
OBP14 (Odorant-Binding Protein) Sensing in Adelphocoris lineolatus Based on Peptide Nucleic Acid and Graphene Oxide


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Reagents
2.2. Insect Rearing and Collection
2.3. Characterization of GO
2.4. Preparation of the mRNA Biosensor
2.5. Target mRNA Detection In Vitro
2.5.1. The Feasibility of the mRNA Biosensor
2.5.2. The Specificity of the mRNA Biosensor
2.6. Target mRNA Detection in A. Lineolatus
2.6.1. PNA-mRNA Hybridization
2.6.2. Fluorescence Signals Observation
2.6.3. Tissue Clearing of A. Lineolatus
3. Results
3.1. Characterization of GO
3.2. Preparation of the mRNA Biosensor
3.3. Target mRNA Detection In Vitro
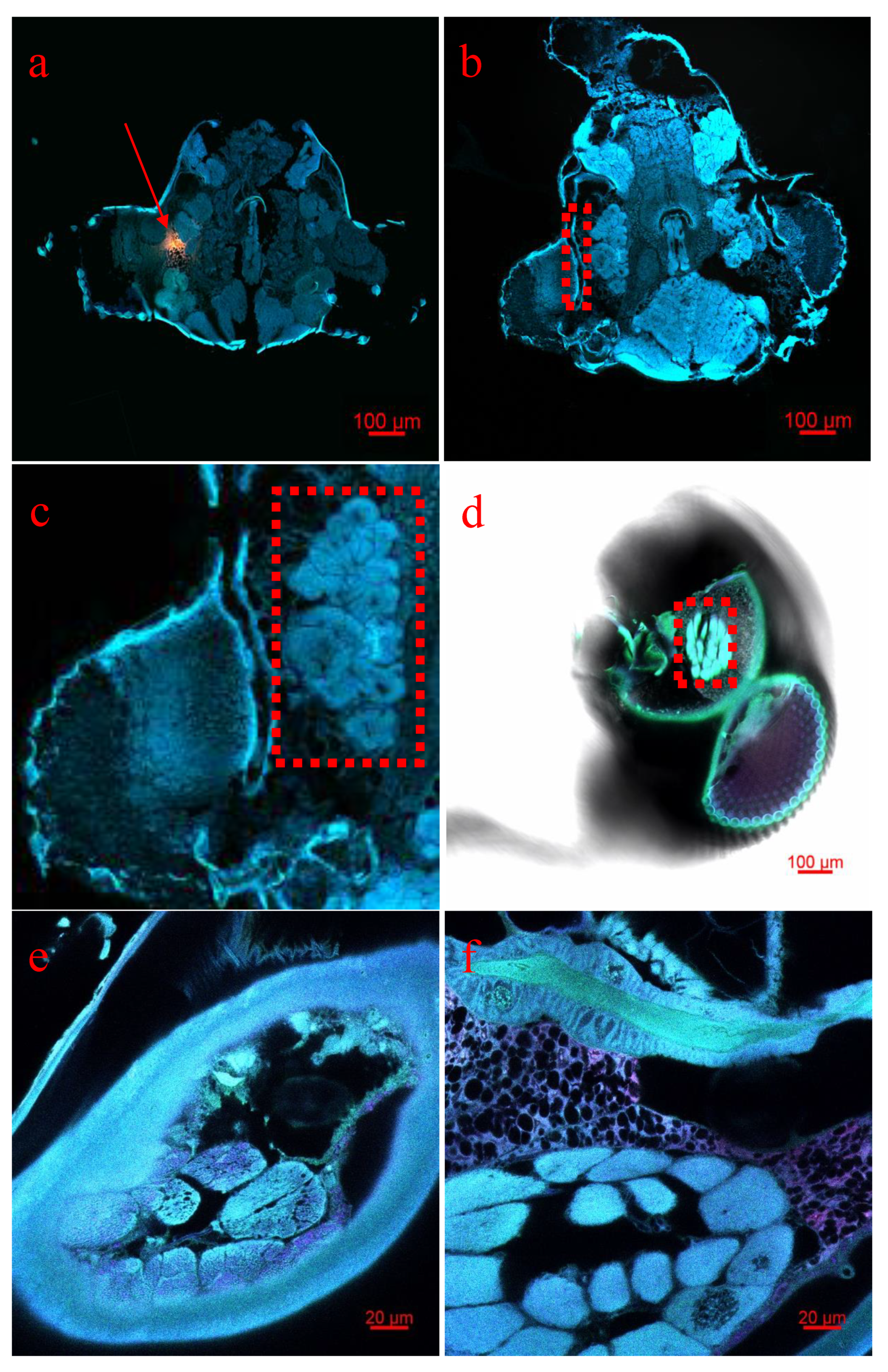
3.4. mRNA Detection in A. Lineolatus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hansson, B.S.; Stensmyr, M.C. Evolution of Insect Olfaction. Neuron 2011, 72, 698–711. [Google Scholar] [CrossRef] [Green Version]
- Sato, K.; Touhara, K. Insect olfaction: Receptors, signal transduction, and behavior. Results Probl. Cell Differ. 2008, 47, 121–138. [Google Scholar] [CrossRef]
- Field, L.M.; Pickett, J.A.; Wadhams, L.J. Molecular studies in insect olfaction. Insect Mol. Biol. 2000, 9, 545–551. [Google Scholar] [CrossRef]
- Fan, J.; Francis, F.; Liu, Y.; Chen, J.L.; Cheng, D.F. An overview of odorant-binding protein functions in insect peripheral olfactory reception. Genet. Mol. Res. 2011, 10, 3056–3069. [Google Scholar] [CrossRef]
- Brito, N.F.; Moreira, M.F.; Melo, A.C.A. A look inside odorant-binding proteins in insect chemoreception. J. Insect Physiol. 2016, 95, 51–65. [Google Scholar] [CrossRef]
- Lu, Y.; Wu, K.; Jiang, Y.; Xia, B.; Li, P.; Feng, H.; Wyckhuys, K.A.G.; Guo, Y. Mirid bug outbreaks in multiple crops correlated with wide-scale adoption of Bt cotton in China. Science 2010, 328, 1151–1154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, K.; Li, W.; Feng, H.; Guo, Y. Seasonal abundance of the mirids, Lygus lucorum and Adelphocoris spp. (Hemiptera: Miridae) on Bt cotton in northern China. Crop. Prot. 2002, 21, 997–1002. [Google Scholar] [CrossRef]
- Wu, K.M.; Guo, Y.Y. The evolution of cotton pest management practices in China. Annu. Rev. Entomol. 2005, 50, 31–52. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Sun, L.; Wang, Q.; Zhang, Q.; Gu, S.H.; Khashaveh, A.; Liu, Z.W.; Zhang, Y.J. Molecular characterization and expression analysis of putative odorant carrier proteins in Adelphocoris lineolatus. J. Asia Pacific Èntomol. 2018, 21, 958–970. [Google Scholar] [CrossRef]
- Campanini, E.B.; Congrains, C.; Torres, F.R.; De Brito, R.A. Odorant-binding proteins expression patterns in recently diverged species of Anastrepha fruit flies. Sci. Rep. 2017, 7, 2194. [Google Scholar] [CrossRef] [Green Version]
- He, M.; He, P. Molecular characterization, expression profiling, and binding properties of odorant binding protein genes in the whitebacked planthopper, Sogatella furcifera. Comp. Biochem. Physiol. B 2014, 174, 1–8. [Google Scholar] [CrossRef]
- Song, Y.Q.; Sun, H.Z.; Du, J. Identification and tissue distribution of chemosensory protein and odorant binding protein genes in Tropidothorax elegans Distant (Hemiptera: Lygaeidae). Sci. Rep. 2018, 8, 7803. [Google Scholar] [CrossRef] [PubMed]
- Bustin, S.A.; Benes, V.; Nolan, T.; Pfaffl, M.W. Quantitative real-time RT-PCR: A perspective. J. Mol. Endocrinol. 2005, 34, 597–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lü, J.; Yang, C.X.; Zhang, Y.J.; Pan, H.P. Selection of reference genes for the normalization of RT-qPCR data in gene expression studies in insects: A systematic review. Front. Physiol. 2018, 9, 1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.K.; Shanbhag, S.R.; Wang, Q.; Hasan, G.; Steinbrecht, R.A.; Pikielny, C.W. Expression patterns of two putative odorant-binding proteins in the olfactory organs of Drosophila melanogaster have different implications for their functions. Cell Tissue Res. 2000, 300, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Biessmann, H.; Nguyen, Q.K.; Le, D.; Walter, M.F. Microarray-based survey of a subset of putative olfactory genes in the mosquito Anopheles gambiae. Insect Mol. Biol. 2005, 14, 575–589. [Google Scholar] [CrossRef]
- Schultze, A.; Schymura, D.; Forstner, M.; Krieger, J. Expression pattern of a ‘Plus-C’ class odorant binding protein in the antenna of the malaria vector Anopheles gambiae. Insect Mol. Biol. 2012, 21, 187–195. [Google Scholar] [CrossRef]
- Compton, O.C.; Nguyen, S.T. Graphene oxide, highly reduced graphene oxide, and graphene: Versatile building blocks for carbon-based materials. Small 2010, 6, 711–723. [Google Scholar] [CrossRef]
- He, S.J.; Song, B.; Li, D.; Zhu, C.F.; Qi, W.P.; Wen, Y.Q.; Wang, L.H.; Song, S.P.; Fang, H.P.; Fan, C.H. A graphene nanoprobe for rapid, sensitive, and multicolor fluorescent DNA analysis. Adv. Funct. Mater. 2010, 20, 453–459. [Google Scholar] [CrossRef]
- Li, D.P.; Zhang, W.S.; Yu, X.Q.; Wang, Z.P.; Su, Z.Q.; Wei, G. When biomolecules meet graphene: From molecular level interactions to material design and applications. Nanoscale 2016, 8, 19491–19509. [Google Scholar] [CrossRef]
- Gu, S.H.; Wang, S.P.; Zhang, X.Y.; Wu, K.M.; Guo, Y.Y.; Zhou, J.J.; Zhang, Y.J. Identification and tissue distribution of odorant binding protein genes in the lucerne plant bug Adelphocoris lineolatus (Goeze). Insect Biochem. Mol. Biol. 2011, 41, 254–263. [Google Scholar] [CrossRef]
- Lu, Y.; Wu, K.; Guo, Y. Flight potential of Lygus lucorum (Meyer-Dür) (Heteroptera: Miridae). Environ. Entomol. 2007, 36, 1007–1013. [Google Scholar] [CrossRef] [PubMed]
- Jing, D.; Zhang, S.W.; Luo, W.J.; Gao, X.F.; Men, Y.; Ma, C.; Liu, X.H.; Yi, Y.T.; Bugde, A.; Zhou, B.O.; et al. Tissue clearing of both hard and soft tissue organs with the PEGASOS method. Cell Res. 2018, 28, 803–818. [Google Scholar] [CrossRef] [Green Version]
- Cui, L.; Chen, Z.; Zhu, Z.; Lin, X.; Chen, X.; Yang, C.J. Stabilization of ssRNA on graphene oxide surface: An effective way to design highly robust RNA probes. Anal. Chem. 2013, 85, 2269–2275. [Google Scholar] [CrossRef]
- Koppelhus, U.; Nielsen, P.E. Cellular delivery of peptide nucleic acid (PNA). Adv. Drug Deliv. Rev. 2003, 55, 267–280. [Google Scholar] [CrossRef]
- Zheng, P.; Wu, N.Q. Fluorescence and sensing applications of graphene oxide and graphene quantum dots: A review. Chem. Asian J. 2017, 12, 2343–2353. [Google Scholar] [CrossRef]
- Zinchuk, V.; Grossenbacher-Zinchuk, O. Recent advances in quantitative colocalization analysis: Focus on neuroscience. Prog. Histochem. Cytochem. 2009, 44, 125–172. [Google Scholar] [CrossRef]
- Xie, G.Y.; Ma, B.W.; Liu, X.L.; Chang, Y.J.; Chen, W.B.; Li, G.P.; Feng, H.Q.; Zhang, Y.J.; Berg, B.G.; Zhao, X.C. Brain organization of Apolygus lucorum: A hemipteran species with prominent antennal lobes. Front. Neuroanat. 2019, 13, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansson, B.S.; Anton, S. Function and morphology of the antennal lobe: New developments. Annu. Rev. Entomol. 2000, 45, 203–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelosi, P.; Zhou, J.J.; Ban, L.P.; Calvello, M. Soluble proteins in insect chemical communication. Cell. Mol. Life Sci. 2006, 63, 1658–1676. [Google Scholar] [CrossRef]
- Zhou, J.J. Odorant-binding proteins in insects. Vitam Horm. 2010, 83, 241–272. [Google Scholar] [CrossRef] [PubMed]
- Leal, W.S. Odorant reception in insects: Roles of receptors, binding proteins, and degrading enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef] [PubMed]
- Vogt, R.G.; Rogers, M.E.; Franco, M.D.; Sun, M. A comparative study of odorant binding protein genes: Differential expression of the PBP1-GOBP2 gene cluster in Manduca sexta (Lepidoptera) and the organization of OBP genes in Drosophila melanogaster (Diptera). J. Exp. Biol. 2002, 205, 719–744. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Zhang, J.; Liu, N.Y.; Zhang, Y.N.; Yang, K.; Dong, S.L. Distinct expression profiles and different functions of odorant binding proteins in Nilaparvata lugens Stål. PLoS ONE 2011, 6, e28921. [Google Scholar] [CrossRef] [Green Version]
- Steinbrecht, R.A. Odorant-binding proteins: Expression and function. Ann. N.Y. Acad. Sci. 1998, 855, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Li, Y.; Zhang, Z.; Guo, H.; Xiao, Q.; Wang, Q.; Zhang, Y. Expression patterns and ligand binding characterization of Plus-C odorant-binding protein 14 from Adelphocoris lineolatus (Goeze). Comp. Biochem. Physiol. B 2019, 227, 75–82. [Google Scholar] [CrossRef]
- Nielsen, P.E.; Egholm, M. An introduction to peptide nucleic acid. Curr. Issues Mol. Biol. 1999, 1, 89–104. [Google Scholar]
- Vilaivan, T. Fluorogenic PNA probes. Beilstein J. Org. Chem. 2018, 14, 253–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robertson, K.L.; Yu, L.P.; Armitage, B.A.; Lopez, A.J.; Peteanu, L.A. Fluorescent PNA probes as hybridization labels for biological RNA. Biochemistry 2006, 45, 6066–6074. [Google Scholar] [CrossRef]
- Egholm, M.; Buchardt, O.; Christensen, L.; Behrens, C.; Freier, S.M.; Driver, D.A.; Berg, R.H.; Kim, S.K.; Norden, B.; Nielsen, P.E. PNA hybridizes to complementary oligonucleotides obeying the Watson–Crick hydrogen-bonding rules. Nature 1993, 365, 566–568. [Google Scholar] [CrossRef]
- Ryoo, S.R.; Lee, J.; Yeo, J.; Na, H.K.; Kim, Y.K.; Jang, H.; Lee, J.H.; Han, S.W.; Lee, Y.; Kim, V.N.; et al. Quantitative and multiplexed microRNA sensing in living cells based on peptide nucleic acid and nano graphene oxide (PANGO). ACS Nano 2013, 7, 5882–5891. [Google Scholar] [CrossRef] [PubMed]
- Galindo, K.; Smith, D.P. A large family of divergent Drosophila odorant-binding proteins expressed in gustatory and olfactory sensilla. Genetics 2001, 159, 1059–1072. [Google Scholar] [CrossRef]
- Wang, S.N.; Shan, S.; Liu, J.T.; Li, R.J.; Lu, Z.Y.; Dhiloo, K.H.; Khashaveh, A.; Zhang, Y.J. Characterization of antennal chemosensilla and associated odorant binding as well as chemosensory proteins in the parasitoid wasp Microplitis mediator (Hymenoptera: Braconidae). Sci. Rep. 2018, 8, 7649. [Google Scholar] [CrossRef] [PubMed]
- Piesik, D.; Kalka, I.; Wenda-Piesik, A.; Bocianowski, J. Apion miniatum Germ. herbivory on the mossy sorrel, Rumex confertus Willd.: Induced plant volatiles and weevil orientation responses. Pol. J. Environ. Stud. 2014, 23, 2149–2156. [Google Scholar] [CrossRef]
- Piesik, D.; Wenda-Piesik, A.; Weaver, D.K.; Morrill, W.L. Influence of Fusarium crown rot disease on semiochemical production by wheat plants. J. Phytopathol. 2007, 155, 488–496. [Google Scholar] [CrossRef]
- Piesik, D.; Delaney, K.J.; Wenda-Piesik, A.; Sendel, S.; Tabaka, P.; Buszewski, B. Meligethes aeneus pollen-feeding suppresses, and oviposition induces, Brassica napus volatiles: Beetle attraction/repellence to lilac aldehydes and veratrole. Chemoecology 2013, 23, 241–250. [Google Scholar] [CrossRef]
- Nagegowda, D.A. Plant volatile terpenoid metabolism: Biosynthetic genes, transcriptional regulation and subcellular compartmentation. Febs. Lett. 2010, 584, 2965–2973. [Google Scholar] [CrossRef] [Green Version]
- Bellés, X.; Martín, D.; Piulachs, M.D. The mevalonate pathway and the synthesis of juvenile hormone in insects. Annu. Rev. Entomol. 2005, 50, 181–199. [Google Scholar] [CrossRef] [Green Version]
- Schulz, D.J.; Sullivan, J.P.; Robinson, G.E. Juvenile hormone and octopamine in the regulation of division of labor in honey bee colonies. Horm. Behav. 2002, 42, 222–231. [Google Scholar] [CrossRef] [Green Version]
- Ignell, R.; Couillaud, F.; Anton, S. Juvenile-hormone-mediated plasticity of aggregation behaviour and olfactory processing in adult desert locusts. J. Exp. Biol. 2001, 204, 249–259. [Google Scholar] [CrossRef]
- Guo, W.; Song, J.; Yang, P.; Chen, X.; Chen, D.; Ren, D.; Kang, L.; Wang, X. Juvenile hormone suppresses aggregation behavior through influencing antennal gene expression in locusts. PLoS Genet. 2020, 16, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Gadenne, C.; Anton, S. Central processing of sex pheromone stimuli is differentially regulated by juvenile hormone in a male moth. Insect Physiol. 2000, 46, 1195–1206. [Google Scholar] [CrossRef]
- Venthur, H.; Zhou, J.J. Odorant receptors and odorant-binding proteins as insect pest control targets: A comparative analysis. Front. Physiol. 2018, 9, 1163. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, W.; Zhang, T.; Gu, S.; Guo, Y.; Gao, X.; Zhang, Y. OBP14 (Odorant-Binding Protein) Sensing in Adelphocoris lineolatus Based on Peptide Nucleic Acid and Graphene Oxide. Insects 2021, 12, 422. https://doi.org/10.3390/insects12050422
Tian W, Zhang T, Gu S, Guo Y, Gao X, Zhang Y. OBP14 (Odorant-Binding Protein) Sensing in Adelphocoris lineolatus Based on Peptide Nucleic Acid and Graphene Oxide. Insects. 2021; 12(5):422. https://doi.org/10.3390/insects12050422
Chicago/Turabian StyleTian, Wenhua, Tao Zhang, Shaohua Gu, Yuyuan Guo, Xiwu Gao, and Yongjun Zhang. 2021. "OBP14 (Odorant-Binding Protein) Sensing in Adelphocoris lineolatus Based on Peptide Nucleic Acid and Graphene Oxide" Insects 12, no. 5: 422. https://doi.org/10.3390/insects12050422
APA StyleTian, W., Zhang, T., Gu, S., Guo, Y., Gao, X., & Zhang, Y. (2021). OBP14 (Odorant-Binding Protein) Sensing in Adelphocoris lineolatus Based on Peptide Nucleic Acid and Graphene Oxide. Insects, 12(5), 422. https://doi.org/10.3390/insects12050422