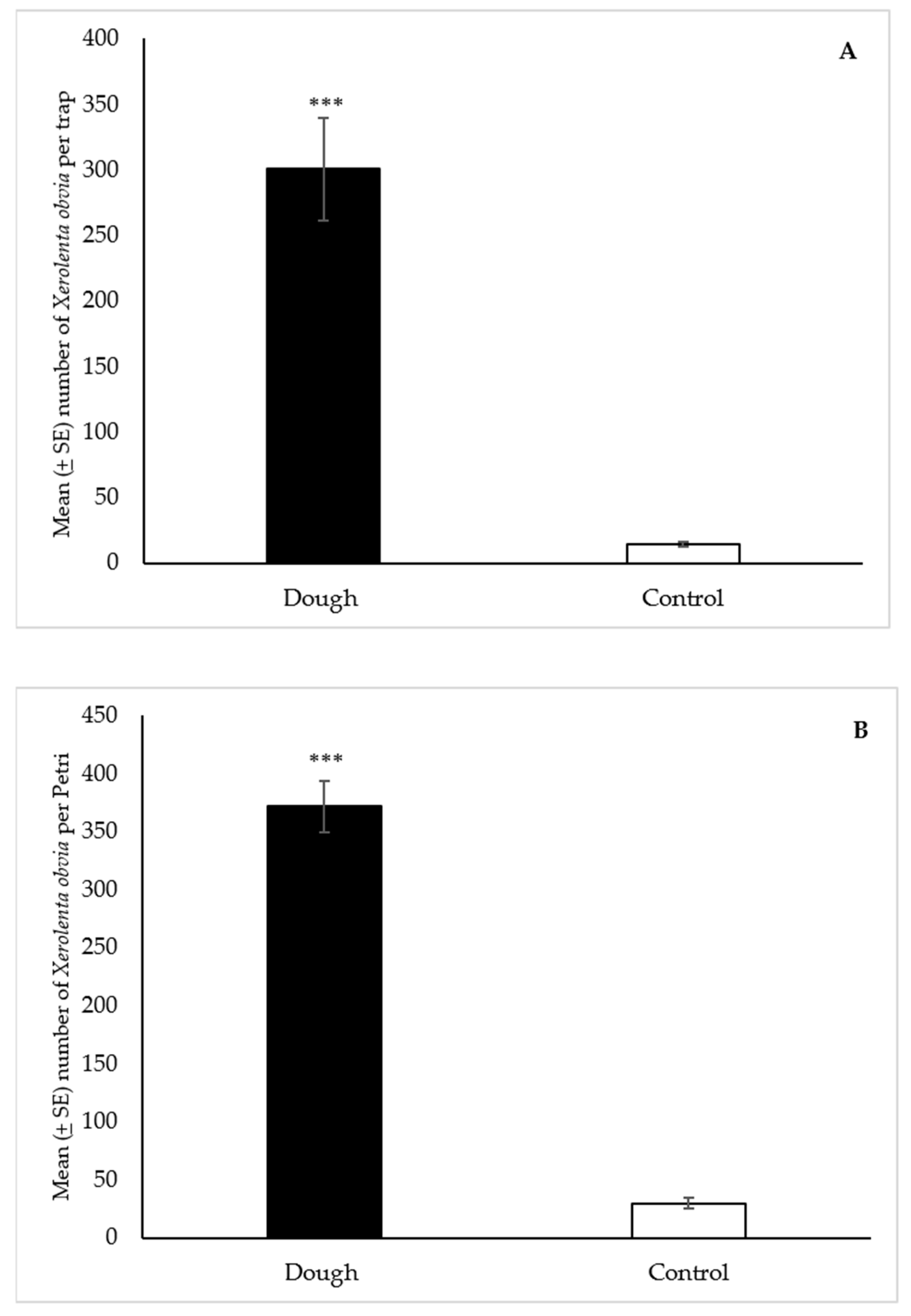
3.2.4. Xerolenta obvia
Snailer® Traps
Petri Dishes
3.3. Effect of Aging on Dough Performance in the Field
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barker, G.M. Molluscs as Crop Pests; CABI: Wallingford, UK, 2002; ISBN 978-0-85199-790-2. [Google Scholar]
- Cowie, R.H. Can snails ever be effective and safe biocontrol agents? Int. J. Pest Manag. 2001, 47, 23–40. [Google Scholar] [CrossRef]
- Bailey, S.E.R. Molluscicidal baits for control of terrestrial gastropods. In Molluscs as Crop Pests; Barker, G.M., Ed.; CABI: Wallingford, UK, 2002; pp. 33–54. ISBN 978-0-85199-790-2. [Google Scholar]
- Hammond, R.B.; Byers, R.A. Agriolimacidae and Arionidae as pests in conservation tillage soybean and maize cropping in North America. In Molluscs as Crop Pests; Barker, G.M., Ed.; CABI: Wallingford, UK, 2002; pp. 301–314. ISBN 978-0-85199-320-1. [Google Scholar]
- Henderson, I.; Triebskorn, R. Chemical control of terrestrial gastropods. In Molluscs as Crop Pests; Barker, G.M., Ed.; CABI: Wallingford, UK, 2002; pp. 1–31. ISBN 978-0-85199-790-2. [Google Scholar]
- Godan, D. Pest Slugs and Snails: Biology and Control; Springer: Berlin/Heidelberg, Germany, 1983; ISBN 978-3-642-68799-0. [Google Scholar]
- Barker, G.M.; Pottinger, R.P.; Lloyd, J.M.; Addison, P.J.; Firth, A.C.; Stewart, A.P. A Novel Bait Formulation for Slug and Snail Control; New Zealand Weed and Pest Control Society Inc.: Palmerstown, New Zealand, 1991; pp. 195–200. [Google Scholar]
- Rae, R.; Verdun, C.; Grewal, P.S.; Robertson, J.F.; Wilson, M.J. Biological control of terrestrial molluscs using Phasmarhabditis hermaphrodita—progress and prospects. Pest Manag. Sci. 2007, 63, 1153–1164. [Google Scholar] [CrossRef] [PubMed]
- Sakovich, N.J. Integrated management of Cantareus aspersus (Müller) (Helicidae) as a pest of citrus in California. In Molluscs as Crop Pests; Barker, G.M., Ed.; CABI: Wallingford, UK, 2002; pp. 353–360. ISBN 978-0-85199-320-1. [Google Scholar]
- Hammond, R.B.; Stinner, B.R. Seedcorn maggots (Diptera: Anthomyiidae) and slugs in conservation tillage systems in Ohio. J. Econ. Entomol. 1987, 80, 680–684. [Google Scholar] [CrossRef]
- Hirano, T.; Yamazaki, D.; Uchida, S.; Saito, T.; Chiba, S. First record of the slug species Semperula wallacei (Issel, 1874) (Gastropoda: Eupulmonata: Veronicellidae) in Japan. BioInvasions Rec. 2019, 8, 258–265. [Google Scholar] [CrossRef]
- Serniotti, E.N.; Guzmán, L.B.; Beltramino, A.A.; Vogler, R.E.; Rumi, A.; Peso, J.G. New distributional records of the exotic land snail Bradybaena similaris (Férussac, 1822) (Gastropoda, Bradybaenidae) in Argentina. BioInvasions Rec. 2019, 8, 301–313. [Google Scholar] [CrossRef]
- Vendetti, J.E.; Burnett, E.; Carlton, L.; Curran, A.T.; Lee, C.; Matsumoto, R.; Donnell, R.J.; Reich, I.; Willadsen, O. The introduced terrestrial slugs Ambigolimax nyctelius (Bourguignat, 1861) and Ambigolimax valentianus (Férussac, 1821) (Gastropoda: Limacidae) in California, with a discussion of taxonomy, systematics, and discovery by citizen science. J. Nat. Hist. 2019, 53, 1607–1632. [Google Scholar] [CrossRef]
- Mc Donnell, R.J.; Vlach, J.J.; Reich, I.; Colton, A.J. Boettgerilla pallens Simroth, 1912 (Boettgerillidae): A new invasive slug species in Oregon, U.S.A. Am. Malacol. Bull. 2020, 38, 63–65. [Google Scholar] [CrossRef]
- Horsák, M.; Naggs, F.; Backeljau, T. Paropeas achatinaceum (Pfeiffer, 1846) and other alien subulinine and opeatine land snails in European greenhouses (Gastropoda, Achatinidae). Malacologia 2020, 63, 123–130. [Google Scholar] [CrossRef]
- El-Danasoury, H.; Iglesias-Piñeiro, J.; Córdoba, M. The effect of climate manipulations on the herbivory of the pest slug Deroceras reticulatum (Müller, 1774) (Pulmonata: Agriolimacidae). Int. J. Biometeorol. Heidelb. 2016, 60, 1501–1507. [Google Scholar] [CrossRef]
- Hollingsworth, R.G.; Howe, K.; Jarvi, S.I. Control measures for slug and snail hosts of Angiostrongylus cantonensis, with special reference to the semi-slug Parmarion martensi. Hawaii J. Med. Public Health 2013, 72, 75–80. [Google Scholar]
- Cordoba, M.; Millar, J.G.; Mc Donnell, R. Development of a high-throughput laboratory bioassay for testing potential attractants for terrestrial snails and slugs. J. Econ. Entomol. 2018, 111, 637–644. [Google Scholar] [CrossRef]
- Roda, A.; Millar, J.G.; Jacobsen, C.; Veasey, R.; Fujimoto, L.; Hara, A.; McDonnell, R.J. A new synthetic lure for management of the invasive giant African snail, Lissachatina fulica. PLoS ONE 2019, 14, e0224270. [Google Scholar] [CrossRef] [PubMed]
- Cordoba, M.; Millar, J.G.; Foley, I.; Anderson, T.O.; Roda, A.L.; Adams, G.D.; Donnell, R.J. Fresh cucumber as an attractant for the invasive snail Xerolenta obvia. Am. Malacol. Bull. 2020, 37, 62–65. [Google Scholar] [CrossRef]
- Lucid, M.K.; Ehlers, S.; Robinson, L.; Cushman, S.A. Beer, brains, and brawn as tools to describe terrestrial gastropod species richness on a montane landscape. Ecosphere 2018, 9, e02535. [Google Scholar] [CrossRef] [Green Version]
- Piechowicz, B.; Grodzicki, P.; Ząbkiewicz, P.; Sobczyk, A.; Dąbrowska, A.; Piechowicz, I.; Pieniążek, M.; Balawejder, M.; Zareba, L. Components of the smell of beer as enticing factor for invasive slugs Arion lusitanicus Non-Mabille. Ecol. Chem. Eng. A 2018, 25, 133–151. [Google Scholar] [CrossRef]
- Pol, J.; Gries, R.; Gries, G. Rye Bread and synthetic bread odorants—Effective trap bait and lure for German cockroaches. Entomol. Exp. Appl. 2018, 166, 81–93. [Google Scholar] [CrossRef]
- Rowson, B.; Turner, J.; Anderson, R.; Symondson, B. Slugs of Britain and Ireland: Identification, Understanding and Control; Field Studies Council: Telford, UK, 2014; ISBN 978-1-908819-13-0. [Google Scholar]
- Jeong, K.J.; Lee, S.W.; Hong, J.K.; Shin, C.Y.; Yun, J.G. Effective control of slug damage through tobacco extract and caffeine solution in combination with alcohol. Hortic. Environ. Biotechnol. 2012, 53, 123–128. [Google Scholar] [CrossRef]
- Sanderson, G.; Sirgel, W. Helicidae as pests in Australian and South African grapevines. In Molluscs as Crop Pests; Barker, G.M., Ed.; CABI: Wallingford, UK, 2002; pp. 255–270. ISBN 978-0-85199-320-1. [Google Scholar]
- Port, G.; Ester, A. Gastropods as pests in vegetable and ornamental crops in Western Europe. In Molluscs as Crop Pests; Barker, G.M., Ed.; CABI: Wallingford, UK, 2002; pp. 337–351. ISBN 978-0-85199-320-1. [Google Scholar]
- Invasive Species Specialist Group ISSG. The Global Invasive Species Database. 2020. Available online: http://www.iucngisd.org/gisd/100_worst.php (accessed on 15 March 2020).
- Lo Re, V.; Gluckman, S.J. Eosinophilic meningitis. Am. J. Med. 2003, 114, 217–223. [Google Scholar] [CrossRef]
- Hollingsworth, R.G.; Kaneta, R.; Sullivan, J.J.; Bishop, H.S.; Qvarnstrom, Y.; da Silva, A.J.; Robinson, D.G. Distribution of Parmarion cf. martensi (Pulmonata: Helicarionidae), a new semi-slug pest on Hawai‘i Island, and its potential as a vector for human angiostrongyliasis. Pac. Sci. 2007, 61, 457–467. [Google Scholar] [CrossRef] [Green Version]
- Brodie, G.; Barker, G. Parmarion martensi Simroth, 1893. Family Ariophantidae; USP Introduced Land Snails of the Fiji Islands; University of the South Pacific: Suva, Fiji, 2012. [Google Scholar]
- Speiser, B. Food and feeding behaviour. In The Biology of Terrestrial Molluscs; Barker, G.M., Ed.; CABI: Wallingford, UK, 2001; pp. 259–288. ISBN 978-0-85199-318-8. [Google Scholar]
- IBM Corp. IBM SPSS Statistics for Windows, Version 24.0; IBM Corp.: Armonk, NY, USA, 2016. [Google Scholar]
- Mc Donnell, R.; Paine, T.; Gormally, M. Slugs: A Guide to the Native and Invasive Fauna of California; UCANR Press: Davis, CA, USA, 2009; ISBN 978-1-60107-564-2. [Google Scholar]
- Forsyth, R.G.; Oldham, M.J.; Snyder, E.; Schueler, F.W.; Layberry, R. Forty Years Later: Distribution of the introduced heath snail, Xerolenta obvia, in Ontario, Canada (Mollusca: Gastropoda: Hygromiidae). Check List 2015, 11, 1711. [Google Scholar] [CrossRef] [Green Version]
- Raut, S.K.; Barker, G.M. Achatina fulica Bowdich and other Achatinidae as pests in tropical agriculture. In Molluscs as Crop Pests; Barker, G.M., Ed.; CABI: Wallingford, UK, 2002; pp. 55–114. ISBN 978-0-85199-320-1. [Google Scholar]
- Cowie, R.; Hayes, K.; Kim, J.; Bustamente, K.; Yeung, N. Parmarion martensi Simroth, 1893 (Gastropoda: Ariophantidae), an intermediate host of Angiostrongylus cantonensis (rat lungworm), on Maui. Bish. Mus. Occas. Pap. 2018, 123, 7–10. [Google Scholar]
- USDA. APHIS Giant African Snail. Available online: https://www.aphis.usda.gov/aphis/resources/pests-diseases/hungry-pests/the-threat/giant-african-snail/giant-african-snail (accessed on 21 February 2021).
- Léo Neto, N.A.; Voeks, R.A.; Dias, T.L.; Alves, R.R. Mollusks of Candomblé: Symbolic and ritualistic importance. J. Ethnobiol. Ethnomedicine 2012, 8, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowie, R.H. Angiostrongylus cantonensis: Agent of a sometimes fatal globally emerging infectious disease (rat lungworm disease). ACS Chem. Neurosci. 2017, 8, 2102–2104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.R.; Hayes, K.A.; Yeung, N.W.; Cowie, R.H. Diverse gastropod hosts of Angiostrongylus cantonensis, the rat lungworm, globally and with a focus on the Hawaiian Islands. PLoS ONE 2014, 9, e94969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mc Donnell, R.; Yoo, J.; Patel, K.; Rios, L.; Hollingsworth, R.; Millar, J.; Paine, T. Can essential oils be used as novel drench treatments for the eggs and juveniles of the pest snail Cornu aspersum in potted plants? J. Pest Sci. 2016, 89, 549–555. [Google Scholar] [CrossRef]
- Dekle, G.W.; Fasulo, T.R. Featured Creatures. Brown Garden Snail—Cornu asperum (Müller); Publication number EENY-240; University of Florida Institute of Food and Agricultural Sciences: Gainesville, FL, USA, 2017. [Google Scholar]
- Bergey, E.A.; Figueroa, L.L.; Mather, C.M.; Martin, R.J.; Ray, E.J.; Kurien, J.T.; Westrop, D.R.; Suriyawong, P. Trading in snails: Plant nurseries as transport hubs for non-native species. Biol. Invasions 2014, 16, 1441–1451. [Google Scholar] [CrossRef]
- Frasse, P.; Lambert, S.; Richard-Molard, D.; Chiron, H. The influence of fermentation on volatile compounds in french bread dough. LWT Food Sci. Technol. 1993, 26, 126–132. [Google Scholar] [CrossRef]
- Gassenmeier, K.; Schieberle, P. Potent aromatic compounds in the crumb of wheat bread (french-type)—Influence of pre-ferments and studies on the formation of key odorants during dough processing. Z. Für Lebensm. Unters. Forsch. 1995, 201, 241–248. [Google Scholar] [CrossRef]
- Cranshaw, W. Attractiveness of Beer and Fermentation Products to the Gray Garden Slug, Agriolimax Reticulatum (Muller) (Mollusca: Limacidae); Technical Bulletin; Colorado State University: Fort Collins, CO, USA, 1997. [Google Scholar]
- CABI. Invasive Species Compendium. Datasheet, Achatina Fulica (Giant African Land Snail). Available online: https://www.cabi.org/isc/datasheet/2640#todistribution (accessed on 15 March 2021).






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Veasey, R.; Cordoba, M.; Colton, A.; Fujimoto, L.; Dodge, C.; Foley, I.; Adams, G.; Anderson, T.; Merenz, R.; Hara, A.; et al. Fermenting Bread Dough as a Cheap, Effective, Nontoxic, and Generic Attractant for Pest Snails and Slugs. Insects 2021, 12, 328. https://doi.org/10.3390/insects12040328
Veasey R, Cordoba M, Colton A, Fujimoto L, Dodge C, Foley I, Adams G, Anderson T, Merenz R, Hara A, et al. Fermenting Bread Dough as a Cheap, Effective, Nontoxic, and Generic Attractant for Pest Snails and Slugs. Insects. 2021; 12(4):328. https://doi.org/10.3390/insects12040328
Chicago/Turabian StyleVeasey, Robin, Maria Cordoba, Andrew Colton, Leonard Fujimoto, Christine Dodge, Ian Foley, Gary Adams, Taelor Anderson, Richard Merenz, Arnold Hara, and et al. 2021. "Fermenting Bread Dough as a Cheap, Effective, Nontoxic, and Generic Attractant for Pest Snails and Slugs" Insects 12, no. 4: 328. https://doi.org/10.3390/insects12040328
APA StyleVeasey, R., Cordoba, M., Colton, A., Fujimoto, L., Dodge, C., Foley, I., Adams, G., Anderson, T., Merenz, R., Hara, A., Roda, A., Millar, J., & Mc Donnell, R. (2021). Fermenting Bread Dough as a Cheap, Effective, Nontoxic, and Generic Attractant for Pest Snails and Slugs. Insects, 12(4), 328. https://doi.org/10.3390/insects12040328