
Mitogenome Analysis of Four Lamiinae Species (Coleoptera: Cerambycidae) and Gene Expression Responses by Monochamus alternatus When Infected with the Parasitic Nematode, Bursaphelenchus mucronatus

 ,
,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling Collection and Microscopic Examination
2.2. DNA Extraction, Sequencing, and Molecular Identification
2.3. Sequence Annotation and Analyses
2.4. Comparison of Protein Coding Genes and Phylogenetic Analyses
2.5. RNA Isolation and cDNA Synthesis
2.6. RT-qPCR
3. Results
3.1. Nematode Identification
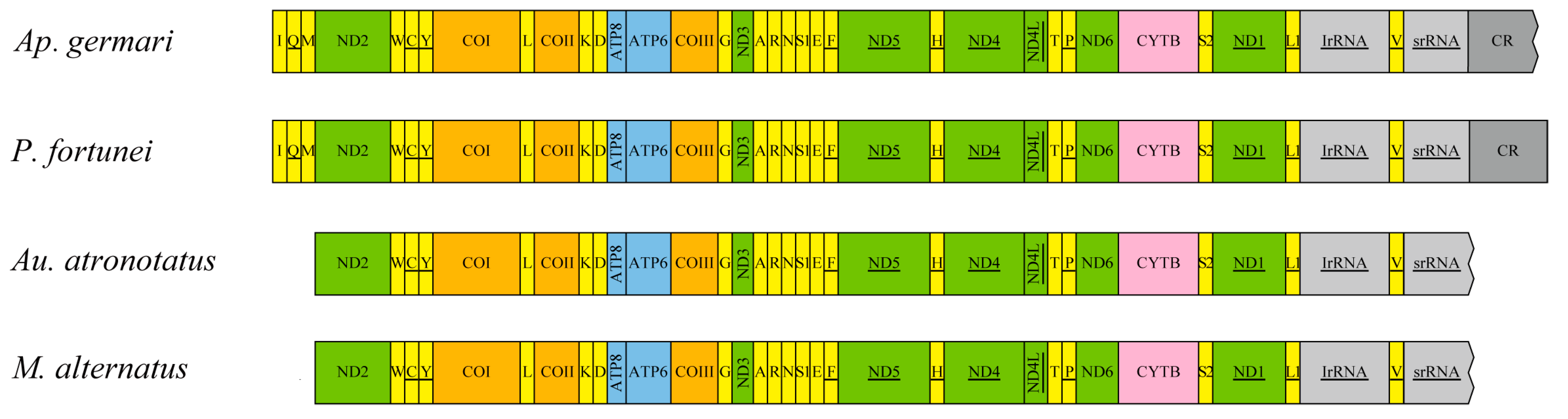
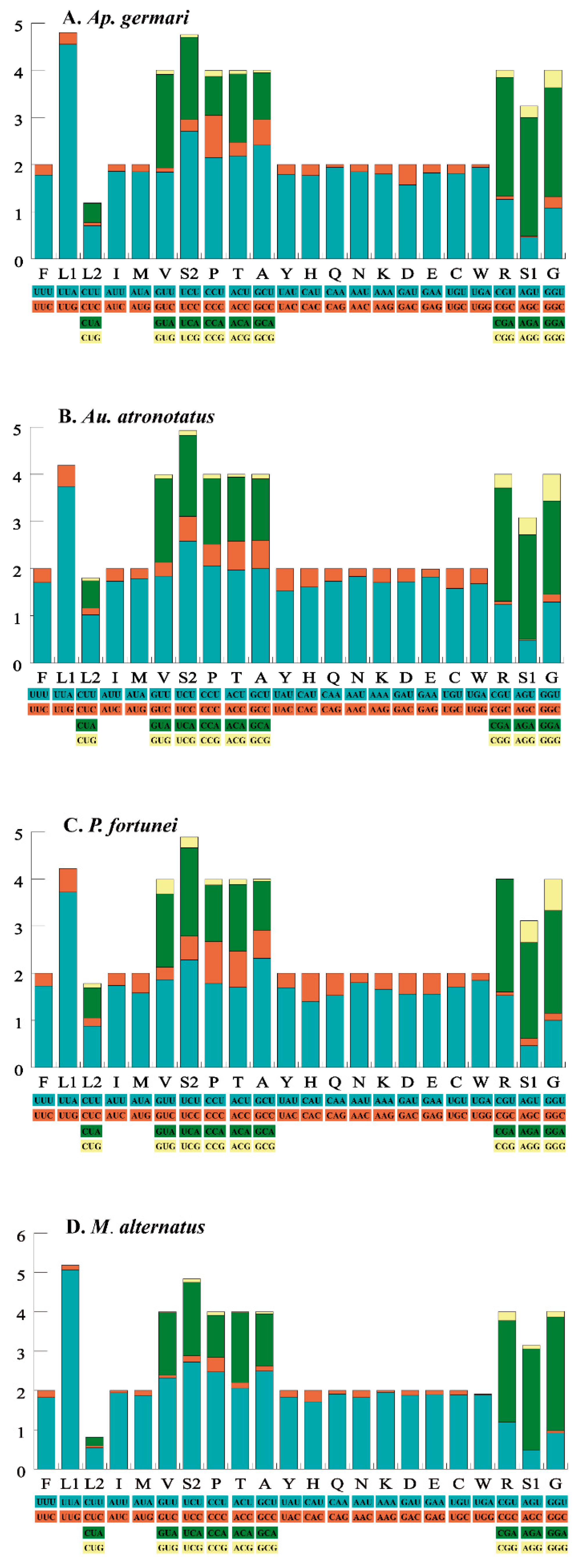
3.2. Components of the Mitochondrial Genomes
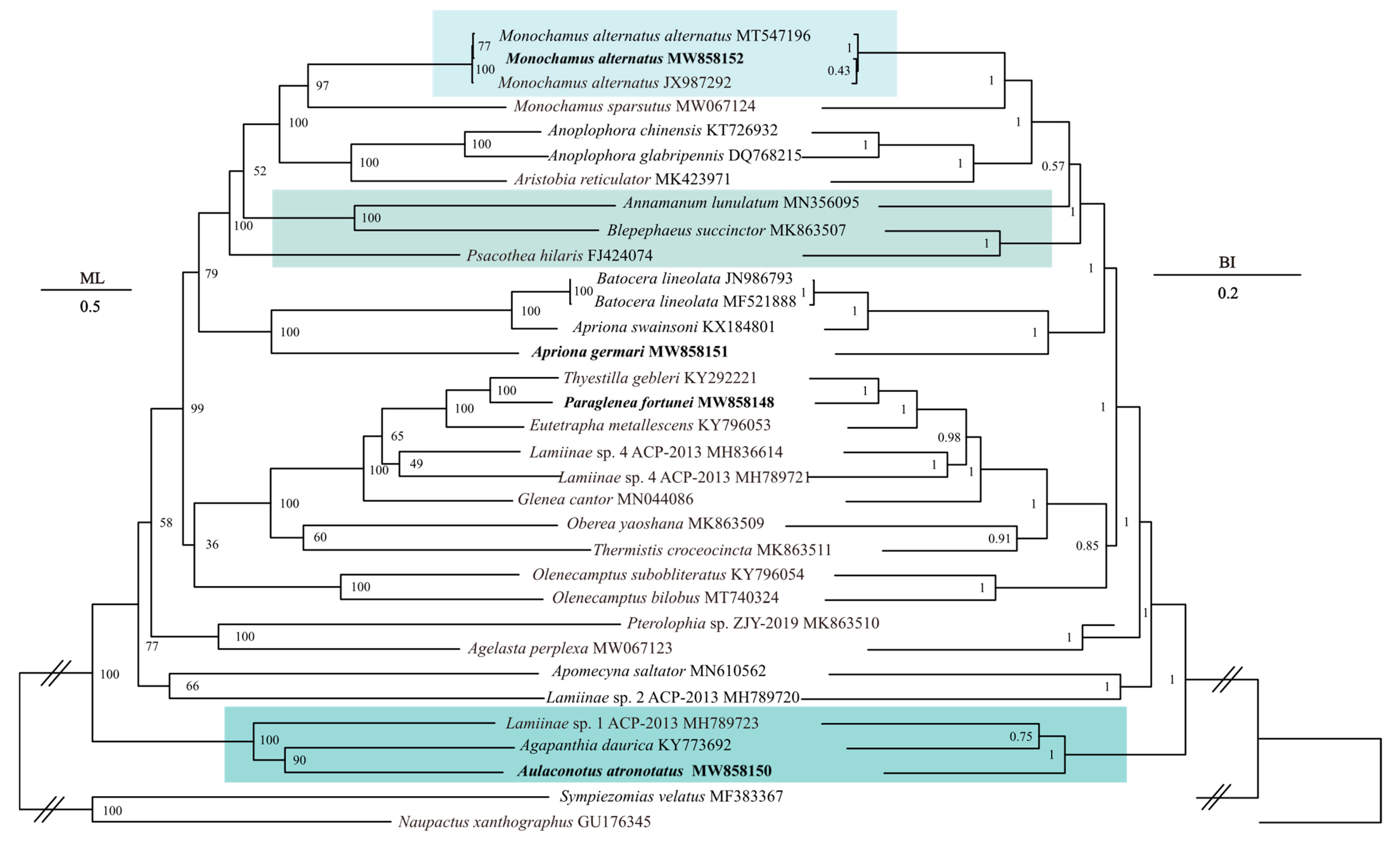
3.3. Phylogenetic Analyses
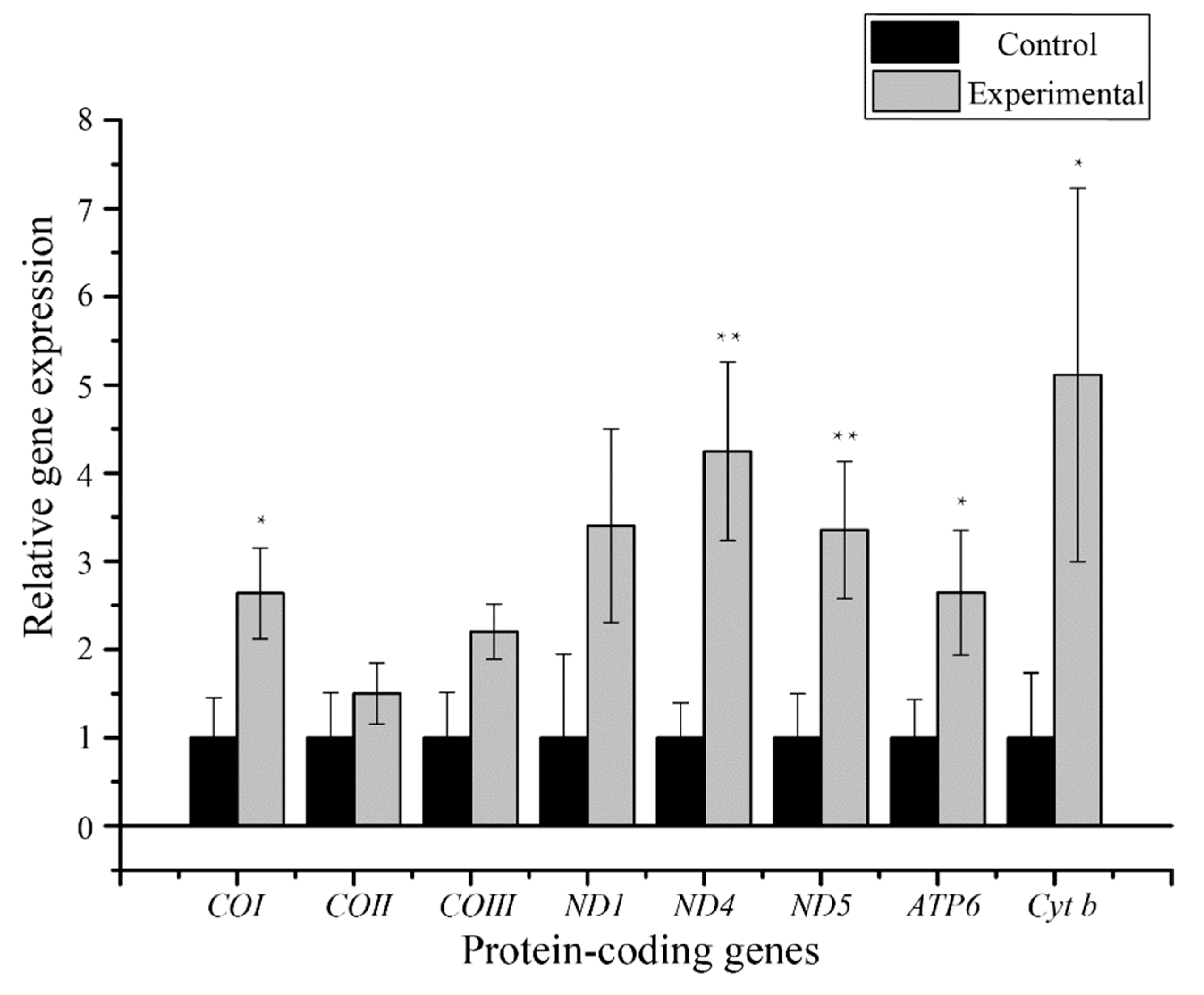
3.4. Transcriptions of Mitochondrial Protein-Coding Genes
4. Discussion
4.1. Phylogenetic Analyses
4.2. Mitochondrial Gene Expression
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nie, R.; Vogler, A.P.; Yang, X.K.; Lin, M.Y. Higher-level phylogeny of longhorn beetles (Coleoptera: Chrysomeloidea) inferred from mitochondrial genomes. Syst. Entomol. 2020, 46, 56–70. [Google Scholar] [CrossRef]
- Wang, J.; Dai, X.Y.; Xu, X.D.; Zhang, Z.Y.; Yu, D.N.; Storey, K.B.; Zhang, J.Y. The complete mitochondrial genomes of five longicorn beetles (Coleoptera: Cerambycidae) and phylogenetic relationships within Cerambycidae. PeerJ 2019, 7, e7633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santana Souza, D.; Marinoni, L.; Monné, M.L.; Gómez-Zurita, J. Molecular phylogenetic assessment of the tribal classification of Lamiinae (Coleoptera: Cerambycidae). Mol. Phylogenet. Evol. 2020, 145, 106736. [Google Scholar] [CrossRef]
- Lin, M.Y.; Tavakilian, G.; Montreuil, O.; Yang, X.K. A study on the Indiana & galathea species-group of the genus Glenea, with descriptions of four new species (Coleoptera: Cerambycidae: Lamiinae: Saperdini). Ann. Soc. Entomol. Franc. 2009, 45, 157–176. [Google Scholar]
- Wang, W.K.; Jiang, S.N. Taxonomic Study on the Chinese Uraecha Thomson (Coleoptera: Cerambycidae: Lamiinae). Entomotaxonomia 2000, 22, 45–47. [Google Scholar]
- Hubweber, L.; Schmitt, M. Differences in genitalia structure and function between subfamilies of longhorn beetles (Coleoptera: Cerambycidae). Genetica 2010, 138, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; An, S.H.; Zhang, H.F.; Wei, Z.H.; Yin, X.M. Study on mitochondrial CO II gene characters and its phylogenetic analysis of Lamiinae partial species. J. Henan Agric. Univ. 2013, 47, 715–721. (In Chinese) [Google Scholar]
- Buermans, H.P.J.; Dunnen, J.T.D. Next generation sequencing technology: Advances and applications. BBA-Mol. Basis Dis. 2014, 1842, 1932–1941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Napp, D.S. Phylogenetic relationships among the subfamilies of Cerambycidae (Coleoptera, Chrysomeloidea). Rev. Bras. Entomol. 1994, 38, 265–419. [Google Scholar]
- Haddad, S.; Shin, S.; Lemmon, A.R.; Lemmon, E.M.; Svacha, P.; Farrell, B.; Ślipiński, A.; Windsor, D.; Mckenna, D.D. Anchored hybrid enrichment provides new insights into the phylogeny and evolution of longhorned beetles (Cerambycidae). Syst. Entomol. 2018, 43, 68–89. [Google Scholar] [CrossRef]
- Dai, X.Y.; Zhang, H.; Xu, X.D.; Jia, Y.Y.; Zhang, J.Y.; Yu, D.N.; Cheng, H.Y. The complete mitochondrial genome of Annamanum lunulatum (Coleoptera: Lamiinae) and its phylogeny. Mitochondrial DNA Part B 2020, 5, 551–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.C.; Lee, S.J.; Kim, S.; Lee, M.R.; Baek, S.; Park, S.E.; Kim, J.; Shin, T.Y.; Kim, J.S. Management of pine wilt disease vectoring Monochamus alternatus adults using spray and soil application of Metarhizium anisopliae JEF isolates. J. Asia Pac. Entomol. 2020, 23, 224–233. [Google Scholar] [CrossRef]
- Shah, K.; Alqudah, M.A.; Jarad, F.; Abdeljawad, T. Semi-analytical study of Pine Wilt Disease model with convex rate under Caputo–Febrizio fractional order derivative. Chaos. Soliton. Fract. 2020, 135, 109754. [Google Scholar] [CrossRef]
- Yan, X.; Cheng, X.Y.; Wang, Y.S.; Luo, J.; Mao, Z.C.; Ferris, V.; Xie, B.Y. Comparative transcriptomics of two pathogenic pinewood nematodes yields insights into parasitic adaptation to life on pine hosts. Gene 2012, 505, 81–90. [Google Scholar] [CrossRef]
- Yasuharu, M.; Nobuo, E. Bursaphelenchus mucronatus n. sp. (Nematoda: Aphelenchoididae) from pine wood and its biology and pathogenicity to pine trees. Nematologica 1979, 25, 353–361. [Google Scholar]
- Kulinich, O.A.; Kruglic, I.A.; Eroshenko, A.S.; Kolosova, N.V. Occurrence and distribution of the nematode Bursaphelenchus mucronatus in the Russian Far East. Russ. J. Nematol. 1994, 2, 113–119. [Google Scholar]
- De Guiran, G.; Bruguier, N. Hybridization and phylogeny of the pine wood nematode (B. ursaphelenchus Spp.). Nematologica 1989, 35, 321–330. [Google Scholar] [CrossRef]
- Wu, S.Y.; Gao, S.H.; Wang, S.; Meng, J.; Wickham, J.; Luo, S.N.; Tan, X.Y.; Yu, H.Y.; Xiang, Y.J.; Hu, S.N.; et al. A reference genome of Bursaphelenchus mucronatus provides new resources for revealing its displacement by pinewood nematode. Genes 2020, 11, 570. [Google Scholar] [CrossRef]
- Xu, H.M.; Chen, J.Y.; Wang, Y.X.; Wang, R.W. Progress of Bursaphelenchus mucronatus. Hubei For. Sci. Technol. 2015, 44, 40–44. (In Chinese) [Google Scholar]
- Ren, B.Z.; Tang, L.H. Progress in quarantine pests of Bursaphelenchus xylophilus. J. Jilin Agric. Univ. 2020, 42, 8–13. [Google Scholar]
- Fukuda, K. Physiological process of the symptom development and resistance mechanism in pine wilt disease. J. For. Res. 1997, 2, 171–181. [Google Scholar] [CrossRef]
- Chong, R.A.; Mueller, R.L. Low metabolic rates in salamanders are correlated with weak selective constraints on mitochondrial genes. Evol. Int. J. Org. Evol. 2013, 67, 894–899. [Google Scholar] [CrossRef]
- Breda, C.N.D.S.; Davanzo, G.G.; Basso, P.J.; Saraiva Câmara, N.O.; Moraes-Vieira, P.M.M. Mitochondria as central hub of the immune system. Redox Biol. 2019, 26, 101255. [Google Scholar] [CrossRef] [PubMed]
- Moritz, C.; Dowling, T.E.; Brown, W.M. Evolution of animal mitochondrial DNA: Relevance for population biology and systematics. Annu. Rev. Ecol. Syst. 1987, 18, 269–292. [Google Scholar] [CrossRef]
- Boore, J.L. Animal mitochondrial genomes. Nucleic Acids Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avise, J.C.; Arnold, J.; Ball, R.M.; Bermingham, E.; Lamb, T.; Neigel, J.E.; Reeb, C.A.; Saunders, N.C. Intraespecific phylogeography: The mitochondrial DNA bridge between population genetics and systematics. Annu. Rev. Ecol. Syst. 1987, 18, 489–522. [Google Scholar] [CrossRef]
- Zhang, J.Y.; Luu, B.E.; Yu, D.N.; Zhang, L.P.; Al-attar, R.; Storey, K.B. The complete mitochondrial genome of Dryophytes versicolor: Phylogenetic relationship among Hylidae and mitochondrial protein-coding gene expression in response to freezing and anoxia. Int. J. Biol. Macromol. 2019, 132, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Flachs, P.; Sponarova, J.; Kopecky, P.; Horvath, O.; Sediva, A.; Nibbelink, M.; Casteilla, L.; Medrikova, D.; Neckar, J.; Kolar, F.; et al. Mitochondrial uncoupling protein 2 gene transcript levels are elevated in maturating erythroid cells. FEBS Lett. 2007, 581, 1093–1097. [Google Scholar] [CrossRef] [Green Version]
- Rudenok, M.M.; Alieva, A.K.; Starovatykh, J.S.; Nesterov, M.S.; Stanishevskaya, V.A.; Kolacheva, A.A.; Ugryumov, M.V.; Slominsky, P.A.; Shadrina, M.I. Expression analysis of genes involved in mitochondrial biogenesis in mice with MPTP-induced model of Parkinson’s disease. Mol. Genet. Metab. Etab. Rep. 2020, 23, 100584. [Google Scholar] [CrossRef]
- Alves, J.P.M.; Fernandes, C.C.L.; Calderón, C.E.M.; Rossetto, R.; Bertolini, M.; Rondina, D. Short-term supplementation of diets rich in lipids or glycogen precursors can affect intra-follicular environment, oocyte mitochondrial gene expression, and embryo development following parthenogenesis in goat. Small Ruminant Res. 2021, 194, 106279. [Google Scholar] [CrossRef]
- Zhou, J.; Zhao, L.L.; Yu, H.Y.; Wang, Y.H.; Zhang, W.; Hu, S.N.; Zou, Z.; Sun, J.H. Immune tolerance of vector beetle to its partner plant parasitic nematode modulated by its insect parasitic nematode. FASEB J. 2018, 32, 4862–4877. [Google Scholar] [CrossRef] [PubMed]
- Ning, J.; Zhang, B.; Tian, H.K.; Liu, X.L.; Yang, B.Y.; Zhao, L.L. Comparative analysis of microRNA profile in Monochamus alternatus in the presence or absence of the pinewood nematode, Bursaphelenchus xylophilus. J. Biosaf. 2018, 27, 20–30. (In Chinese) [Google Scholar]
- Li, M.Y.; Zhou, J.; Wang, H.X.; Zhao, L.L. Transcriptome analysis of differentially expressed genes in Monochamus alternatus (Coleoptera: Cerambycidae) carrying and without carrying pine wood nematode (Bursaphelenchus xylophilus). Acta Entomol. Sin. 2020, 63, 207–217. [Google Scholar]
- Smeitink, J.; Lambert, V.D.H.; Dimauro, S. The genetics and pathology of oxidative phosphorylation. Nat. Rev. Genet. 2001, 2, 342–352. [Google Scholar] [CrossRef]
- Cheng, X.F.; Zhang, L.P.; Yu, D.N.; Storey, K.B.; Zhang, J.Y. The complete mitochondrial genomes of four cockroaches (Insecta: Blattodea) and phylogenetic analyses within cockroaches. Gene 2016, 586, 115–122. [Google Scholar] [CrossRef]
- Gao, X.Y.; Cai, Y.Y.; Yu, D.N.; Storey, K.B.; Zhang, J.Y. Characteristics of the complete mitochondrial genome of Suhpalacsa longialata (Neuroptera, Ascalaphidae) and its phylogenetic implications. PeerJ 2018, 6, e5914. [Google Scholar] [CrossRef] [Green Version]
- Simon, C.; Buckley, T.R.; Frati, F.; Stewart, J.B.; Beckenbach, A.T. Incorporating molecular evolution into phylogenetic analysis, and a new compilation of conserved polymerase chain reaction primers for animal mitochondrial DNA. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 545–579. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.Y.; Zhou, C.F.; Gai, Y.H.; Song, D.X.; Zhou, K.Y. The complete mitochondrial genome of Parafronurus youi (Insecta: Ephemeroptera) and phylogenetic position of the Ephemeroptera. Gene 2008, 424, 18–24. [Google Scholar] [CrossRef]
- Zhang, L.P.; Cai, Y.Y.; Yu, D.N.; Storey, K.B.; Zhang, J.Y. Gene characteristics of the complete mitochondrial genomes of Paratoxodera polyacantha and Toxodera hauseri (Mantodea: Toxoderidae). PeerJ 2018, 6, e4595. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.P.; Yu, D.N.; Storey, K.B.; Cheng, H.Y.; Zhang, J.Y. Higher tRNA gene duplication in mitogenomes of praying mantises (Dictyoptera, Mantodea) and the phylogeny within Mantodea. Int. J. Biol. Macromol. 2018, 111, 787–795. [Google Scholar] [CrossRef]
- Sultana, T.; Han, H.; Park, J.K. Comparison of complete mitochondrial genomes of pine wilt nematode Bursaphelenchus xylophilus and Bursaphelenchus mucronatus (Nematoda: Aphelenchoidea) and development of a molecular tool for species identification. Gene 2013, 520, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Burland, T.G. Methods in Molecular Biology: DNASTAR’s Lasergene Sequence Analysis Software. In Bioinformatics Methods and Protocols; Misener, S., Krawetz, S.A., Eds.; Humana Press: Totowa, NJ, USA, 1999; pp. 71–91. [Google Scholar]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenet. Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef]
- Wang, C.Y.; Feng, Y.; Chen, X.M. Complete coding region of the mitochondrial genome of Monochamus alternatus Hope (Coleoptera: Cerambycidae). Zool. Sci. 2013, 30, 570–576. [Google Scholar] [CrossRef]
- Liao, Q.L.; Yang, X.Z.; Ma, J.Y.; Sheng, L.J.; Zou, S.Q. The complete mitochondrial genome of Monochamus alternatus alternatus (Coleoptera: Cerambycidae). Mitochondrial DNA Part B 2020, 5, 3399–3400. [Google Scholar] [CrossRef] [PubMed]
- Li, W.B.; Yang, X.J.; Qian, L.; An, Y.L.; Fang, J. The complete mitochondrial genome of the citrus long-horned beetle, Anoplophora chinensis (Coleoptera: Cerambycidae). Mitochondrial DNA Part A 2016, 27, 4665–4667. [Google Scholar] [CrossRef]
- Behere, G.T.; Tay, W.T.; Firake, D.M.; Kunz, D.; Burange, P.S.; Ramamurthy, V.V. Characterization of draft mitochondrial genome of guava trunk borer, Aristobia reticulator (Fabricius, 1781) (Coleoptera: Cerambycidae: Lamiinae) from India. Mitochondrial DNA Part B 2019, 4, 1592–1593. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.G.; Hong, M.Y.; Kim, M.J.; Im, H.H.; Kim, M.I.; Bae, C.H.; Seo, S.J.; Lee, S.H.; Kim, I. Complete mitochondrial genome sequence of the yellow-spotted long-horned beetle Psacothea hilaris (Coleoptera: Cerambycidae) and phylogenetic analysis among coleopteran insects. Mol. Cells 2009, 27, 429–441. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.H.; Jia, P.F.; Luo, T.; Wang, Q.M. Complete mitochondrial genome of white-striped long-horned beetle, Batocera lineolata (Coleoptera: Cerambycidae) by next-generation sequencing and its phylogenetic relationship within superfamily Chrysomeloidea. Mitochondrial DNA Part B 2017, 2, 520–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crampton-Platt, A.; Timmermans, M.J.T.N.; Gimmel, M.L.; Kutty, S.N.; Cockerill, T.D.; Chey, V.K.; Vogler, A.P. Soup to tree: The phylogeny of beetles inferred by mitochondrial metagenomics of a bornean rainforest sample. Mol. Biol. Evol. 2015, 32, 2302–2316. [Google Scholar] [CrossRef]
- Tang, P.A.; Zhang, L.; Li, X.P.; Li, F.F.; Yuan, M.L. The complete mitochondrial genome of Sympiezomias velatus (Coleoptera: Curculionidae). Mitochondrial DNA Part B 2017, 2, 449–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, H.J.; Sheffield, N.C.; Cameron, S.L.; Miller, K.B.; Whiting, M.F. When phylogenetic assumptions are violated: Base compositional heterogeneity and among-site rate variation in beetle mitochondrial phylogenomics. Syst. Entomol. 2010, 35, 429–448. [Google Scholar] [CrossRef]
- Cameron, S.L. How to sequence and annotate insect mitochondrial genomes for systematic and comparative genomics research. Syst. Entomol. 2014, 39, 400–411. [Google Scholar] [CrossRef] [Green Version]
- Perna, N.T.; Kocher, T.D. Patterns of nucleotide composition at fourfold degenerate sites of animal mitochondrial genomes. J. Mol. Evol. 1995, 41, 353–358. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.L.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.P.; Ma, Y.; Yu, D.N.; Storey, K.B.; Zhang, J.Y. The mitochondrial genomes of Statilia maculata and S. nemoralis (Mantidae: Mantinae) with different duplications of trnR genes. Int. J. Biol. Macromol. 2019, 121, 839–845. [Google Scholar] [CrossRef] [PubMed]
- Castresana, J. Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. Partitionfinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Mark, P.V.D.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguilar, O.A.; Hadj-Moussa, H.; Storey, K.B. Freeze-responsive regulation of MEF2 proteins and downstream gene networks in muscles of the wood frog, Rana sylvatica. J. Therm. Biol. 2017, 67, 1–8. [Google Scholar] [CrossRef]
- Lalitha, S. Primer Premier 5. Biotech Softw. Internet Rep. 2004, 1, 270–272. [Google Scholar] [CrossRef]
- Li, H.; He, X.Y.; Tao, R.; Chen, H.J.; Gong, X.Y.; Li, S.Y.; Hao, D.J. cDNA cloning and expression profiling of small heat shock protein genes and their response to temperature stress in Monochamus alternatus (Coleoptera: Cerambycidae). Acta Entomol. Sin. 2018, 61, 749–760. [Google Scholar]
- Dorn, G.W.; Vega, R.B.; Kelly, D.P. Mitochondrial biogenesis and dynamics in the developing and diseased heart. Genes Dev. 2015, 29, 1981–1991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehrabian, Z.; Liu, L.I.; Fiskum, G.; Rapoport, S.; Chandrasekaran, K. Regulation of mitochondrial gene expression by energy demand in neural cells. J. Neurochem. 2005, 93, 850–860. [Google Scholar] [CrossRef]
- Sousa, J.S.; D’Imprima, E.; Vonck, J. Mitochondrial respiratory chain complexes. In Membrane Protein Complexes: Structure and Function. Subcellular Biochemistry; Harris, J., Boekema, E., Eds.; Springer: Singapore, 2018; Volume 87, pp. 167–227. [Google Scholar]
- Ott, M.; Amunts, A.; Brown, A. Organization and regulation of mitochondrial protein synthesis. Annu. Rev. Biochem. 2016, 85, 77–101. [Google Scholar] [CrossRef]
- Janssen, R.J.R.J.; Nijtmans, L.G.; Heuvel, L.P.V.D.; Smeitink, J.A.M. Mitochondrial complex I: Structure, function and pathology. J. Inherit. Metab. Dis. 2006, 29, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Storey, K.B. Anoxia-induced gene expression in turtle heart: Upregulation of mitochondrial genes for NADH-ubiquinone oxidoreductase subunit 5 and cytochrome c oxidase subunit 1. Eur. J. Biochem. 1996, 241, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Ndi, M.; Marin-Buera, L.; Salvatori, R.; Singh, A.P.; Ott, M. Biogenesis of the bc 1 complex of the mitochondrial respiratory chain. J. Mol. Biol. 2018, 430, 3892–3905. [Google Scholar] [CrossRef]
- Bourens, M.; Barrientos, A. A CMC1-knockout reveals translation-independent control of human mitochondrial complex IV biogenesis. EMBO Rep. 2017, 18, 477–494. [Google Scholar] [CrossRef] [Green Version]
- Wikström, M.; Krab, K.; Sharma, V. Oxygen activation and energy conservation by cytochrome c oxidase. Chem. Rev. 2018, 118, 2469–2490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mathur, D.; López-Rodas, G.; Casanova, B.; Marti, M.B. Perturbed glucose metabolism: Insights into multiple sclerosis pathogenesis. Front. Nurol. 2014, 5, 250. [Google Scholar] [CrossRef] [PubMed]
- Weinberg, S.E.; Sena, L.A.; Chandel, N.S. Mitochondria in the regulation of innate and adaptive immunity. Immunity 2015, 42, 406–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosomi, K.; Kunisawa, J. Diversity of energy metabolism in immune responses regulated by micro-organisms and dietary nutrition. Int. Immunol. 2020, 32, 447–454. [Google Scholar] [CrossRef]
- Mehta, M.M.; Weinberg, S.E.; Chandel, N.S. Mitochondrial control of immunity: Beyond ATP. Nat. Rev. Immunol. 2017, 17, 608. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Yu, H.Y.; Zhang, W.; Ahmad, F.; Hu, S.N.; Zhao, L.L.; Zou, Z.; Sun, J.H. Comparative analysis of the Monochamus alternatus immune system. Insect Sci. 2018, 25, 581–603. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.L.; Zhang, X.X.; Wei, Y.N.; Zhou, J.; Zhang, W.; Qin, P.J.; Chinta, S.; Kong, X.B.; Liu, Y.P.; Yu, H.Y.; et al. Ascarosides coordinate the dispersal of a plant-parasitic nematode with the metamorphosis of its vector beetle. Nat. Commun. 2016, 7, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, W.E.; Herrmann, M.; Sommer, R.J. Molecular phylogeny of beetle associated diplogastrid nematodes suggests host switching rather than nematode-beetle coevolution. BMC Evol. Biol. 2009, 9, 1–11. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Subamily | Genus | Species | GenBank No. | References |
|---|---|---|---|---|
| Lamiinae | Monochamus | Monochamus alternatus alternatus | MT547196 | Liao (2020) |
| Monochamus alternatus | MW858152 | This study | ||
| Monochamus alternatus | JX987292 | Wang (2012) | ||
| Monochamus sparsutus | MW067124 | Wu (2020) | ||
| Anoplophora | Anoplophora chinensis | KT726932 | Li (2015) | |
| Anoplophora glabripennis | DQ768215 | An (2006) | ||
| Aristobia | Aristobia reticulator | MK423971 | Behere (2019) | |
| Psacothea | Psacothea hilaris | FJ424074 | Kim (2009) | |
| Annamanum | Annamanum lunulatum | MN356095 | Dai (2020) | |
| Blepephaeus | Blepephaeus succinctor | MK863507 | Wang (2019) | |
| Batocera | Batocera lineolata | JN986793 | Wang (2011) | |
| Batocera lineolata | MF521888 | Liu (2017) | ||
| Apriona | Apriona swainsoni | KX184801 | Que (2016) | |
| Apriona germari | MW858151 | This study | ||
| Thyestilla | Thyestilla gebleri | KY292221 | Yang (2016) | |
| Paraglenea | Paraglenea fortunei | MW858148 | This study | |
| Eutetrapha | Eutetrapha metallescens | KY796053 | Yang (2017) | |
| Lamiinae sp. 4 ACP-2013 | MH836614 | Crampton-Platt (2015) | ||
| Lamiinae sp. 4 ACP-2013 | MH789721 | Crampton-Platt (2015) | ||
| Glenea | Glenea cantor | MN044086 | Wang (2019) | |
| Oberea | Oberea yaoshana | MK863509 | Wang (2019) | |
| Thermisti | Thermistis croceocincta | MK863511 | Wang (2019) | |
| Olenecamptus | Olenecamptus subobliteratus | KY796054 | Yang (2017) | |
| Olenecamptus bilobus strain fentianniu | MT740324 | Dong (2020) | ||
| Pterolophia | Pterolophia sp. ZJY-2019 | MK863510 | Wang (2019) | |
| Agelasta | Agelasta perplexa | MW067123 | Chen (2020) | |
| Apomecyna | Apomecyna saltator | MN610562 | Nie (2021) | |
| Lamiinae sp. 2 ACP-2013 | MH789720 | Crampton-Platt (2015) | ||
| Lamiinae sp. 1 ACP-2013 | MH789723 | Crampton-Platt (2015) | ||
| Agapanthia | Agapanthia daurica | KY773692 | Yang (2017) | |
| Aulaconotus | Aulaconotus atronotatus | MW858150 | This study | |
| naupactus | Naupactus xanthographus | GU176345 | Song (2010) | |
| sympiezomias | Sympiezomias velatus | MF383367 | Tang (2017) |
| Nucleotide Sequence Alignments | ||
|---|---|---|
| Subset | Subset Partitions | Best Model |
| Partition 1 | COX1_codon1, COX2_codon1, ATP6_codon1, COX3_codon1, CYTB_codon1 | GTR + I + G |
| Partition 2 | COX1_codon2, COX3_codon2, ATP6_codon2, COX2_codon2, CYTB_codon2 | TVM + I + G |
| Partition 3 | COX1_codon3, COX2_codon3, ATP6_codon3, ATP8_codon3 | TIM + G |
| Partition 4 | ATP8_codon2, ND2_codon1, ND3_codon1, ND6_codon1, ATP8_codon1 | GTR + I + G |
| Partition 5 | ND6_codon3, COX3_codon3, ND3_codon3, CYTB_codon3 | HKY + G |
| Partition 6 | ND4L_codon1, ND1_codon1, ND5_codon1, ND4_codon1 | TVM + I + G |
| Partition 7 | ND4L_codon2, ND1_codon2, ND5_codon2, ND4_codon2 | GTR + I + G |
| Partition 8 | ND1_codon3 | HKY + G |
| Partition 9 | ND6_codon2, ND2_codon2, ND3_codon2 | TVM + I + G |
| Partition 10 | ND2_codon3 | TRN + G |
| Partition 11 | ND5_codon3, ND4_codon3, ND4L_codon3 | HKY + G |
| Gene | Forward Primer | Forward Primer |
|---|---|---|
| β-actin | CTCAACCCCAAGGCTAACC | CACCATCTCCAGAGTCCAAT |
| COI | CTC(T)TTACCTCCTTCTTTAACTC | CAACTGATGAACCTCTATGAG |
| COII | GATGCAACTCCTGGACGAT | ATCTATGATTGGCACCACAA |
| COIII | AGAGCCTTATCTCCTAGAATTG | GCTCAAGTTACTGTTAATCCTG |
| ND1 | ATTATC(T)GCAAATCCACCTCT | TAGCAGAAACTAATCGTACTCC |
| ND4 | GAAGGAGGAGCAGCCATA | CTTCAGGTTTATTTTGTTTAGC |
| ND5 | TAGTAAAGCAACATCCCCA | TATTAGGGTGAGATGGTTTAG |
| ATP6 | TTAGTACCTCAAGGAACTCC | GATAATCGAACTGCCAATGT |
| Cyt b | ATCATTCTGAGGAGCAACTG | TGAAAGGTAAAAAATCGTGT |
| Species | A + T (%) | AT-Skew | GC-Skew | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Mito | PCGs | rRNAs | tRNAs | Mito | PCGs | rRNAs | tRNAs | Mito | PCGs | rRNAs | tRNAs | |
| A. atronotatus | 74.2 | 73.5 | 76.8 | 78.5 | 0.019 | −0.15 | −0.052 | 0.029 | −0.0222 | −0.034 | 0.401 | 0.133 |
| A. germari | 76.8 | 76.3 | 77.7 | 77.2 | 0.013 | −0.151 | −0.028 | 0.036 | −0.0208 | −0.002 | 0.409 | 0.154 |
| P. fortunei | 74.3 | 73 | 75.7 | 76 | 0.038 | −0.152 | −0.062 | 0.031 | −0.287 | −0.014 | 0.472 | 0.116 |
| M. alternatus | 78.4 | 77.7 | 82.1 | 79.9 | 0.006 | −0.152 | −0.023 | 0.034 | −0.153 | 0.026 | 0.342 | 0.112 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.-Y.; Guan, J.-Y.; Cao, Y.-R.; Dai, X.-Y.; Storey, K.B.; Yu, D.-N.; Zhang, J.-Y. Mitogenome Analysis of Four Lamiinae Species (Coleoptera: Cerambycidae) and Gene Expression Responses by Monochamus alternatus When Infected with the Parasitic Nematode, Bursaphelenchus mucronatus. Insects 2021, 12, 453. https://doi.org/10.3390/insects12050453
Zhang Z-Y, Guan J-Y, Cao Y-R, Dai X-Y, Storey KB, Yu D-N, Zhang J-Y. Mitogenome Analysis of Four Lamiinae Species (Coleoptera: Cerambycidae) and Gene Expression Responses by Monochamus alternatus When Infected with the Parasitic Nematode, Bursaphelenchus mucronatus. Insects. 2021; 12(5):453. https://doi.org/10.3390/insects12050453
Chicago/Turabian StyleZhang, Zi-Yi, Jia-Yin Guan, Yu-Rou Cao, Xin-Yi Dai, Kenneth B. Storey, Dan-Na Yu, and Jia-Yong Zhang. 2021. "Mitogenome Analysis of Four Lamiinae Species (Coleoptera: Cerambycidae) and Gene Expression Responses by Monochamus alternatus When Infected with the Parasitic Nematode, Bursaphelenchus mucronatus" Insects 12, no. 5: 453. https://doi.org/10.3390/insects12050453
APA StyleZhang, Z. -Y., Guan, J. -Y., Cao, Y. -R., Dai, X. -Y., Storey, K. B., Yu, D. -N., & Zhang, J. -Y. (2021). Mitogenome Analysis of Four Lamiinae Species (Coleoptera: Cerambycidae) and Gene Expression Responses by Monochamus alternatus When Infected with the Parasitic Nematode, Bursaphelenchus mucronatus. Insects, 12(5), 453. https://doi.org/10.3390/insects12050453