
Morphological, Ecological, and Molecular Divergence of Conogethes pinicolalis from C. punctiferalis (Lepidoptera: Crambidae)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
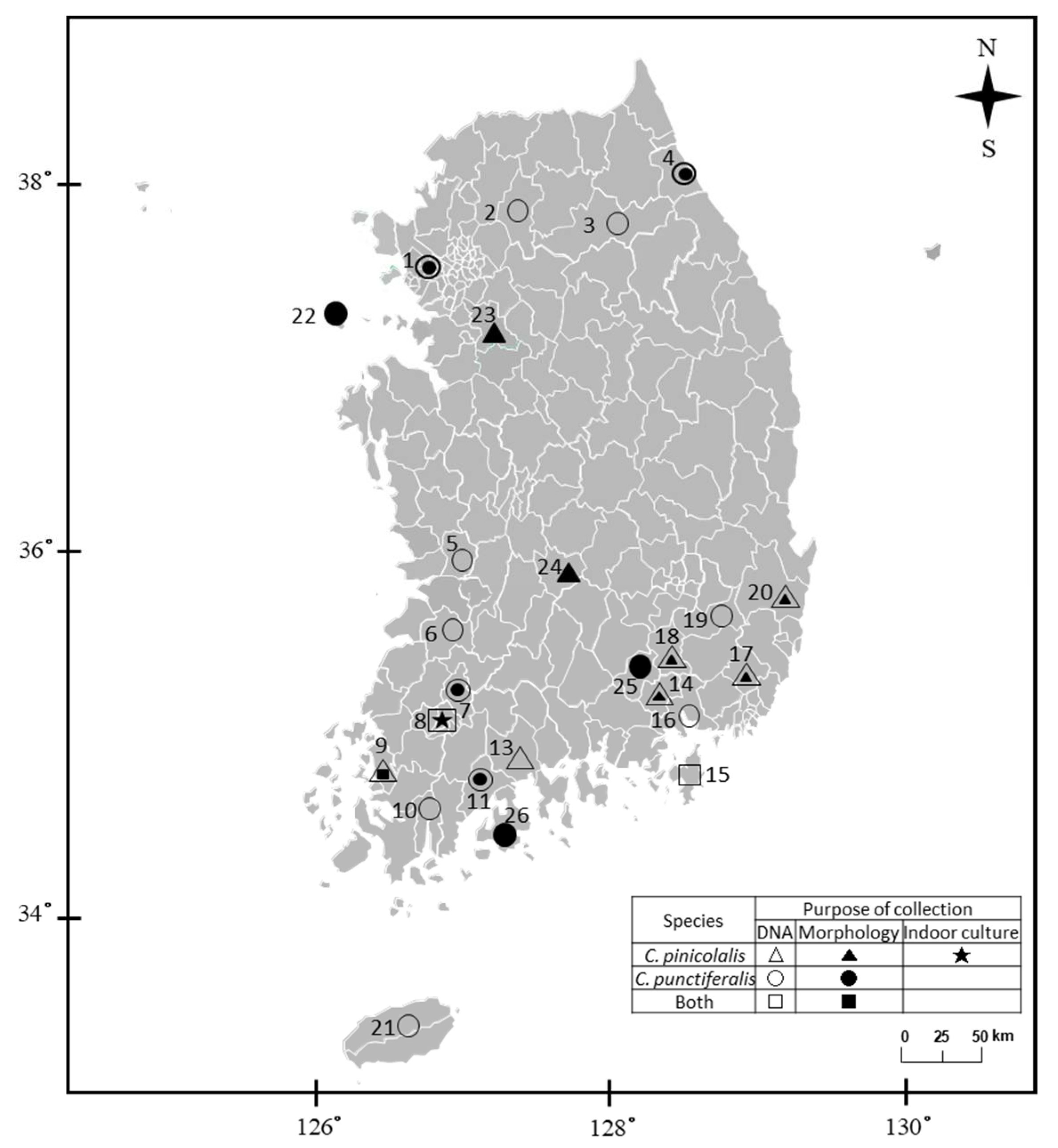
2.1. Sampling
2.2. Insect Rearing
2.3. Genomic DNA Extraction
2.4. PCR and Sequencing
2.5. Sequence Analysis
2.6. Phylogenetic Analysis
2.7. Population Genetic Structure
3. Results
3.1. Biology
3.2. Systematics
3.3. Molecular Analyses
3.3.1. Haplotype Diversity
3.3.2. Haplotype Relationships
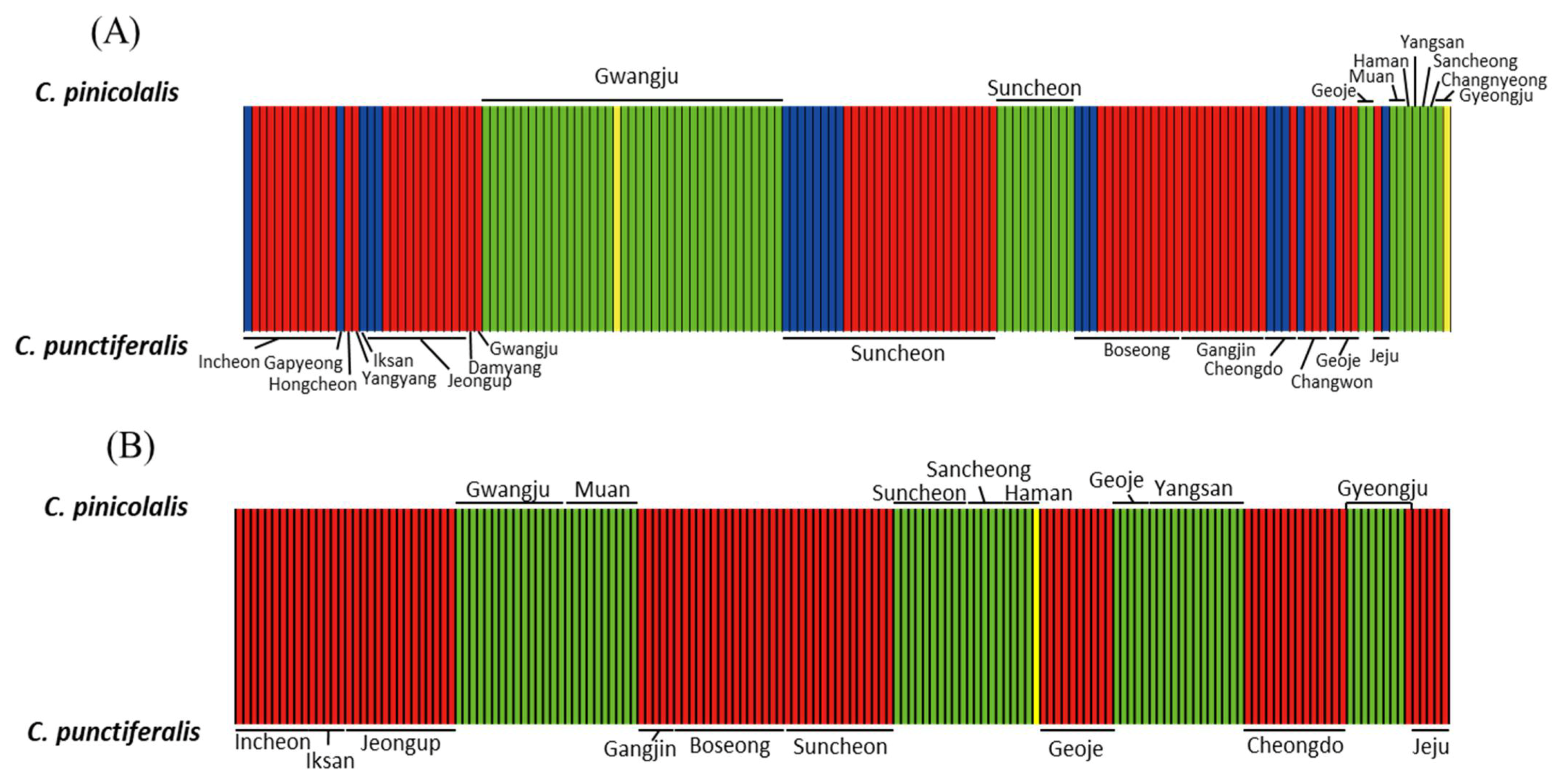
3.3.3. Population Structure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Shaffer, M.; Nielsen, E.S.; Horak, M. Checklist of the Lepidoptera of Australia: Pyraloidea; CSIRO: Melbourne, Australia, 1996.
- Nuss, M.; Landry, B.; Mally, R.; Vegliante, F.; Tränkner, A.; Bauer, F.; Hayden, J.; Segerer, A.; Schouten, R.; Li, H.; et al. GlobiZ–Global Information System on Pyraloidea, 2003–2021. Available online: www.pyraloidea.org (accessed on 4 May 2021).
- Mally, R. Moths of the Genus Conogethes: Taxonomy, Systematics and similar species. In The Black Spotted, Yellow Borer, Conogethes Punctiferalis Guenee and Allied Species; Springer: Singapore, 2018; pp. 1–12. [Google Scholar]
- Inoue, H.; Yamanaka, H. Redescription of Conogethes punctiferalis (Guenée) and description of two new closely allied species from Eastern Palaearctic and Oriental regions (Pyralidae, Pyraustinae). Tinea 2006, 19, 80–91. Available online: https://iss.ndl.go.jp/books/R000000004-I8513605-00 (accessed on 4 May 2021).
- Sekiguchi, K. Morphology, biology and control of the yellow peach moth, Dichocrosis punctiferalis Guenée (Lepidoptera, Pyraustidae). Bull. Ibaraki ken Hortic. Exp. Stn. 1974, 1–90, Special Issue. Available online: https://agris.fao.org/agris-search/search.do?recordID=JP19750017024 (accessed on 4 May 2021).
- Waterhouse, D.F. The Major Arthropod Pests and Weeds of Agriculture in Southeast Asia: Distribution, Importance and Origin; Australian Centre for International Agricultural Research (ACIAR): Canberra, Australia, 1993.
- Kang, C.H.; Lee, S.M.; Chung, Y.J.; Choi, K.S.; Park, C.G. Overwintering ecology of the peach pyralid moth, Dichocrosis punctiferalis in southern regions of Korea. Korean J. Appl. Entomol. 2004, 43, 201–209. Available online: http://entomology2.or.kr/journal/article.php?code=10886 (accessed on 4 May 2021). [CrossRef]
- Kang, C.H.; Lee, K.C.; Park, C.G.; Lee, D.W. Seasonal occurrence pattern of peach pyralid moth, Dichocrocis punctiferalis, in fuyu persimmon orchards and fruit damage at harvesting time. Korean J. Appl. Entomol. 2002, 41, 107–112. Available online: http://entomology2.or.kr/journal/article.php?code=10776 (accessed on 4 May 2021).
- Choi, K.S.; Han, K.S.; Jeon, M.J.; Chung, Y.J.; Kim, C.S.; Shin, S.C.; Park, J.D.; Boo, K.S. Seasonal occurrence of the peach pyralid moth, Dichocrocis punctiferalis at chestnut orchards in some Provinces of Korea. J. Korean Soc. For. Sci. 2004, 93, 134–139. Available online: http://kiss.kstudy.com/thesis/thesis-view.asp?key=2264224 (accessed on 4 May 2021).
- Lee, C.K. The changes of damage rate by peach pyralid moth, Dichocrocis punctiferalis (Lepidoptera: Pyralidae) in chestnut orchards from 1995 to 2010 year. Korean J. Appl. Entomol. 2011, 50, 379–382. [Google Scholar] [CrossRef]
- Koizumi, K. Yellow peach moth, Dichocrocis punctiferalis Guenée. Kobe Plant Prot. News. 1963, 323, 58. [Google Scholar]
- Honda, H. Bio systematics of Japanese Conogethes spp. In With Special Reference to Host Plant Preference and Reproductive Isolation; Chakravarthy, A.K., Ashok Kumar, C.T., Varghese, A., Thyagaraj, N.E., Eds.; New horizons in insect science. Abstracts ICIS-2013; Springer: Bangalore, Indian, 2013. [Google Scholar]
- Konno, Y.; Arai, K.; Sekiguchi, K.; Matsumoto, Y. (E)-10-Hexadecenal, a sex pheromone component of the yellow peach moth, Dichocrocis punctiferalis Guenée (Lepidoptera:Pyralidae). Appl. Entomol. Zool. 1982, 17, 207–217. [Google Scholar] [CrossRef] [Green Version]
- Honda, H.; Mitsuhashi, W. Morphological and morphometrical differences between the fruit- and Pinaceae-feeding type of yellow peach moth, Conogethes punctiferalis (Guenée) (Lepidoptera: Pyralidae). Appl. Entomol. Zool. 1989, 24, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Honda, H. Post-mating reproductive isolation between fruit- and pinaceae-feeding types of the yellow peach moth, Conogethes punctiferalis (Guenée) (Lepidoptera: Pyralidae). Appl. Entomol. Zool. 1986, 21, 489–491. [Google Scholar] [CrossRef] [Green Version]
- Paek, M.K.; Hwang, J.M.; Jung, K.S.; Kim, T.W.; Kim, M.C.; Lee, Y.J.; Cho, Y.B.; Park, S.W.; Lee, H.S.; Ku, D.S.; et al. Checklist of Korean Insects; Nature and Ecology: Seoul, Korea, 2010.
- Park, K.T. Illustrated Flora and Fauna of Korea vol. 27, Insecta IX: Microlepidoptera; Samhwa Publicatioin, Ministry of Education: Seoul, Korea, 1983. [Google Scholar]
- Munroe, E. Changes in classification and names of Hawaiian Pyralidae since the publication of Insects of Hawaii, vol. 8, by Zimmerman, E.C. (1958) (Lepidoptera). Bish. Mus. Occas. Pap. 1989, 29, 199–212. Available online: https://www.cabdirect.org/cabdirect/abstract/19911150642 (accessed on 4 May 2021).
- Honda, H.; Kaneko, J.; Konno, Y.; Matsumoto, Y. A simple method for mass-rearing of the yellow peach moth, Dichocrocis punctiferalis Guenée (Lepidoptera: Pyralidae), on an artificial diet. Appl. Entomol. Zool. 1979, 14, 464–468. [Google Scholar] [CrossRef] [Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. Available online: https://www.mbari.org/wp-content/uploads/2016/01/Folmer_94MMBB.pdf (accessed on 4 May 2021). [PubMed]
- Hundsdöerfer, A.K.; Rubinoff, D.; Attié, M.; Wink, M.; Kitching, I.J. A revised molecular phylogeny of the globally distributed hawkmoth genus Hyles (Lepidoptera: Sphingidae), based on mitochondrial and nuclear DNA sequences. Mol. Phylogenet. Evol. 2009, 52, 852–865. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, A.; Pierce, N.E. Phylogeny of Bicyclus butterflies (Lepidoptera: Nymphalidae) inferred from COI, COII and EF1α gene sequences. Mol. Phylogenet. Evol. 2001, 18, 264–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sievers, F.; Higgins, D.G. Clustal Omega, accurate alignment of very large numbers of sequences. Methods Mol. Biol. 2014, 1079, 105–116. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP* Phylogenetic Analysis Using Parsimony Version 4.10; Sinauer Associates: Sunderland, UK, 2002. [Google Scholar]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; Mark, P.; Ayres, D.L.; Darling, A.; Hohna, S.; Laget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. In Proceedings of the 9th Gateway Computing Environments Workshop (GCE), 14 November 2010; Available online: https://doi.org/10.1109/GCE.2010.5676129 (accessed on 4 May 2021).
- Park, J.S.; Kim, M.J.; Ahn, S.J.; Kim, I. Complete mitochondrial genome of the grass moth Glyphodes quadrimaculalis (Lepidoptera: Crambidae). Mitochondrial DNA Part A 2015, 26, 247–249. [Google Scholar] [CrossRef]
- Mally, R.; Hayden, J.E.; Neinhuis, C.; Jordal, B.H.; Nuss, M. The phylogenetic systematics of Spilomelinae and Pyraustinae (Lepidoptera: Pyraloidea: Crambidae) inferred from DNA and morphology. Arthropod Syst. Phylogeny 2019, 77, 141–204. [Google Scholar] [CrossRef]
- Orlóci, L. Multivariate Analysis in Vegetation Research, 2nd ed.; Junk: The Hague, The Netherlands, 1978. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Corander, J.; Tang, J. Bayesian analysis of population structure based on linked molecular information. Math. Biosci. 2007, 205, 19–31. [Google Scholar] [CrossRef]
- Bae, Y.S.; Byun, B.K.; Paek, M.K. Pyralid Moths of Korea (Lepidoptera, Pyraloidea); National Arboretum: Pocheon, Korea, 2008.
- Inoue, H. Pyralidae. In Moths of Japan Volume I.; Inoue, H., Sugi, S., Kuroko, H., Moriuti, S., Kawabe, A., Eds.; Kodansha: Tokyo, Japan, 1982; pp. 307–404. [Google Scholar]
- Lee, B.Y.; Chung, Y.J. Insect Pests of Trees and Shrubs in Korea; Sungandang: Seoul, Korea, 1997. [Google Scholar]
- Hong, Y.; Kwon, K.; Kim, Y. Post-harvest treatment on the peach pyralid moth and the small tea tortrix moth infesting apples using controlled atmosphere and temperature. Korean J. Appl. Entomol. 2015, 54, 11–17. [Google Scholar] [CrossRef]
- Armstrong, K. DNA barcoding: A new module in New Zealand′s plant biosecurity diagnostic toolbox. Bull OEPP/EPPO 2010, 40, 91–100. [Google Scholar] [CrossRef]
- Kammar, V.; Shashank, P.R.; Rani, A.T.; Slevanarayan, V. Molecular status of Conogethes spp.: An Overview.pp. 13–22. In The Black Spotted, Yellow Borer, Conogethes Punctiferalis Guenee and Allied Species; Chakravarthy, A.K., Ed.; Springer: Singapore, 2018; p. 347. [Google Scholar]
- Lee, C.K. Study on chestnut insect pests by environmentally friendly controls in Korea. Korean J. Appl. Entomol. 2009, 48, 95–100. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, T.; Wang, Z.; He, K.; Liu, Y.; Li, J. Molecular taxonomy of Conogethes punctiferalis and Conogethes pinicolalis (Lepidoptera: Crambidae) based on mitochondrial DNA sequences. J. Integr. Agric. 2014, 13, 1982–1989. [Google Scholar] [CrossRef]
- Kim, J.S.; Kim, M.J.; Jeong, J.S.; Kim, I. Complete mitochondrial genome of Saturnia jonasii (Lepidoptera:Saturniidae): Genomic comparisons and phylogenetic inference among Bombycoidea. Genomics 2018, 110, 274–282. [Google Scholar] [CrossRef] [PubMed]
- Zink, R.M.; Barrowclough, G.F. Mitochondrial DNA under siege in avian phylogeography. Mol. Ecol. 2008, 17, 2107–2121. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Lee, W.; Lee, S. Estimation of a new molecular marker of the genus Stathmopoda (Lepidoptera: Stathmopodidae): Comparing EF1a and COI sequences. J. Asia-Pac. Entomol. 2017, 20, 269–280. [Google Scholar] [CrossRef]
- Hardwick, D.F. The corn earworm complex. Mem. Entomol. Soc. Can. 1965, 40, 1–247. [Google Scholar] [CrossRef]
- Pogue, M. A new synonym of Helicoverpa zea (Boddie) and differentiation of adult males of H. zea and H. armigera (Hübner) (Lepidoptera: Noctuidae: Heliothinae). Ann. Entomol. Soc. Am. 2004, 97, 1222–1226. [Google Scholar] [CrossRef] [Green Version]
- Nagoshi, R.N.; Gilligan, T.M.; Brambila, J. Combining Tpi and COI genetic markers to discriminate invasive Helicoverpa armigera from local Helicoverpa zea (Lepidoptera: Noctuidae) populations in the Southeastern United States. J. Econ. Entomol. 2016, 109, 2115–2124. [Google Scholar] [CrossRef] [Green Version]
- Leite, N.A.; Côrrea, A.S.; Alves-Pereira, A.; Campos, J.B.; Zucchi, M.I.; Omoto, C. Cross-species amplification and polymorphism of microsatellite loci in Helicoverpa armigera and Helicoverpa zea (Lepidoptera:Noctuidae) in Brazilian cropping systems. Genet. Mol. Res. 2016, 2, 1–12. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | No. Haplotype | Min. | Max. | Aver. |
|---|---|---|---|---|
| Intraspecific divergence in each Conogethes species | ||||
| Conogethes punctiferalis (current study only) | 15 | 0.15 | 1.53 | 0.58 |
| Conogethes punctiferalis (current study and public data) | 33 | 0.15 | 5.80 | 2.50 |
| Conogethes punctiferalis (current study and public data, excluding Australian haplotypes) | 23 | 0.15 | 1.53 | 0.56 |
| Conogethes pinicolalis | 12 | 0.15 | 1.68 | 0.62 |
| Conogethes pluto | 5 | 0.15 | 0.46 | 0.31 |
| Conogethes semifascialis | 8 | 0.15 | 5.19 | 2.78 |
| Conogethes tharsalea | 4 | 0.15 | 0.61 | 0.43 |
| Conogethes ersealis | 2 | - | - | 0.46 |
| Conogethes evaxalis | 3 | 0.15 | 1.83 | 1.22 |
| Interspecific divergence between Conogethes species | ||||
| C. punctiferalis and C. pinicolalis (current study only) | 4.89 | 6.26 | 5.44 | |
| C. punctiferalis and C. pinicolalis (current study and public data) | 4.89 | 7.02 | 5.76 | |
| C. punctiferalis and C. pinicolalis (current study and public data, excluding Australian haplotypes) | 4.89 | 6.26 | 5.46 | |
| C. punctiferalis and C. pluto | 5.34 | 6.72 | 6.15 | |
| C. punctiferalis and C. semifascialis | 0.15 | 5.80 | 4.25 | |
| C. punctiferalis and C. tharsalea | 9.16 | 10.69 | 10.06 | |
| C. punctiferalis and C. ersealis | 7.18 | 8.55 | 7.72 | |
| C. punctiferalis and C. evaxalis | 7.63 | 10.23 | 8.83 | |
| C. punctiferalis and C. diminutiva | 7.79 | 8.86 | 8.36 | |
| C. punctiferalis and C. haemactalis | 9.31 | 10.08 | 9.63 | |
| C. punctiferalis and C. pandamalis | 5.65 | 7.18 | 6.20 | |
| C. punctiferalis and C. pluto (excluding Australian haplotypes 1) | 5.34 | 6.72 | 6.13 | |
| C. punctiferalis and C. semifascialis (excluding Australian haplotypes 1) | 1.99 | 5.80 | 4.38 | |
| C. punctiferalis and C. tharsalea (excluding Australian haplotypes 1) | 9.16 | 10.53 | 9.93 | |
| C. punctiferalis and C. ersealis (excluding Australian haplotypes 1) | 7.33 | 8.55 | 7.94 | |
| C. punctiferalis and C. evaxalis (excluding Australian haplotypes 1) | 7.79 | 9.47 | 8.61 | |
| C. punctiferalis and C. diminutive (excluding Australian haplotypes 1) | 7.79 | 8.86 | 8.32 | |
| C. punctiferalis and C. haemactalis (excluding Australian haplotypes 1) | 9.31 | 10.08 | 9.63 | |
| C. punctiferalis and C. pandamalis (excluding Australian haplotypes 1) | 5.65 | 6.41 | 5.95 | |
| C. punctiferalis and C. pluto (C. punctiferalis from current study only) | 5.34 | 6.72 | 6.12 | |
| C. punctiferalis and C. semifascialis (C. punctiferalis from current study only) | 1.99 | 5.80 | 4.36 | |
| C. punctiferalis and C. tharsalea (C. punctiferalis from current study only) | 9.16 | 10.53 | 9.92 | |
| C. punctiferalis and C. ersealis (C. punctiferalis from current study only) | 7.33 | 8.55 | 7.96 | |
| C. punctiferalis and C. evaxalis (C. punctiferalis from current study only) | 7.79 | 9.47 | 8.60 | |
| C. punctiferalis and C. diminutive (C. punctiferalis from current study only) | 7.79 | 8.86 | 8.29 | |
| C. punctiferalis and C. haemactalis (C. punctiferalis from current study only) | 9.31 | 10.08 | 9.65 | |
| C. punctiferalis and C. pandamalis (C. punctiferalis from current study only) | 5.65 | 6.41 | 5.91 | |
| C. pinicolalis and C. pluto | 5.95 | 6.72 | 6.51 | |
| C. pinicolalis and C. semifascialis | 5.04 | 6.57 | 5.89 | |
| C. pinicolalis and C. tharsalea | 10.99 | 12.06 | 11.52 | |
| C. pinicolalis and C. ersealis | 6.87 | 7.94 | 7.29 | |
| C. pinicolalis and C. evaxalis | 7.94 | 9.47 | 8.85 | |
| C. pinicolalis and C. diminutive | 7.48 | 8.24 | 7.77 | |
| C. pinicolalis and C. haemactalis | 9.01 | 9.77 | 9.37 | |
| C. pinicolalis and C. pandamalis | 4.89 | 5.80 | 5.43 | |
| C. pluto and C. semifascialis | 6.11 | 6.87 | 6.44 | |
| C. pluto and C. tharsalea | 10.69 | 11.45 | 11.04 | |
| C. pluto and C. ersealis | 6.87 | 7.63 | 7.27 | |
| C. pluto and C. evaxalis | 9.16 | 9.92 | 9.56 | |
| C. pluto and C. diminutiva | 8.24 | 8.70 | 8.43 | |
| C. pluto and C. haemactalis | 9.77 | 10.23 | 10.02 | |
| C. pluto and C. pandamalis | 5.80 | 6.11 | 5.98 | |
| C. semifascialis and C. tharsalea | 9.93 | 11.45 | 10.67 | |
| C. semifascialis and C. ersealis | 6.72 | 8.86 | 8.00 | |
| C. semifascialis and C. evaxalis | 7.48 | 9.92 | 9.10 | |
| C. semifascialis and C. diminutiva | 7.63 | 9.31 | 8.66 | |
| C. semifascialis and C. haemactalis | 9.01 | 9.47 | 9.18 | |
| C. semifascialis and C. pandamalis | 5.50 | 7.02 | 6.47 | |
| C. tharsalea and C. ersealis | 11.15 | 11.60 | 11.38 | |
| C. tharsalea and C. evaxalis | 11.60 | 12.06 | 11.82 | |
| C. tharsalea and C. diminutiva | 11.76 | 12.06 | 11.95 | |
| C. tharsalea and C. haemactalis | 10.99 | 11.60 | 11.34 | |
| C. tharsalea and C. pandamalis | 11.60 | 12.06 | 11.83 | |
| C. ersealis and C. evaxalis | 9.16 | 9.47 | 9.29 | |
| C. ersealis and C. diminutiva | 2.60 | 3.05 | 2.83 | |
| C. ersealis and C. haemactalis | 8.70 | 8.86 | 8.78 | |
| C. ersealis and C. pandamalis | 7.94 | 8.24 | 8.09 | |
| C. evaxalis and C. diminutiva | 10.23 | 10.69 | 10.53 | |
| C. evaxalis and C. haemactalis | 9.31 | 9.62 | 9.47 | |
| C. evaxalis and C. pandamalis | 9.47 | 9.62 | 9.52 | |
| C. diminutiva and C. haemactalis | - | - | 9.62 | |
| C. diminutiva and C. pandamalis | - | - | 8.40 | |
| C. haemactalis and C. pandamalis | - | - | 10.23 | |
| Within Conogethes divergence | 0.15 | 12.06 | 5.77 | |
| Taxon | No. Haplotype | Min. | Max. | Aver. |
|---|---|---|---|---|
| Intraspecific divergence of C. pinicolalis (inicluding PI 1) | 48 | 0.13 | 5.33 | 0.87 |
| Intraspecific divergence of C. pinicolalis (excluding PI 1) | 42 | 0.13 | 5.33 | 0.86 |
| Intraspecific divergence of C. pinicolalis (only PI 1) | 6 | 0.64 | 1.27 | 0.93 |
| Intraspecific divergence of C. punctiferalis (including PU 2) | 62 | 0.13 | 1.52 | 0.66 |
| Intraspecific divergence of C. punctiferalis (excluding PU 2) | 55 | 0.13 | 1.52 | 0.63 |
| Intraspecific divergence of C. punctiferalis (only PU 2) | 7 | 0.51 | 1.14 | 0.91 |
| C. punctiferalis and C. pinicolalis (including untranslated haplotypes) | 1.02 | 5.96 | 2.11 | |
| C. punctiferalis and C. pinicolalis (excluding untranslated haplotypes) | 1.02 | 5.96 | 2.10 | |
| C. punctiferalis and C. pinicolalis (only untranslated haplotypes) | 1.90 | 2.54 | 2.25 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, N.-R.; Kim, M.-J.; Kim, S.-S.; Choi, S.-W.; Kim, I.-S. Morphological, Ecological, and Molecular Divergence of Conogethes pinicolalis from C. punctiferalis (Lepidoptera: Crambidae). Insects 2021, 12, 455. https://doi.org/10.3390/insects12050455
Jeong N-R, Kim M-J, Kim S-S, Choi S-W, Kim I-S. Morphological, Ecological, and Molecular Divergence of Conogethes pinicolalis from C. punctiferalis (Lepidoptera: Crambidae). Insects. 2021; 12(5):455. https://doi.org/10.3390/insects12050455
Chicago/Turabian StyleJeong, Na-Ra, Min-Jee Kim, Sung-Soo Kim, Sei-Woong Choi, and Ik-Soo Kim. 2021. "Morphological, Ecological, and Molecular Divergence of Conogethes pinicolalis from C. punctiferalis (Lepidoptera: Crambidae)" Insects 12, no. 5: 455. https://doi.org/10.3390/insects12050455
APA StyleJeong, N. -R., Kim, M. -J., Kim, S. -S., Choi, S. -W., & Kim, I. -S. (2021). Morphological, Ecological, and Molecular Divergence of Conogethes pinicolalis from C. punctiferalis (Lepidoptera: Crambidae). Insects, 12(5), 455. https://doi.org/10.3390/insects12050455