
Effect of Cereal α-Amylase/Trypsin Inhibitors on Developmental Characteristics and Abundance of Digestive Enzymes of Mealworm Larvae (Tenebrio molitor L.)

 ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
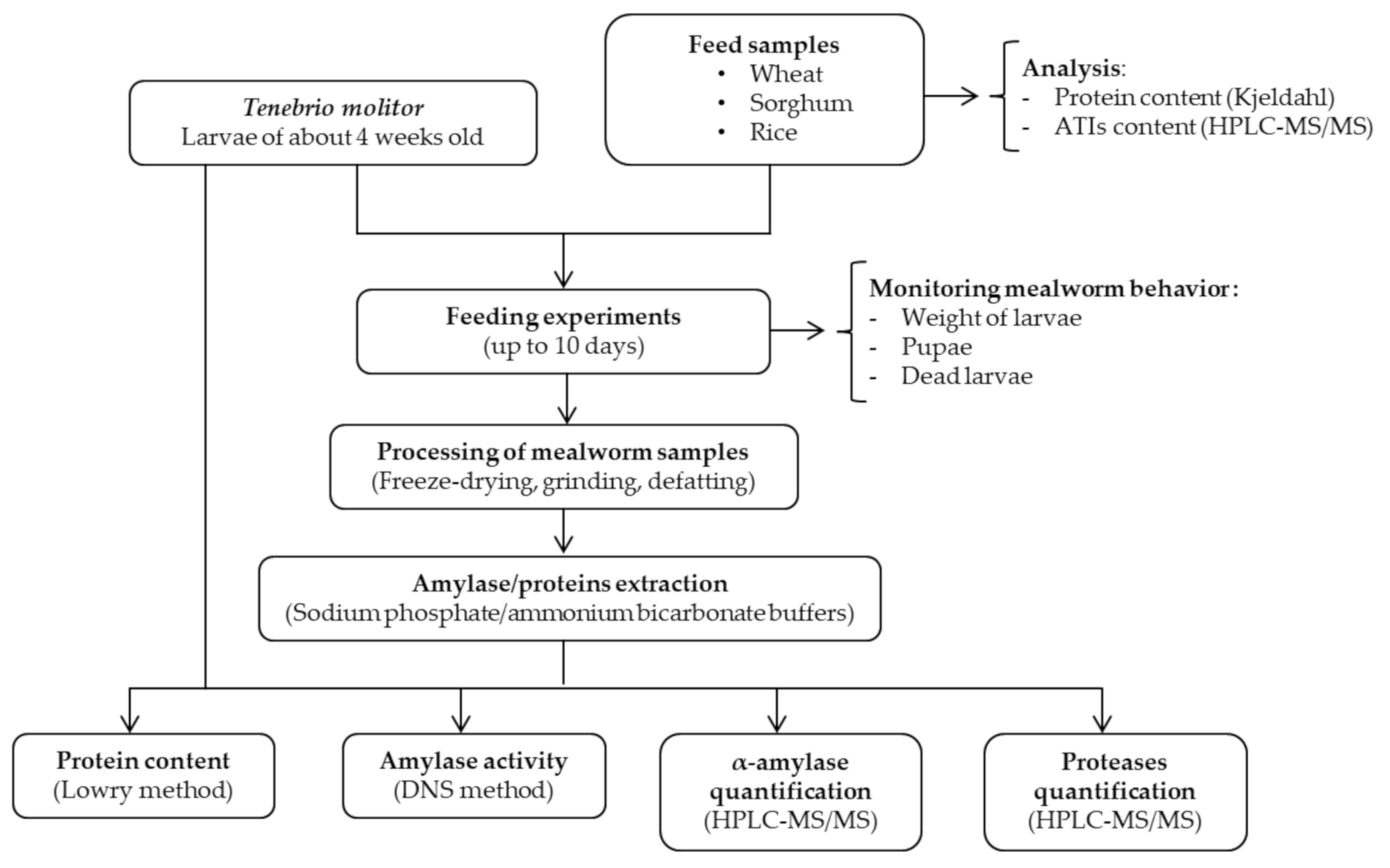
2. Materials and Methods
2.1. Biological Materials and Chemicals
2.2. Design and Implementation of the Feeding Experiments
2.2.1. Feeding Experiments
2.2.2. Sample Preparation for Amylase Activity
2.3. Quantification of α-Amylase and Proteases Using Tandem Mass Spectrometry
2.3.1. Sample Preparation for Mass Spectrometry
2.3.2. LC–ESI–MS/MS Method Development
2.4. Statistic Analysis
3. Results
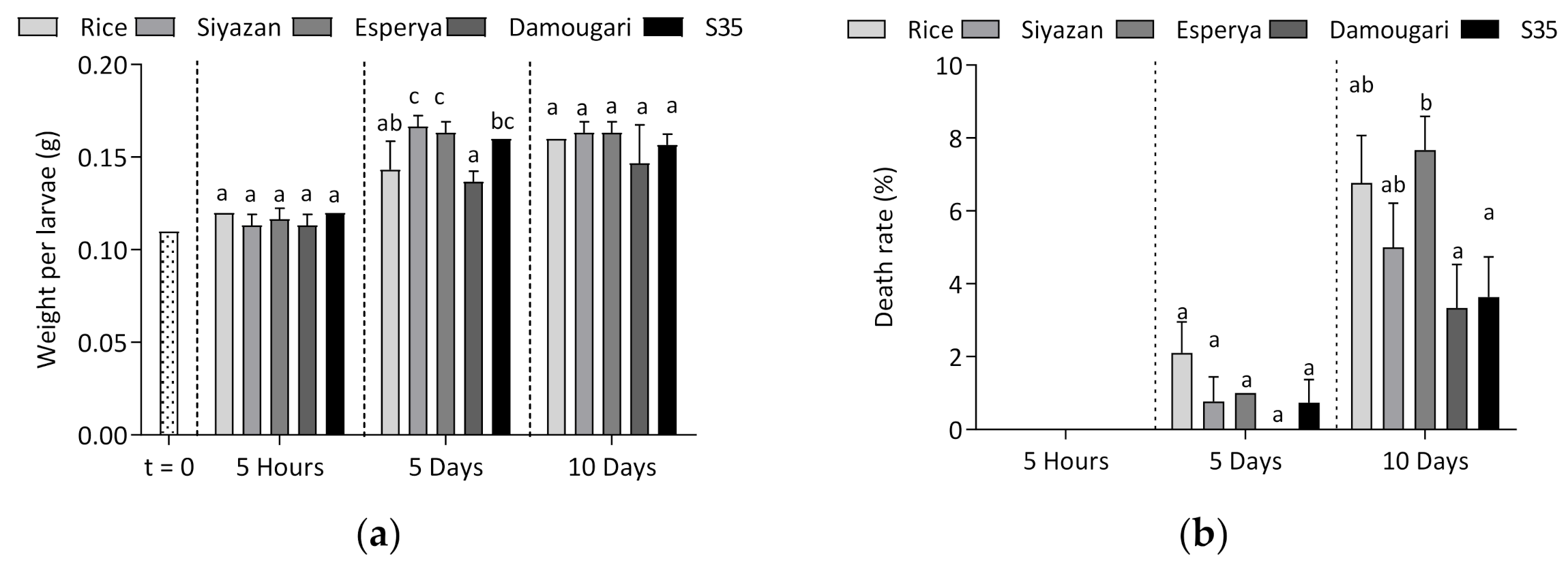
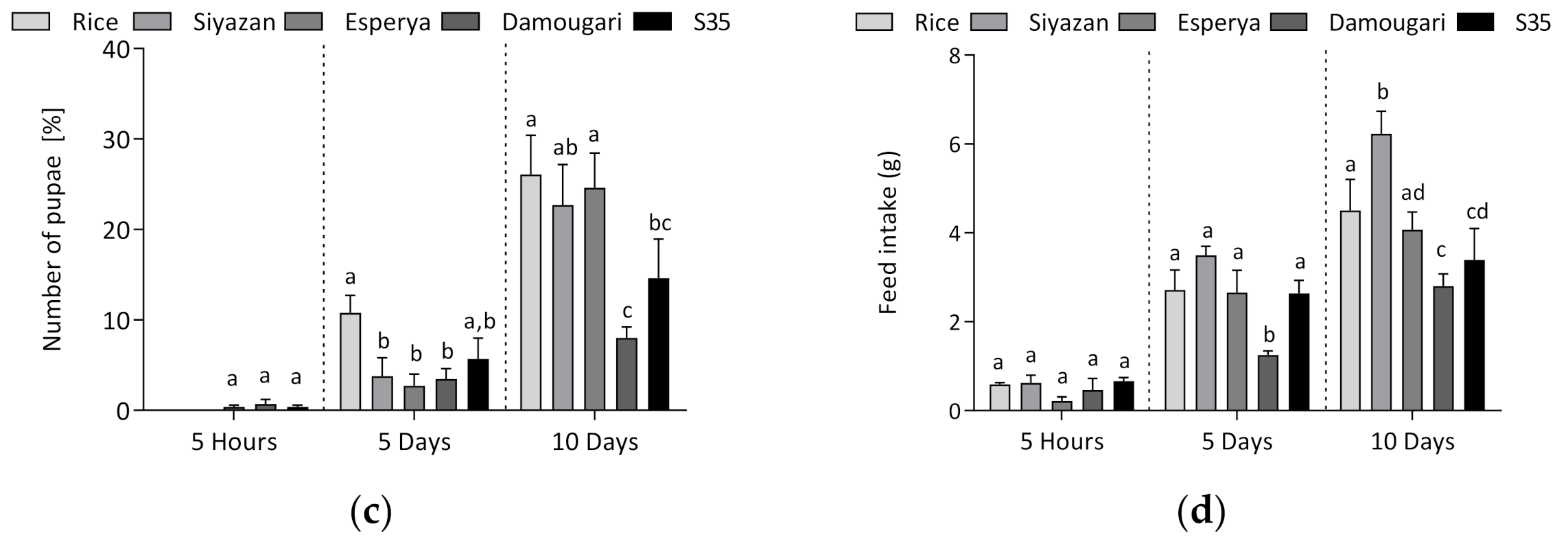
3.1. Developmental Characteristics of Tenebrio molitor Larvae during the Feeding Experiments
3.2. Effect of Feeding Cereals on Larvae Protein Content and Amylase Activity
3.2.1. Protein Content
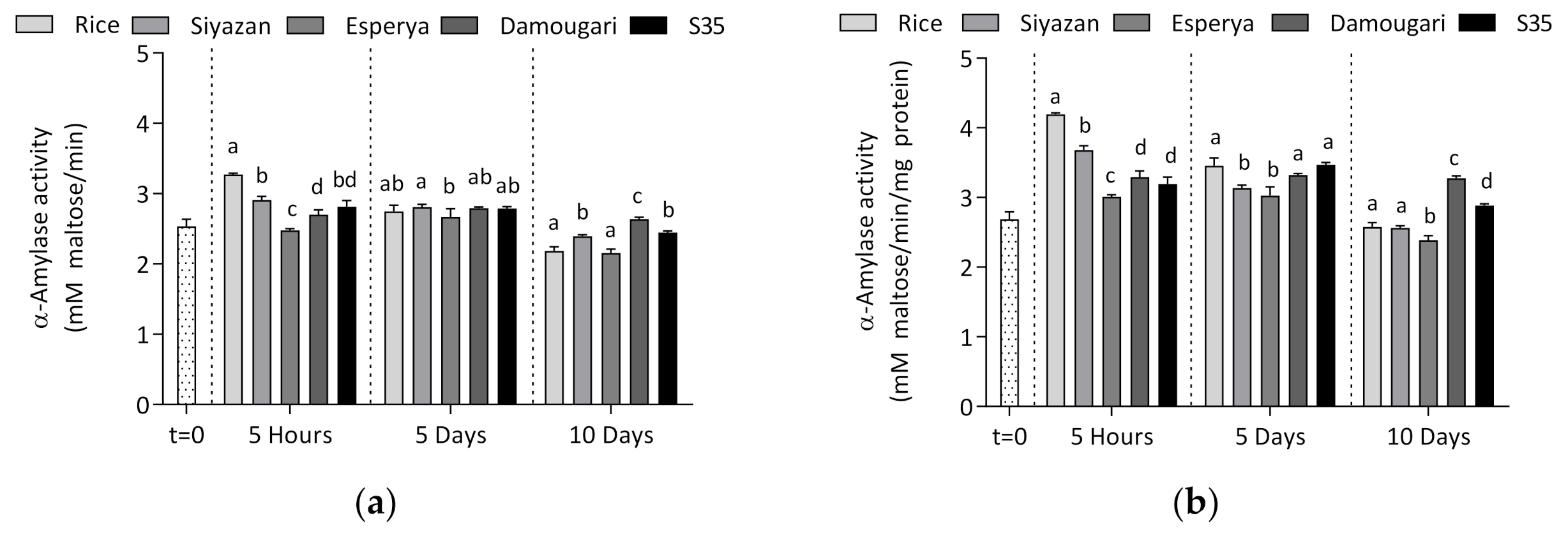
3.2.2. Effect of Feeding with Cereals on α-Amylase Activity
3.3. Determination of the Relative α-Amylase Content in T. molitor
3.3.1. Method Development and Validation
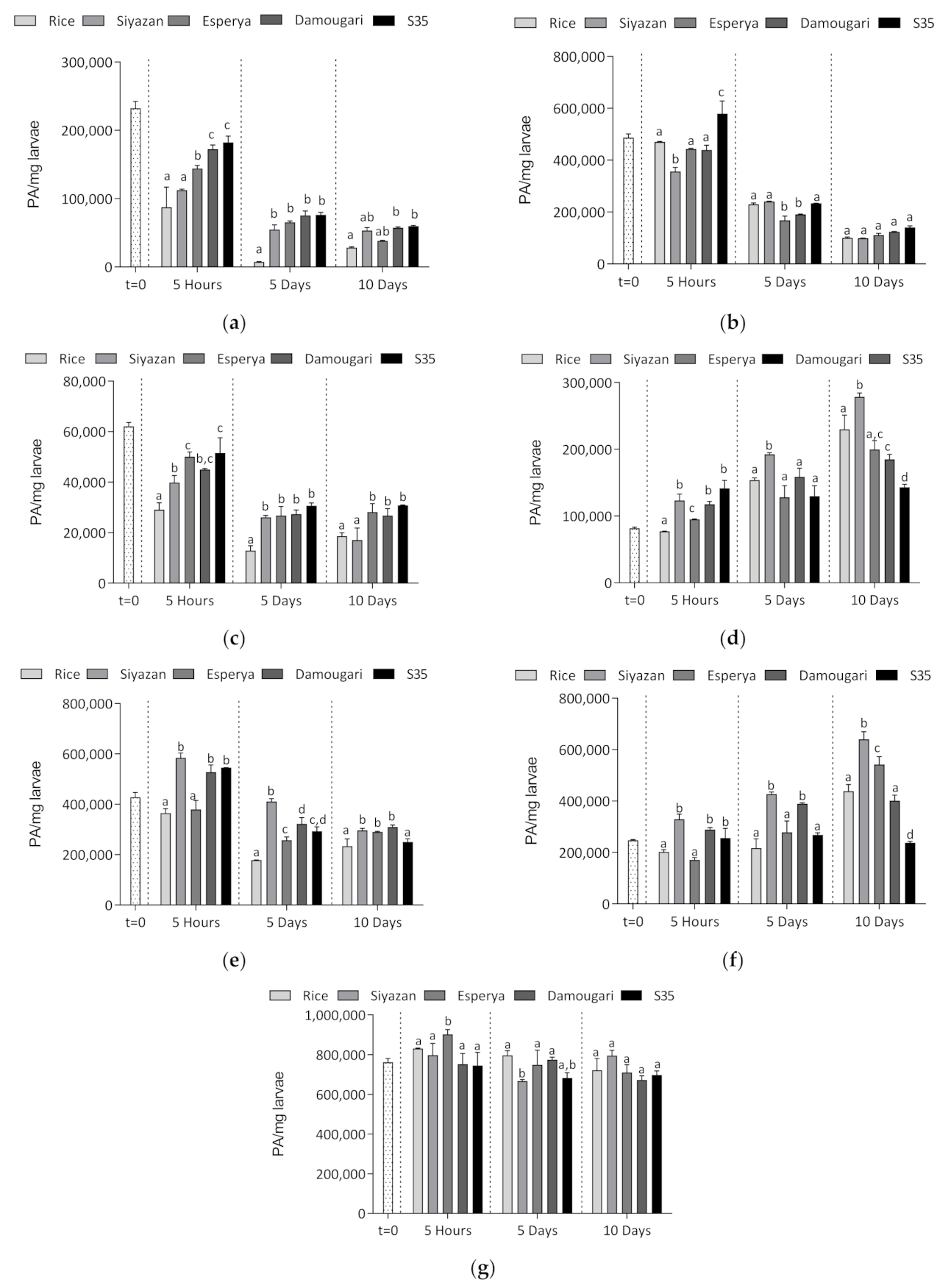
3.3.2. Evaluation of Relative Abundance of Larval α-Amylase and Selected Proteases
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Eriksson, T.; Andere, A.A.; Kelstrup, H.; Emery, V.J.; Picard, C.J. The yellow mealworm (Tenebrio molitor) genome: A resource for the emerging insects as food and feed industry. J. Insects Food Feed 2020, 6, 445–455. [Google Scholar] [CrossRef]
- Bin Park, J.; Choi, W.H.; Kim, S.H.; Jin, H.J.; Han, Y.S.; Lee, Y.S.; Kim, N.J. Developmental characteristics of Tenebrio molitor larvae (Coleoptera: Tenebrionidae) in different instars. Int. J. Ind. Entomol. 2014, 28, 5–9. [Google Scholar] [CrossRef] [Green Version]
- Ghaly, A.E.; Alkoaik, F.N. The Yellow Mealworm as a Novel Source of Protein. Am. J. Agric. Biol. Sci. 2009, 4, 319–331. [Google Scholar] [CrossRef] [Green Version]
- Siemianowska, E.; Kosewska, A.; Aljewicz, M.; Skibniewska, K.A.; Polak-Juszczak, L.; Jarocki, A.; Jedras, M. Larvae of mealworm (Tenebrio molitor L.) as European novel food. Agric. Sci. 2013, 4, 287–291. [Google Scholar] [CrossRef] [Green Version]
- Costa, S.; Pedro, S.; Lourenço, H.; Batista, I.; Teixeira, B.; Bandarra, N.M.; Murta, D.; Nunes, R.; Pires, C. Evaluation of Tenebrio molitor larvae as an alternative food source. NFS J. 2020, 21, 57–64. [Google Scholar] [CrossRef]
- Liu, C.; Masri, J.; Perez, V.; Maya, C.; Zhao, J. Growth Performance and Nutrient Composition of Mealworms (Tenebrio Molitor) Fed on Fresh Plant Materials-Supplemented Diets. Foods 2020, 9, 151. [Google Scholar] [CrossRef] [Green Version]
- Jajić, I.; Popović, A.; Urošević, M.; Krstović, S.; Petrović, M.; Guljaš, D. Chemical Composition of Mealworm Larvae (Tenebrio molitor) Reared in Serbia. Contemp. Agric. 2019, 68, 23–27. [Google Scholar] [CrossRef] [Green Version]
- Shewry, P.R.; Halford, N.G.; Belton, P.S.; Tatham, A.S. The structure and properties of gluten: An elastic protein from wheat grain. Philos. Trans. R. Soc. B Biol. Sci. 2002, 357, 133–142. [Google Scholar] [CrossRef]
- Shewry, P.R.; Halford, N.G. Cereal seed storage proteins: Structures, properties and role in grain utilization. J. Exp. Bot. 2002, 53, 947–958. [Google Scholar] [CrossRef] [Green Version]
- Shewry, P.R.; Tatham, A.S. The prolamin storage proteins of cereal seeds: Structure and evolution. Biochem. J. 1990, 267, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Goptar, I.; Semashko, T.; Danilenko, S.; Lysogorskaya, E.; Oksenoit, E.; Zhuzhikov, D.; Belozersky, M.; Dunaevsky, Y.; Oppert, B.; Filippova, I.; et al. Cysteine digestive peptidases function as post-glutamine cleaving enzymes in tenebrionid stored-product pests. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2012, 161, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Vinokurov, K.; Elpidina, E.; Oppert, B.; Prabhakar, S.; Zhuzhikov, D.; Dunaevsky, Y.; Belozersky, M. Diversity of digestive proteinases in Tenebrio molitor (Coleoptera: Tenebrionidae) larvae. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2006, 145, 126–137. [Google Scholar] [CrossRef]
- Vinokurov, K.; Elpidina, E.; Oppert, B.; Prabhakar, S.; Zhuzhikov, D.; Dunaevsky, Y.; Belozersky, M. Fractionation of digestive proteinases from Tenebrio molitor (Coleoptera: Tenebrionidae) larvae and role in protein digestion. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2006, 145, 138–146. [Google Scholar] [CrossRef] [PubMed]
- Martynov, A.G.; Elpidina, E.N.; Perkin, L.; Oppert, B. Functional analysis of C1 family cysteine peptidases in the larval gut of Тenebrio molitor and Tribolium castaneum. BMC Genom. 2015, 16, 75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prabhakar, S.; Chen, M.-S.; Elpidina, E.N.; Vinokurov, K.S.; Smith, C.M.; Marshall, J.; Oppert, B. Sequence analysis and molecular characterization of larval midgut cDNA transcripts encoding peptidases from the yellow mealworm, Tenebrio molitor L. Insect Mol. Biol. 2007, 16, 455–468. [Google Scholar] [CrossRef]
- Terra, W.R.; Cristofoletti, P.T. Midgut proteinases in three divergent species of Coleoptera. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 1996, 113, 725–730. [Google Scholar] [CrossRef]
- Elpidina, E.N.; Goptar, I.A. Digestive peptidases in Tenebrio molitor and possibility of use to treat celiac disease. Entomol. Res. 2007, 37, 139–147. [Google Scholar] [CrossRef]
- Hangay, G.; Gayubo, S.F.; Hoy, M.A.; Goula, M.; Sanborn, A.; Morrill, W.L.; Gäde, G.; Marco, H.G.; Kabissa, J.C.B.; Ellis, J.; et al. Alimentary Canal and Digestion. Encycl. Entomol. 2008, 111–118. [Google Scholar] [CrossRef]
- Tsybina, T.A.; Dunaevsky, Y.E.; Belozersky, M.A.; Zhuzhikov, D.P.; Oppert, B.; Elpidina, E.N. Digestive proteinases of yellow mealworm (Tenebrio molitor) larvae: Purification and characterization of a trypsin-like proteinase. Biochemistry 2005, 70, 300–305. [Google Scholar] [CrossRef]
- Elpidina, E.; Tsybina, T.; Dunaevsky, Y.; Belozersky, M.; Zhuzhikov, D.; Oppert, B. A chymotrypsin-like proteinase from the midgut of larvae. Biochimie 2005, 87, 771–779. [Google Scholar] [CrossRef]
- Strobl, S.; Maskos, K.; Wiegand, G.; Huber, R.; Gomis-Rüth, F.X.; Glockshuber, R. A novel strategy for inhibition of α-amylases: Yellow meal worm α-amylase in complex with the Ragi bifunctional inhibitor at 2.5 å resolution. Structure 1998, 6, 911–921. [Google Scholar] [CrossRef] [Green Version]
- Capinera, J.L.; Hoy, M.A.; Paré, P.W.; Farag, M.A.; Trumble, J.T.; Isman, M.B.; Adams, B.J.; Nguyen, K.B.; Panizzi, A.R.; Sánchez, N.E.; et al. (Eds.) Nutrition in Insects. In Encyclopedia of Entomology; Springer: Dordrecht, The Netherlands, 2008; pp. 2646–2654. [Google Scholar]
- Strobl, S.; Maskos, K.; Betz, M.; Wiegand, G.; Huber, R.; Gomis-Rüth, X.; Glockshuber, R. Crystal structure of yellow meal worm α-amylase at 1.64 Å resolution. J. Mol. Biol. 1998, 278, 617–628. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, M.C.M.; Grossi de Sá, M.F.; Chrispeels, M.J.; Togawa, R.C.; Neshich, G. Analysis of structural and physico-chemical parameters involved in the specificity of binding between α-amylases and their inhibitors. Protein Eng. Des. Sel. 2000, 13, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Strobl, S.; Gomis-Rüth, F.-X.; Maskos, K.; Frank, G.; Huber, R.; Glockshuber, R. The α-amylase from the yellow meal worm: Complete primary structure, crystallization and preliminary X-ray analysis. FEBS Lett. 1997, 409, 109–114. [Google Scholar] [CrossRef] [Green Version]
- Lazarević, J.; Janković-Tomanić, M. Dietary and phylogenetic correlates of digestive trypsin activity in insect pests. Entomol. Exp. Appl. 2015, 157, 123–151. [Google Scholar] [CrossRef]
- Spit, J.; Zels, S.; Dillen, S.; Holtof, M.; Wynant, N.; Broeck, J.V. Effects of different dietary conditions on the expression of trypsin- and chymotrypsin-like protease genes in the digestive system of the migratory locust, Locusta migratoria. Insect Biochem. Mol. Biol. 2014, 48, 100–109. [Google Scholar] [CrossRef]
- Sagu, S.T.; Huschek, G.; Bönick, J.; Homann, T.; Rawel, H.M. A New Approach of Extraction of α-Amylase/trypsin Inhibitors from Wheat (Triticum aestivum L.), Based on Optimization Using Plackett–Burman and Box–Behnken Designs. Molecules 2019, 24, 3589. [Google Scholar] [CrossRef] [Green Version]
- Priya, S.; Kumar, S.; Kaur, N.; Gupta, A. Specificity of α-amylase and trypsin inhibitor proteins in wheat against insect pests. N.Z.J. Crop. Hortic. Sci. 2013, 41, 49–56. [Google Scholar] [CrossRef] [Green Version]
- Sagu, S.T.; Zimmermann, L.; Landgräber, E.; Homann, T.; Huschek, G.; Özpinar, H.; Schweigert, F.J.; Rawel, H.M. Comprehensive Characterization and Relative Quantification of α-Amylase/Trypsin Inhibitors from Wheat Cultivars by Targeted HPLC-MS/MS. Foods 2020, 9, 1448. [Google Scholar] [CrossRef] [PubMed]
- Altenbach, S.B.; Vensel, W.H.; Dupont, F.M. The spectrum of low molecular weight alpha-amylase/protease inhibitor genes expressed in the US bread wheat cultivar Butte 86. BMC Res. Notes 2011, 4, 242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iulek, J.; Franco, O.L.; Silva, M.; Slivinski, C.T.; Bloch, C.; Rigden, D.J.; de Sá, M.F.G. Purification, biochemical characterisation and partial primary structure of a new α-amylase inhibitor from Secale cereale (rye). Int. J. Biochem. Cell Biol. 2000, 32, 1195–1204. [Google Scholar] [CrossRef]
- Cuccioloni, M.; Mozzicafreddo, M.; Ali, I.; Bonfili, L.; Cecarini, V.; Eleuteri, A.M.; Angeletti, M. Interaction between wheat alpha-amylase/trypsin bi-functional inhibitor and mammalian digestive enzymes: Kinetic, equilibrium and structural characterization of binding. Food Chem. 2016, 213, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Kuwar, S.S.; Pauchet, Y.; Vogel, H.; Heckel, D.G. Adaptive regulation of digestive serine proteases in the larval midgut of Helicoverpa armigera in response to a plant protease inhibitor. Insect Biochem. Mol. Biol. 2015, 59, 18–29. [Google Scholar] [CrossRef]
- Silva, C.; Terra, W.; De Sá, M.; Samuels, R.; Isejima, E.; Bifano, T.; Almeida, J. Induction of digestive α-amylases in larvae of Zabrotes subfasciatus (Coleoptera: Bruchidae) in response to ingestion of common bean α-amylase inhibitor 1. J. Insect Physiol. 2001, 47, 1283–1290. [Google Scholar] [CrossRef]
- Oppert, B.; Morgan, T.; Hartzer, K.; Kramer, K. Compensatory proteolytic responses to dietary proteinase inhibitors in the red flour beetle, Tribolium castaneum (Coleoptera: Tenebrionidae). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2005, 140, 53–58. [Google Scholar] [CrossRef]
- Feng, G.H.; Richardson, M.; Chen, M.S.; Kramer, K.J.; Morgan, T.D.; Reeck, G.R. α-amylase inhibitors from wheat: Amino acid sequences and patterns of inhibition of insect and human α-amylases. Insect Biochem. Mol. Biol. 1996, 26, 419–426. [Google Scholar] [CrossRef]
- Buonocore, V.; Gramenzi, F.; Pace, W.; Petrucci, T.; Poerio, E.; Silano, V. Interaction of wheat monomeric and dimeric protein inhibitors with α-amylase from yellow mealworm (Tenebrio molitor L. larva). Biochem. J. 1980, 187, 637–645. [Google Scholar] [CrossRef] [PubMed]
- Sagu, S.T.; Landgräber, E.; Rackiewicz, M.; Huschek, G.; Rawel, H. Relative Abundance of Alpha-Amylase/Trypsin Inhibitors in Selected Sorghum Cultivars. Molecules 2020, 25, 5982. [Google Scholar] [CrossRef]
- Tchewonpi, S.S.; Nso, E.J.; Homann, T.; Kapseu, C.; Rawel, H.M. Extraction and purification of beta-amylase from stems of Abrus precatorius by three phase partitioning. Food Chem. 2015, 183, 144–153. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- van Broekhoven, S.; Oonincx, D.G.; van Huis, A.; van Loon, J.J. Growth performance and feed conversion efficiency of three edible mealworm species (Coleoptera: Tenebrionidae) on diets composed of organic by-products. J. Insect Physiol. 2015, 73, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Franco, O.L.; Melo, F.R.; Mendes, P.A.; Paes, N.S.; Yokoyama, M.; Coutinho, M.V.; Bloch, C.; Grossi-De-Sa, M.F. Characterization of Two Acanthoscelides obtectus α-Amylases and Their Inactivation by Wheat Inhibitors. J. Agric. Food Chem. 2005, 53, 1585–1590. [Google Scholar] [CrossRef]
- Morales-Ramos, J.A.; Rojas, M.G.; Shelby, K.S.; Coudron, T.A. Nutritional Value of Pupae Versus Larvae of Tenebrio molitor (Coleoptera: Tenebrionidae) as Food for Rearing Podisus maculiventris (Heteroptera: Pentatomidae). J. Econ. Entomol. 2015, 109, 564–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu-Salzman, K.; Zeng, R. Insect Response to Plant Defensive Protease Inhibitors. Annu. Rev. Entomol. 2015, 60, 233–252. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Cultivar | Protein Content * (g/100 g) | ATI Content ** | |
|---|---|---|---|---|
| (PA/mg Flour) | (PA/µg Protein) | |||
| Wheat | Siyazan | 15.6 | 79,431 ± 1218 | 931 ± 14 |
| Esperya | 13.9 | 1,460,159 ± 95,183 | 25,231 ± 1645 | |
| Sorghum | Damougari | 8.5 | 14,903 ± 223 | 935 ± 14 |
| S35 | 7.9 | 147,671 ± 2509 | 11,017 ± 187 | |
| Rice | n.a. | 7.9 | 473,317 ± 5349 | 17,174 ± 193 |
| Feeding Samples | ||||||||
|---|---|---|---|---|---|---|---|---|
| I.L. | Rice | Siyazan | Esperya | Damougari | S35 | |||
| Protein content (mg/mL) | Na3PO4 buffer | t = 0 | 9.42 ± 0.19 | - | - | - | - | - |
| 5 Hours | - | 7.80 ± 0.09 a | 7.90 ± 0.16 a,b | 8.22 ± 0.04 b | 8.20 ± 0.22 a,b | 8.80 ± 0.10 c | ||
| 5 Days | - | 7.94 ± 0.18 a | 8.97 ± 0.28 b | 8.82 ± 0.25 b | 8.40 ± 0.23 c | 8.05 ± 0.19 a,c | ||
| 10 Days | - | 8.50 ± 0.08 a | 9.33 ± 0.04 b | 9.02 ± 0.08 b | 8.06 ± 0.07 c | 8.49 ± 0.30 a | ||
| Ambi/urea buffer | t = 0 | 34.93 ± 0.44 | ||||||
| 5 Hours | - | 26.73 ± 0.72 a,b | 27.05 ± 0.16 b | 25.21 ± 0.69 a | 29.45 ± 0.11 c | 26.56 ± 0.71 a,b | ||
| 5 Days | - | 37.47 ± 1.38 a | 32.74 ± 0.21 b | 26.91 ± 1.01 c | 34.75 ± 0.38 d | 33.97 ± 0.75 b,d | ||
| 10 Days | - | 22.39 ± 0.74 a | 19.78 ± 0.16 b | 19.95 ± 0.34 b,c | 30.41 ± 0.21 d | 27.48 ± 0.75 e | ||
| Total Protein (g/100 g) | t = 0 | 53.4 | - | - | - | - | - | |
| 5 Hours | - | 48.6 | 49.8 | 49.7 | 49.9 | 48.7 | ||
| 5 Days | - | 45.1 | 46.0 | 46.8 | 48.1 | 44.8 | ||
| 10 Days | - | 44.0 | 45.7 | 47.5 | 45.6 | 43.9 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sagu, S.T.; Landgräber, E.; Henkel, I.M.; Huschek, G.; Homann, T.; Bußler, S.; Schlüter, O.K.; Rawel, H. Effect of Cereal α-Amylase/Trypsin Inhibitors on Developmental Characteristics and Abundance of Digestive Enzymes of Mealworm Larvae (Tenebrio molitor L.). Insects 2021, 12, 454. https://doi.org/10.3390/insects12050454
Sagu ST, Landgräber E, Henkel IM, Huschek G, Homann T, Bußler S, Schlüter OK, Rawel H. Effect of Cereal α-Amylase/Trypsin Inhibitors on Developmental Characteristics and Abundance of Digestive Enzymes of Mealworm Larvae (Tenebrio molitor L.). Insects. 2021; 12(5):454. https://doi.org/10.3390/insects12050454
Chicago/Turabian StyleSagu, Sorel Tchewonpi, Eva Landgräber, Ina M. Henkel, Gerd Huschek, Thomas Homann, Sara Bußler, Oliver K. Schlüter, and Harshadrai Rawel. 2021. "Effect of Cereal α-Amylase/Trypsin Inhibitors on Developmental Characteristics and Abundance of Digestive Enzymes of Mealworm Larvae (Tenebrio molitor L.)" Insects 12, no. 5: 454. https://doi.org/10.3390/insects12050454
APA StyleSagu, S. T., Landgräber, E., Henkel, I. M., Huschek, G., Homann, T., Bußler, S., Schlüter, O. K., & Rawel, H. (2021). Effect of Cereal α-Amylase/Trypsin Inhibitors on Developmental Characteristics and Abundance of Digestive Enzymes of Mealworm Larvae (Tenebrio molitor L.). Insects, 12(5), 454. https://doi.org/10.3390/insects12050454