
Capsella bursa-pastoris Is a Key Overwintering Plant for Aphids in the Mediterranean Region


 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling Area
2.2. Sampling
2.3. Data Analysis
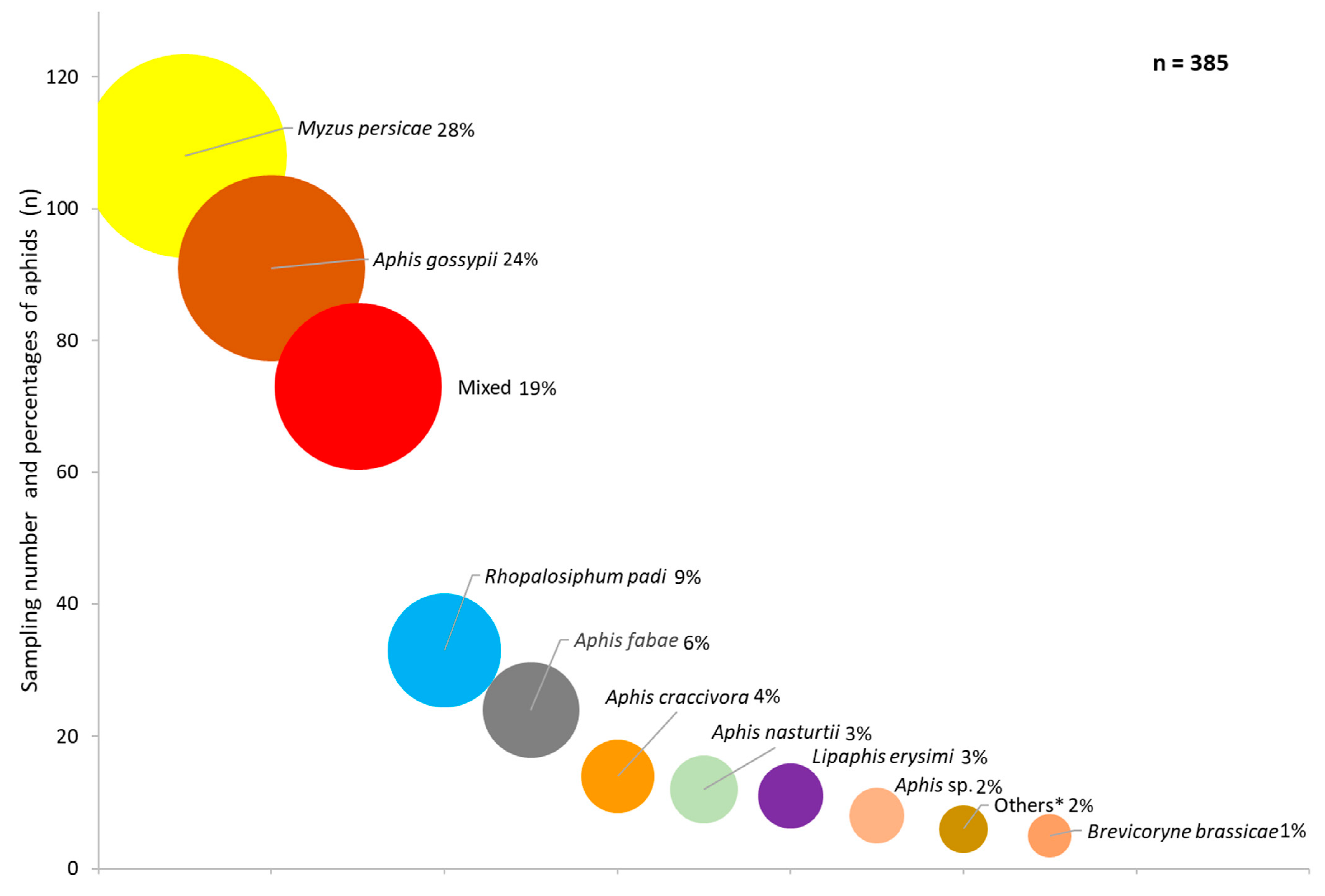
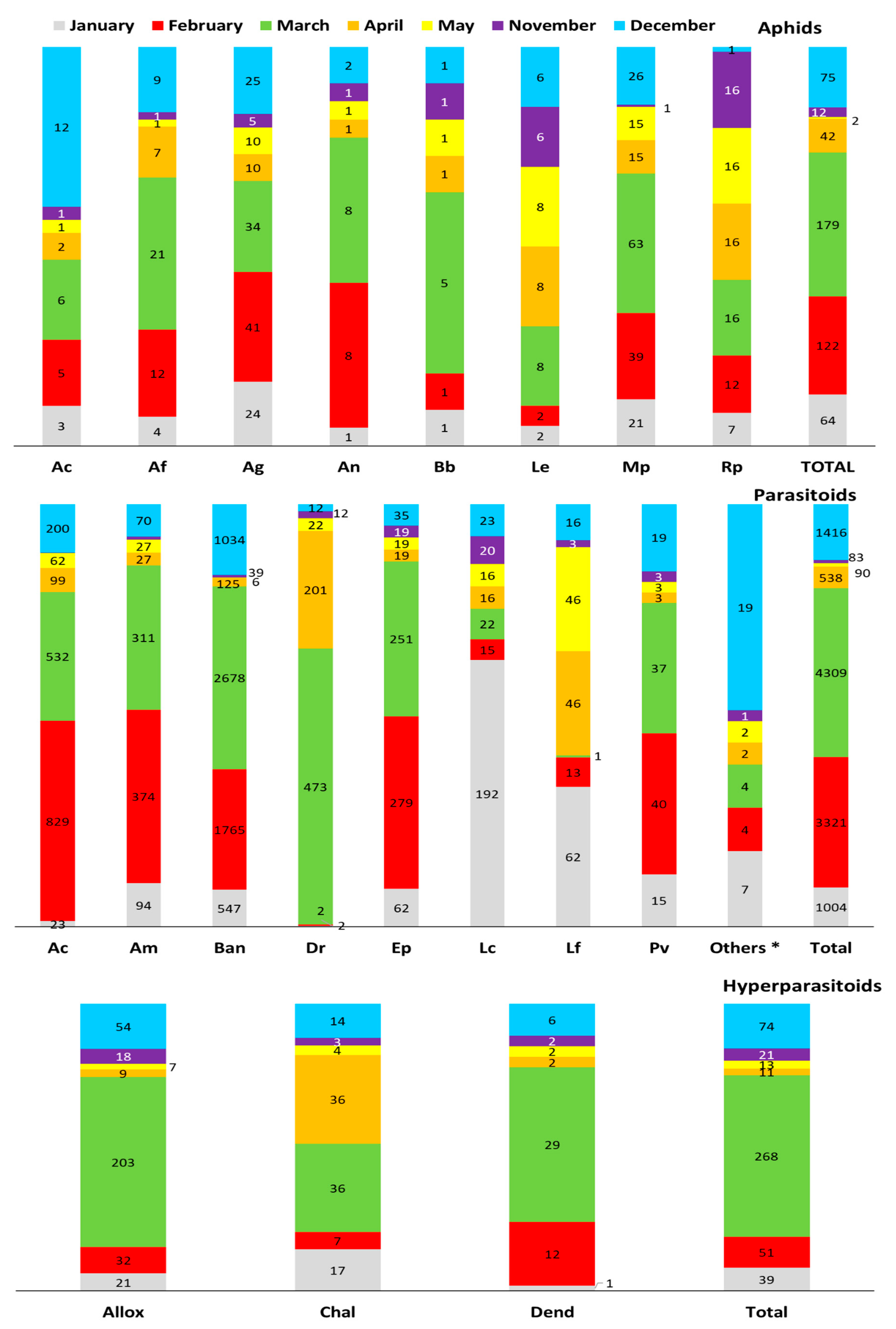
3. Results
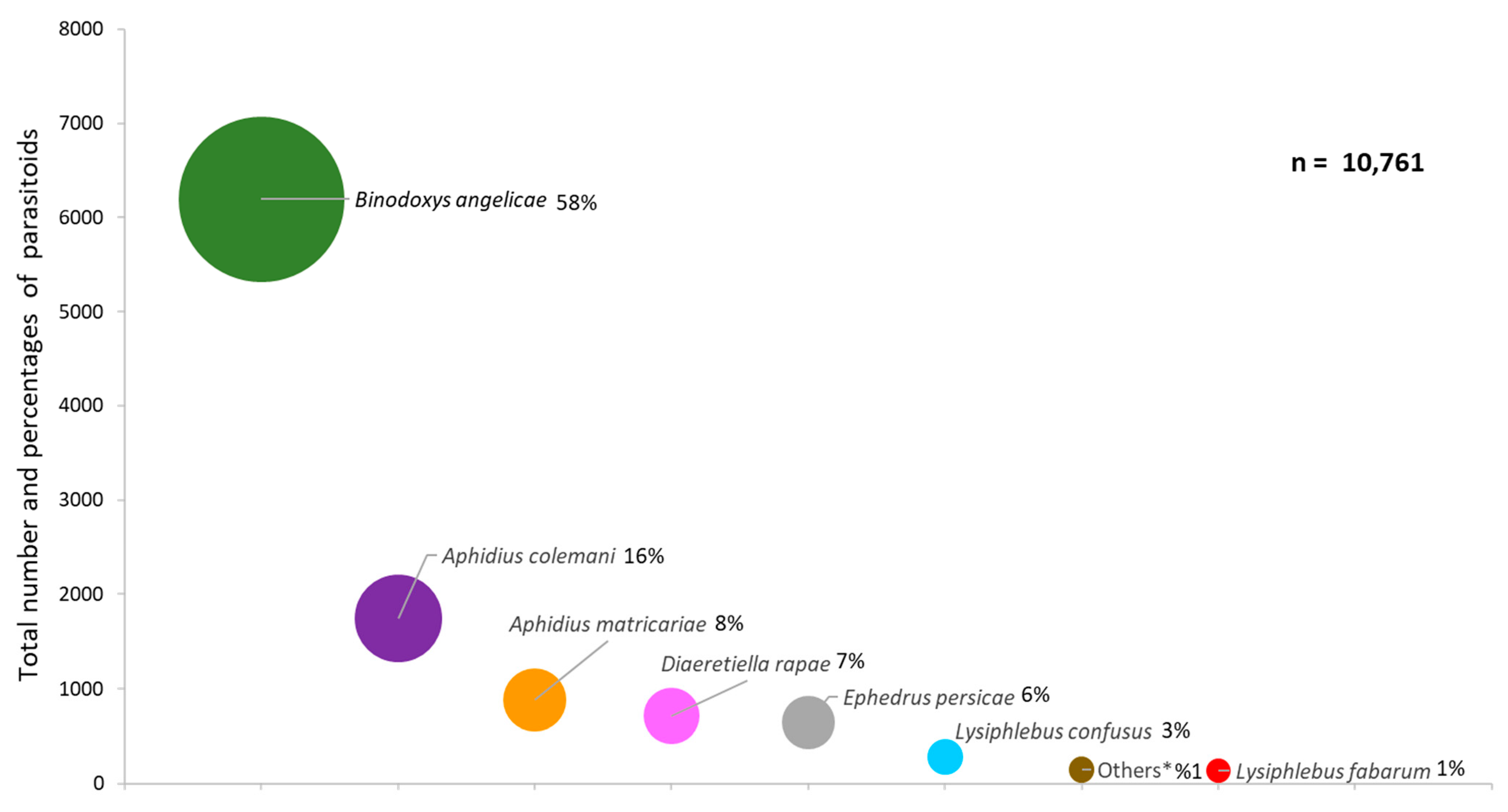
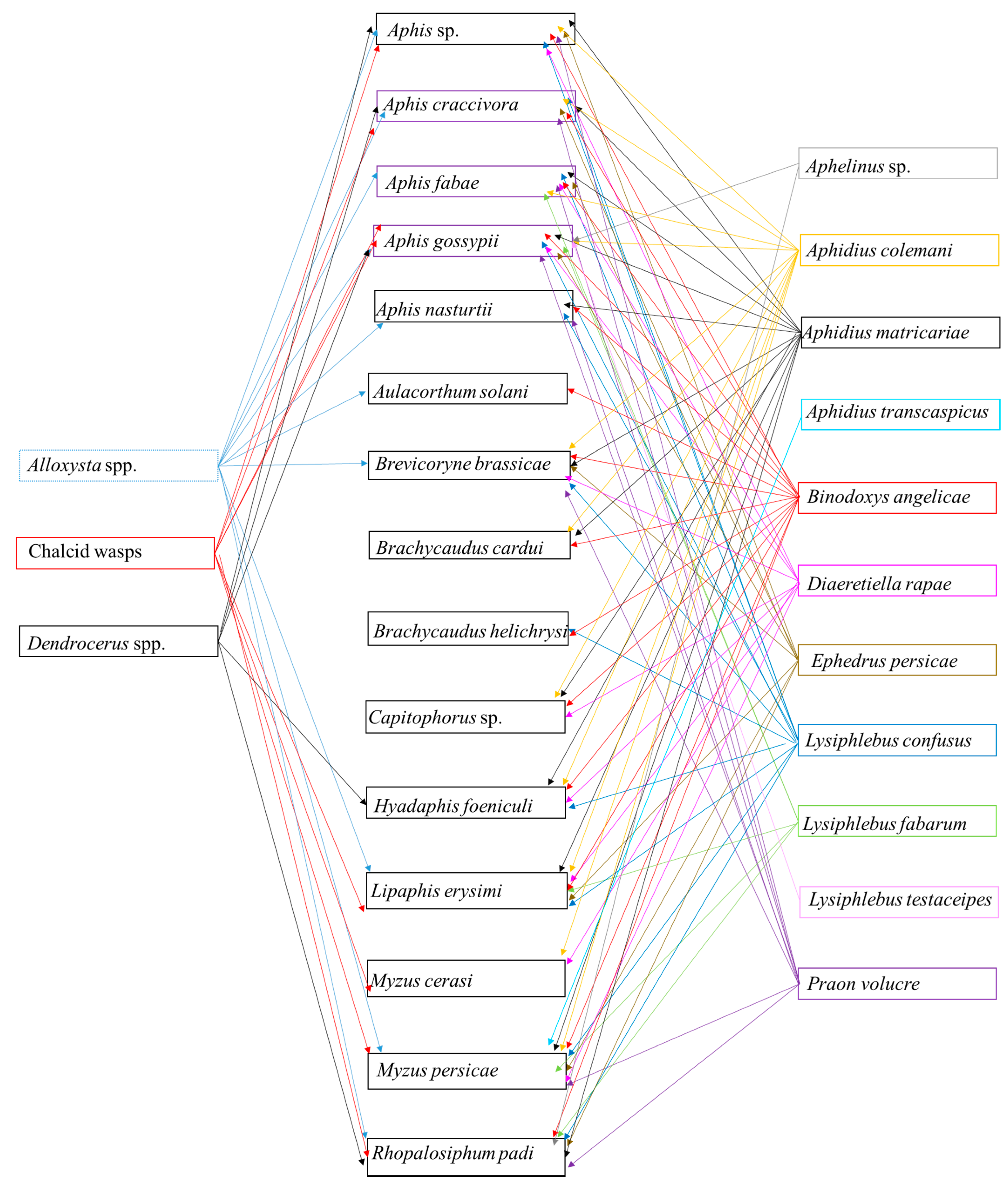
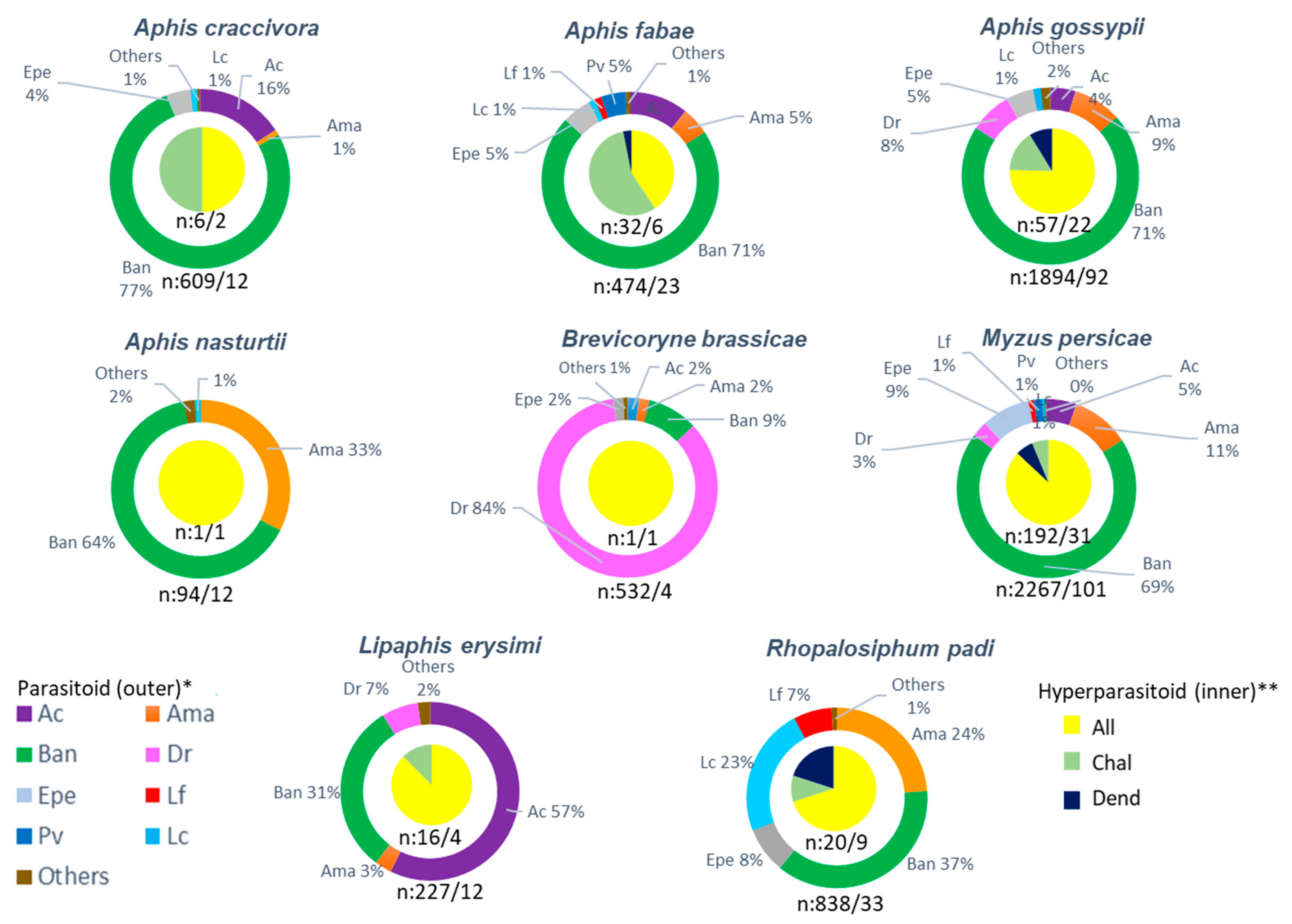
Relationship between Aphids and Parasitoids
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aksoy, A.; Dixon, J.M.; Hale, W.H. Capsella bursa-pastoris (L.) medikus (Thlaspi bursa-pastoris L., Bursa bursa-pastoris (L.) shull, Bursa pastoris (L.) weber). J. Ecol. 1998, 86, 171–186. [Google Scholar] [CrossRef]
- Satar, S. Biological Characteristic of Aphis spiraecola Patch (Homoptera: Aphididae) and Relationship with Parasitoid Lysiphlebia japonica (Ashmead) (Hymenoptera: Aphididae). Ph.D. Thesis, Cukurova University, Faculty of Agricultural, Department of Plant Protection, Adana, Turkey, 2003. [Google Scholar]
- Ceplitis, A.L.F.; Su, Y.; Lascoux, M. Bayesian inference of evolutionary history from chloroplast microsatellites in the cosmopolitan weed Capsella bursa-pastoris (Brassicaceae). Mol. Ecol. 2005, 14, 4221–4233. [Google Scholar] [CrossRef]
- Blackman, R.L.; Eastop, V.F. Aphids of the World’s Plants: An Online Identification and Information Guide. Host Lists and Keys. Available online: http://www.aphidsonworldsplants.info/C_HOSTS_Cab_Car.htm#Capsella (accessed on 10 October 2020).
- Fox, L.; Biever, K.D.; Toba, H.H.; Duffus, J.E.; Thomas, P.E. Overwintering and monitoring of potato leafroll virus in some wild crucifers. Am. Potato J. 1993, 70, 505–515. [Google Scholar] [CrossRef]
- Stevens, M.; Smith, H.G.; Hallsworth, P.B. The host-range of beet yellowing viruses among common arable weed species. Plant Pathol. 1994, 43, 579. [Google Scholar] [CrossRef]
- Satar, S.; Uygun, N. Life cycle of Aphis spiraecola Patch (Homoptera: Aphididae) in east Mediterranean region of Turkey and it’s development on some important host plants. IOBC/WPRS Bull. 2007, 38, 216–224. [Google Scholar]
- Satar, S.; Kersting, U.; Uygun, N. Transmission of Turkish citrus tristeza virus isolates by Aphis gossypii Glover (Homoptera: Aphididae) in the laboratory condition. IOBC/WPRS Bull. 2007, 38, 328–335. [Google Scholar]
- Satar, S.; Kersting, U.; Yokomi, R. Presence of two host races of Aphis gossypii Glover (Hemiptera: Aphididae) collected in Turkey. Ann. Appl. Boil. 2013, 162, 41–49. [Google Scholar] [CrossRef]
- Uygun, N.; Toros, S.; Ulusoy, R.; Satar, S.; Özdemir, I. Doğu Akdeniz Bölgesi Aphidoidea (Homoptera) Türleri ile Bunların Parazitoit ve Predatörlerinin Saptanması. Türkiye Bilimsel ve Teknik Araştırma Kurumu, Tarım ve Orman Araştırma Grubu Proje; No: TÜBİTAK-TOGTAG 1720; Ankara University: Ankara, Turkey, 2001; p. 214. [Google Scholar]
- Görür, G.; Senol, O.; Geizici, G.; Akyıldırım Begen, H.; Parmaksız, D. New aphid (Hemiptera: Aphidoidea) records from south eastern parts of Turkey. J. Insect Biodivers. Syst. 2017, 3, 257–264. [Google Scholar]
- Ng, J.C.K.; Perry, K.L. Transmission of plant viruses by aphid vectors. Mol. Plant Pathol. 2004, 5, 505–511. [Google Scholar] [CrossRef]
- Starý, P. Aphidiidae. In Aphids, Their Biology, Natural Enemies and Control; Minks, A.K., Harrewijn, P., Eds.; Elsevier: Amsterdam, The Netherland, 1988; pp. 171–184. [Google Scholar]
- Hågvar, E.B.; Hofsvang, T. Aphid parasitoids (Hymenoptera, Aphidiidae): Biology, host selection and use in biological control. Biocontrol. News Inf. 1991, 12, 13–42. [Google Scholar]
- Starý, P. Field establishment of Aphidius colemani Vier. (Hym., Braconidae, Aphidiinae) in the Czech Republic. J. Appl. Entomol. 2002, 126, 405–408. [Google Scholar] [CrossRef]
- Starý, P. Biology of Aphid Parasites (Hymenoptera: Aphidiidae) with Respect to Integrated Control; Dr. W. Junk N.V.: The Hague, The Netherlands, 1970. [Google Scholar]
- Kılınçer, N. Der parasitenkomplex der mehligen kohlblattlaus in Ankara umfasst zwei parasiten und vier hyperparasitenarten. Plant Prot. Bull. 1982, 22, 1–12. [Google Scholar]
- Zeren, O.; Düzgüneş, Z. A study on identification, host plants, effect on host plant and natural enemies of aphids, causing damage on vegetables in Çukurova region. Ministry turkish ministry of agriculture, Ankara, Turkey. Res. Ser. 1989, 59, 205. [Google Scholar]
- Ölmez, S.; Ulusoy, M.R. A survey of aphid parasitoids (Hymenoptera: Braconidae: Aphidiinae) in Diyarbakır, Turkey. Phytoparasitica 2003, 31, 524–528. [Google Scholar] [CrossRef]
- Aslan, M.M.; Uygun, N.; Starý, P. A survey of aphid parasitoids in Kahramanmaraş, Turkey (Hymenoptera: Braconidae: Aphidiinae and Hymenoptera: Aphelinidae). Phytoparasitica 2004, 32, 255–263. [Google Scholar] [CrossRef]
- Uysal, M.; Starý, P.; Sahbaz, A.; Özsemerci, F. A review of aphid parasitoids (Hymenoptera: Braconidae: Aphidiinae) of Turkey. Egypt. J. Biol. Pest Control 2004, 14, 355–370. [Google Scholar]
- Güz, N.; Kılınçer, N. Aphid parasitoids (Hymenoptera: Braconidae: Aphidiinae) on weeds from Ankara, Turkey. Phytoparasitica 2005, 33, 359–366. [Google Scholar] [CrossRef]
- Tomanović, Ž.; Beyarslan, A.; Erdoğan, Ö.Ç. New records of aphid parasitoids (Hymenoptera: Braconidae: Aphidiinae) from Turkey. Period. Biol. 2008, 110, 335–338. [Google Scholar]
- Satar, S.; Karacaoğlu, M.; Satar, G.; Uygun, N. Studies on introduction of Lysiphlebus testaceipes (Cresson) (Hymenoptera: Braconidae), an important parasitoid of Aphis gossypii Glover (Hemiptera: Aphididae) in citrus orchards of east Mediterranean region of Turkey. Türk. Biyo. Müc. Derg. 2011, 2, 55–62. [Google Scholar]
- Satar, S.; Satar, G.; Karacaoğlu, M.; Uygun, N.; Kavallieratos, N.G.; Starý, P.; Athanassiou, C.G. Parasitoids and hyperparasitoids (Hymenoptera) on aphids (Hemiptera) infesting citrus in east Mediterranean region of Turkey. J. Insect Sci. 2014, 14, 178. [Google Scholar] [CrossRef] [Green Version]
- Rakhshani, E.; Barahoei, H.; Ahmad, Z.; Starý, P.; Ghafouri Moghaddam, M.; Mehrparvar, M.; Kavallieratos, N.G.; Čkrkić, J.; Tomanović, Ž. Review of Aphidiinae parasitoids (Hymenoptera: Braconidae) of the Middle East and North Africa: Key to species and host associations. Eur. J. Taxon. 2019, 552. [Google Scholar] [CrossRef]
- Starý, P.; Havelka, J. Fauna and associations of aphid parasitoids in an up-dated farmland area (Czech Republic). Bull. Insectol. 2008, 61, 251–276. [Google Scholar]
- Stenberg, J.A. A conceptual framework for integrated pest management. Trends Plant Sci. 2017, 22, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat management to conserve natural enemies of arthropod pests in agriculture. Annu. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef] [PubMed]
- Amoabeng, B.W.; Stevenson, P.C.; Mochiah, B.M.; Asare, K.P.; Gurr, G.M. Scope for non-crop plants to promote conservation biological control of crop pests and serve as sources of botanical insecticides. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Winiarska, W. Predatory and parasitic insects in colonies of Aphis/Aphis/ fabae cirsiiacanthoidis Scop. on the thistle, Cirsium arvense (L.) Scop. In Materialy Ogolnopolskie Konferencii Naukowej, Nauka y Practyce Ogrodniczej z Okazji XXV-Lecia, Wydaw. Ogrod. Akademii Rolniczej v Lublinie; Filipowicz, A.J., Ed.; Adademia Rolnicza: Lublin, Poland, 1995; pp. 403–406. [Google Scholar]
- Starý, P. Philadelphus coronarius L. as reservoir of aphids and parasitoids. J. Appl. Entomol. 1991, 112, 1–10. [Google Scholar] [CrossRef]
- Barczak, T. Reservoirs and-alternative hosts of aphid parasitoids. Discussion on methods used. Wiad. Entomol. 1996, 15, 181–187. [Google Scholar]
- Bennewicz, J.; Kaczorowski, G.; Barczak, T. Aphids in midfield thickets in lower vistula valley landscape park. J. Plant Prot. Res. 2001, 41, 348–362. [Google Scholar]
- Kavallieratos, N.G.; Stathas, G.J.; Athanassiou, C.G.; Papadoulis, G.T. Dittrichia viscosa and Rubus ulmifolius as reservoirs of aphid parasitoids (Hymenoptera: Braconidae: Aphidiinae) and the role of certain coccinellid species. Phytoparasitica 2002, 30, 231–242. [Google Scholar] [CrossRef]
- Tomanović, Ž.; Kavallieratos, N.G.; Starý, P.; Petrović, O.; Tomanović, S.; Jovanović, S. Aphids and parasitoids on willows and poplars in southeastern Europe (Homoptera: Aphidoidea, Hymenoptera: Braconidae: Aphidiinae). J. Plant Dis. Prot. 2006, 113, 174–180. [Google Scholar] [CrossRef]
- Kavallieratos, N.G.; Tomanović, Ž.; Starý, P.; Emmanouel, N.E. Vitex agnus-castus and Euphorbia characias ssp. wulfenii as reservoirs of aphid parasitoids (Hymenoptera: Braconidae: Aphidiinae). Fla. Entomol. 2008, 91, 179–191. [Google Scholar] [CrossRef]
- Tomanović, Ž.; Kavallieratos, N.G.; Starý, P.; Stanisavljević, L.Ž.; Ćetković, A.; Stamenković, S.; Jovanović, S.; Athanassiou, C.G. Regional tritrophic relationship patterns of five aphid parasitoid species (Hymenoptera: Braconidae: Aphidiinae) in agroecosystem-dominated landscapes of southeastern Europe. J. Econ. Entomol. 2009, 102, 836–854. [Google Scholar] [CrossRef] [PubMed]
- Kavallieratos, N.G.; Tomanović, Ž.; Petrović, A.; Kocić, K.; Janković, M.; Starý, P. Parasitoids (Hymenoptera: Braconidae: Aphidiinae) of aphids feeding on ornamental trees in southeastern Europe: Key for identification and tritrophic associations. Ann. Entomol. Soc. Am. 2016, 109, 473–487. [Google Scholar] [CrossRef]
- Pons, X.; Comas, J.; Albajes, R. Overwintering of cereal aphids (Homoptera: Aphididae) on durum wheat in a Mediterranean climate. Environ. Entomol. 1993, 22, 381–387. [Google Scholar] [CrossRef]
- Eastop, V.F.; van Emden, H.F. The insect material. In Aphid Technology; van Emden, H.F., Ed.; Academic Press: London, UK, 1972; pp. 1–45. [Google Scholar]
- Krantz, G.W. A Manual of Acarology, 2nd ed.; Oregon State University Book Stores: Corvallis, OR, USA, 1978. [Google Scholar]
- Rohlf, F.J.; Sokal, R.R. Statistical Tables; Freeman & Company: New York, NY, USA, 1994. [Google Scholar]
- Norris, R.F.; Kogan, M. Interactions between weeds, arthropod pests, and their natural enemies in managed ecosystems. Weed Sci. 2000, 48, 94–158. [Google Scholar] [CrossRef]
- Powell, G.; Tosh, C.R.; Hardie, J. Host plant selection by aphids: Behavioral, evolutionary, and applied perspectives. Annu. Rev. Entomol. 2006, 51, 309–330. [Google Scholar] [CrossRef]
- Tamaki, G.; Olsen, D. Evaluation of orchard weed hosts of green peach aphid and the production of winged migrants. Environ. Entomol. 1979, 8, 314–317. [Google Scholar] [CrossRef]
- Cole, R.A. The relative importance of glucosinolates and amino acids to the development of two aphid pests Brevicoryne brassicae and Myzus persicae on wild and cultivated brassica species. Entomol. Exp. Appl. 1997, 85, 121–133. [Google Scholar] [CrossRef]
- Van Emden, H.F.; Storeck, A.P.; Douloumpaka, S.; Eleftherianos, I.; Poppy, G.M.; Powell, W. Plant chemistry and aphid parasitoids (Hymenoptera: Braconidae): Imprinting and memory. Eur. J. Entomol. 2008, 105, 477–483. [Google Scholar] [CrossRef] [Green Version]
- Satar, S.; Karacaoğlu, M.; Satar, G.; Uygun, N. Citrus aphids (Hemiptera: Aphididae): Incidence, population fluctuations, host plant and age preferences. Plant Protect. Bull. 2020, 60, 111–119. [Google Scholar]
- Satar, S.; Kersting, U.; Uygun, N. Effect of temperature on population parameters of Aphis gossypii Glover and Myzus persicae (Sulzer) (Homoptera: Aphididae) on pepper. J. Plant Dis. Prot. 2008, 115, 69–74. [Google Scholar] [CrossRef]
- Manachini, B.; Casati, P.; Cinanni, L.; Bianco, P. Role of Myzus persicae (Hemiptera: Aphididae) and its secondary hosts in plum pox virus propagation. J. Econ. Entomol. 2007, 100, 1047–1052. [Google Scholar] [CrossRef]
- Plećaš, M.; Gagić, V.; Janković, M.; Petrović Obradović, O.; Kavallieratos, N.G.; Tomanović, Ž.; Thies, C.; Tscharntke, T.; Ćetković, A. Landscape composition and configuration influence cereal aphid-parasitoid-hyperparasitoid interactions and biological control differentially across years. Agric. Ecosyst. Environ. 2014, 183, 1–10. [Google Scholar] [CrossRef]
- Bayhan, S.Ö.; Ulusoy, M.R. Adana ilinde lahana unlu yaprakbiti (Brevicoryne brassicae L. (Hemiptera: Aphididae) ‘nin bazı curiciferae familyasına bağlı bitkilerdeki populasyon gelişimi. Harran Üni. Zir. Fak. Der. 2010, 14, 37–46. [Google Scholar]
- Powell, W.; Wright, A.F. The influence of host food plants on host recognition by four aphidiine parasitoids (Hymenoptera: Braconidae). Bull. Entomol. Res. 1992, 81, 449–453. [Google Scholar] [CrossRef]
- Decker, U.M. Evidence for Semiochemicals Affecting the Reproductive Behaviour of the Aphid Parasitoids Aphidius rhopalosiphi De Stefani-Perez and Praon volucre Haliday (Hymenoptera: Aphidiidae)—A Contribution towards Integrated Pest Management in Cereals. Ph.D. Thesis, University of Hohenheim, Stuttgart, Germany, 1988. [Google Scholar]
- Vinson, S.B. Host selection by insect parasitoids. Annu. Rev. Entomol. 1976, 21, 109–133. [Google Scholar] [CrossRef]
- de Vos, M.; Jander, G. Volatile communication in plant-aphid interactions. Curr. Opin. Plant Boil. 2010, 13, 366–371. [Google Scholar] [CrossRef]
- Kos, M.; Houshyani, B.; Wietsma, R.; Kabouw, P.; Vet, L.E.; van Loon, J.J.; Dicke, M. Effects of glucosinolates on a generalist and specialist leaf-chewing herbivore and an associated parasitoid. Phytochemistry 2012, 77, 162–170. [Google Scholar] [CrossRef] [Green Version]
- Kos, M.; Houshyani, B.; Achhami, B.B.; Wietsma, R.; Gols, R.; Weldegergis, B.T.; Kabouw, P.; Bouwmeester, H.J.; Vet, L.E.M.; Dicke, M.; et al. Herbivore-mediated effects of glucosinolates on different natural enemies of a specialist aphid. J. Chem. Ecol. 2012, 38, 100–115. [Google Scholar] [CrossRef] [Green Version]
- Murali Baskaran, R.K.; Sharma, K.C.; Kaushal, P.; Kumar, J.; Parthiban, P.; Senthil Nathan, S.; Mankin, R.W. Role of kairomone in biological control of crop pests—A review. Physiol. Mol. Plant Path. 2018, 101, 3–15. [Google Scholar] [CrossRef]
- Singh, R.; Tripathi, R.N. Records of aphid hyperparasitoids in India. Bioved 1991, 1, 141–150. [Google Scholar]
- Brodeur, J.; McNeil, J.N. Host behavior modification by the endoparasitoid Aphidius nigripes: A strategy to reduce hyperparasitism. Ecol. Entomol. 1992, 17, 97–104. [Google Scholar] [CrossRef]
- Müller, C.B.; Völkl, V.; Godfray, H.C.J. Are behavioural changes in parasitised aphids a protection against hyperparasitism? Eur. J. Entomol. 1997, 94, 221–234. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Main Aphid Species | Mixed Aphid Species | Parasitoid Species | ||||
|---|---|---|---|---|---|---|
| A. colemani | A. matricariae | B. angelicae | Others | Total | ||
| A.fabae | A. pisum, Aphis sp., | 953 | 38 | 564 | 133 | 1688 |
| A. craccivora, A. fabae solanella, A. gossypii, Cavariella sp., Coloradoa sp., H. foeniculi, | ||||||
| H. lactucae, L. erysimi, | ||||||
| M. ornatus, M. persicae, | ||||||
| M. rosarum | ||||||
| A. gossypii | A. craccivora, B. brassicae, | 41 | 106 | 397 | 165 | 709 |
| M. persicae, Phopalosiphum sp., R. maidis | ||||||
| B. brassicae | Capitophorus sp., M. persicae | 12 | 0 | 8 | 2 | 22 |
| Hyadaphis sp. | L. erysimi | 2 | 2 | 23 | 0 | 27 |
| M. persicae | Aphis sp., A. craccivora, | 3 | 19 | 43 | 14 | 79 |
| R. maidis, R. padi | ||||||
| R. padi | A. craccivora, A. fabae solanella | 1 | 0 | 3 | 3 | 7 |
| Parasitoid Numbers in Aphid Species | Df | χ2 | Aphid-Parasitoid Associations | Df | χ2 |
|---|---|---|---|---|---|
| A. craccivora vs. A. fabae | 5 | 27.9 | A. colemani vs. A. matricariae | 7 | 434.5 |
| A. craccivora vs. A. gossypii | 5 | 169.4 | A. colemani vs. B. angelicae | 7 | 498.1 |
| A. craccivora vs. A. nasturtii | 4 | 180.4 | A. colemani vs. D. rapae | 5 | 640.9 |
| A. craccivora vs. B. brassicae | 5 | 923.5 | A. colemani vs. E. persicae | 6 | 238.4 |
| A. craccivora vs. L. erysimi | 5 | 188.6 | A. colemani vs. L. confusus | 6 | 503.1 |
| A. craccivora vs. M. persicae | 5 | 205.4 | |||
| A. craccivora vs. R. padi | 4 | 491.1 | |||
| A. fabae vs. A. gossypii | 5 | 59.7 | A. matricariae vs. B. angelicae | 7 | 384.6 |
| A. fabae vs. A. nasturtii | 5 | 73. 7 | A. matricariae vs. D. rapae | 7 | 793.5 |
| A. fabae vs. B. brassicae | 5 | 755.9 | A. matricariae vs. E. persicae | 7 | 68.8 |
| A. fabae vs. L. erysimi | 5 | 184.5 | A. matricariae vs. L. confusus | 7 | 196.9 |
| A. fabae vs. M. persicae | 5 | 88.5 | |||
| A. fabae vs. R. padi | 5 | 317.8 | |||
| A. gossypii vs. A. nasturtii | 5 | 69.7 | B. angelicae vs. D. rapae | 7 | 2345.3 |
| A. gossypii vs. B. brassicae | 5 | 1399.6 | B. angelicae vs. E. persicae | 7 | 185.0 |
| A. gossypii vs. L. erysimi | 5 | 504.9 | B. angelicae vs. L. confusus | 7 | 910.5 |
| A. gossypii vs. M. persicae | 5 | 181.9 | |||
| A. gossypii vs. R. padi | 5 | 675.8 | |||
| A. nasturtii vs. B. brassicae | 5 | 413.2 | D. rapae vs. E. persicae | 6 | 636.2 |
| A. nasturtii vs. L. erysimi | 5 | 118.9 | D. rapae vs. L. confusus | 6 | 715.7 |
| A. nasturtii vs. M. persicae | 5 | 40.7 | |||
| A. nasturtii vs. R. padi | 3 | 43.6 | |||
| B. brassicae vs. L. erysimi | 5 | 448.8 | E. persicae vs. L. confusus | 6 | 241.8 |
| B. brassicae vs. M. persicae | 5 | 1373.7 | |||
| B. brassicae vs. R. padi | 5 | 1087.9 | |||
| L. erysimi vs. M. persicae | 5 | 307.0 | |||
| L. erysimi vs. R. padi | 5 | 585.8 | |||
| M. persicae vs. R. padi | 5 | 458.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Satar, S.; Kavallieratos, N.G.; Tüfekli, M.; Satar, G.; Athanassiou, C.G.; Papanikolaou, N.E.; Karacaoğlu, M.; Özdemir, I.; Starý, P. Capsella bursa-pastoris Is a Key Overwintering Plant for Aphids in the Mediterranean Region. Insects 2021, 12, 744. https://doi.org/10.3390/insects12080744
Satar S, Kavallieratos NG, Tüfekli M, Satar G, Athanassiou CG, Papanikolaou NE, Karacaoğlu M, Özdemir I, Starý P. Capsella bursa-pastoris Is a Key Overwintering Plant for Aphids in the Mediterranean Region. Insects. 2021; 12(8):744. https://doi.org/10.3390/insects12080744
Chicago/Turabian StyleSatar, Serdar, Nickolas G. Kavallieratos, Mustafa Tüfekli, Gül Satar, Christos G. Athanassiou, Nikos E. Papanikolaou, Mehmet Karacaoğlu, Işıl Özdemir, and Petr Starý. 2021. "Capsella bursa-pastoris Is a Key Overwintering Plant for Aphids in the Mediterranean Region" Insects 12, no. 8: 744. https://doi.org/10.3390/insects12080744
APA StyleSatar, S., Kavallieratos, N. G., Tüfekli, M., Satar, G., Athanassiou, C. G., Papanikolaou, N. E., Karacaoğlu, M., Özdemir, I., & Starý, P. (2021). Capsella bursa-pastoris Is a Key Overwintering Plant for Aphids in the Mediterranean Region. Insects, 12(8), 744. https://doi.org/10.3390/insects12080744