
In Vitro Rearing Changes Social Task Performance and Physiology in Honeybees

 , , , and
, , , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. In Vitro Rearing
2.2. Comparison of Nursing Behavior
2.3. Comparison of Foraging Behavior
2.4. Weight, Juvenile Hormone, Triglycerides and Sucrose Responsiveness
2.5. Data Analysis
3. Results
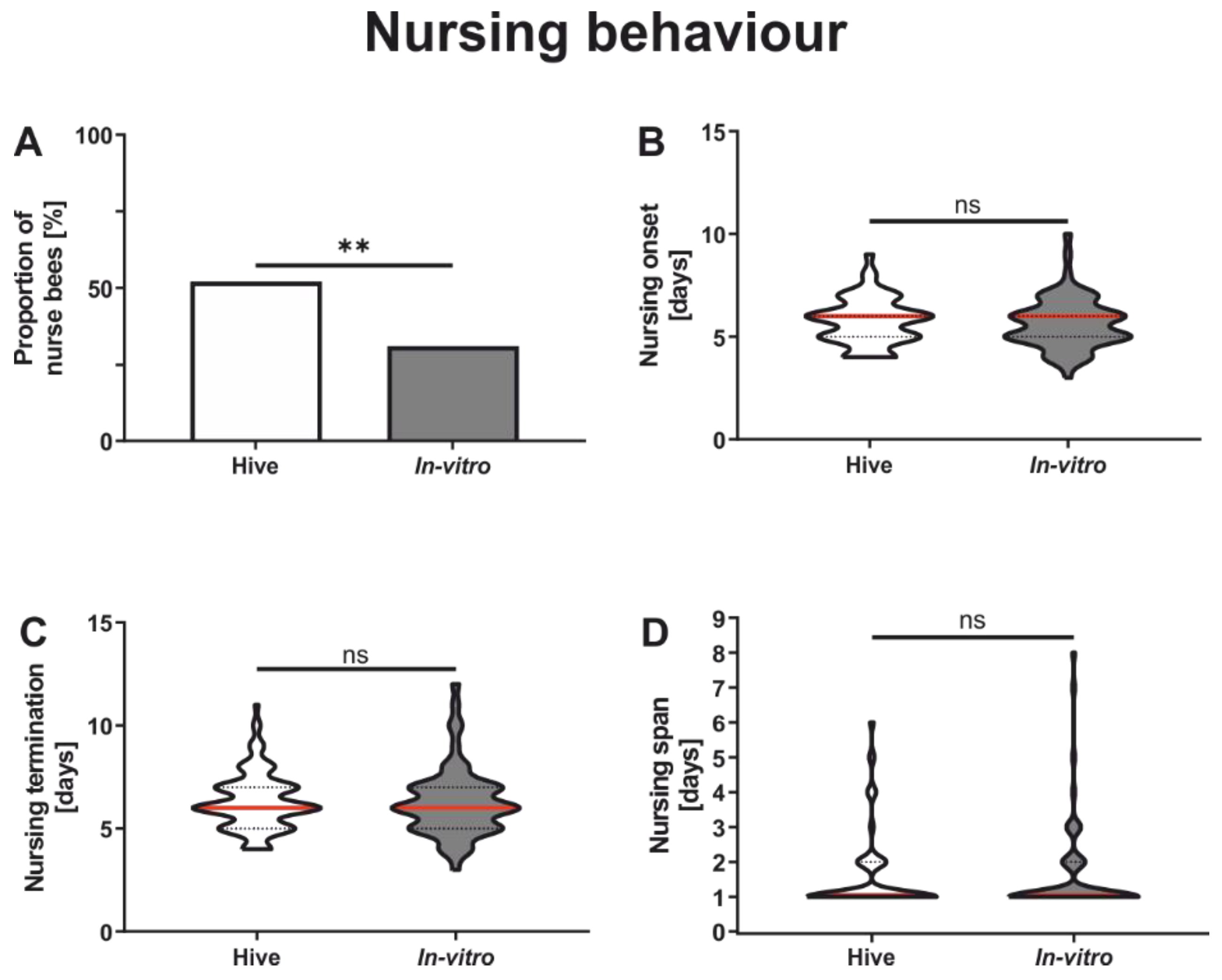
3.1. Comparison of Nursing Behavior
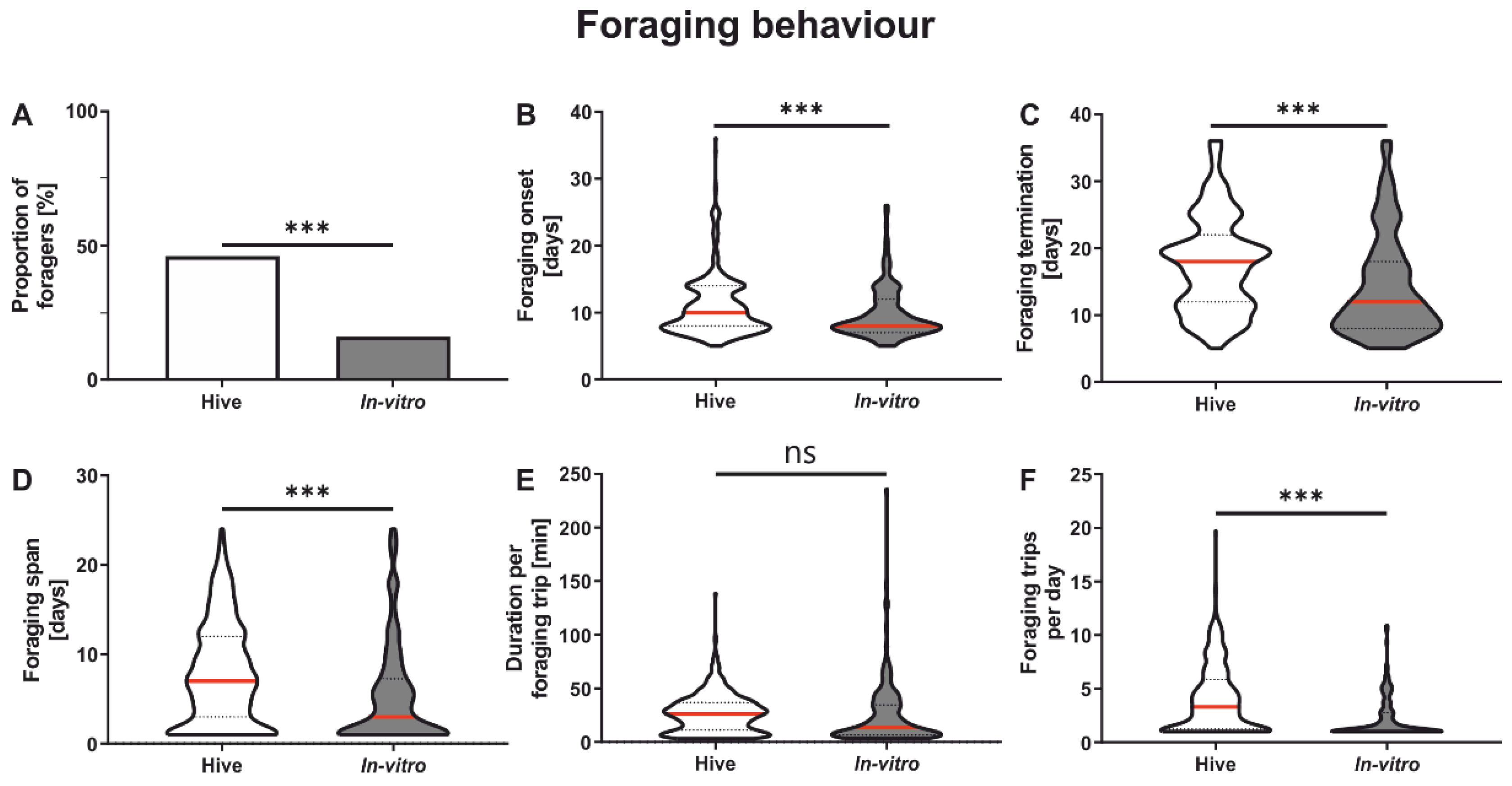
3.2. Foraging Behavior
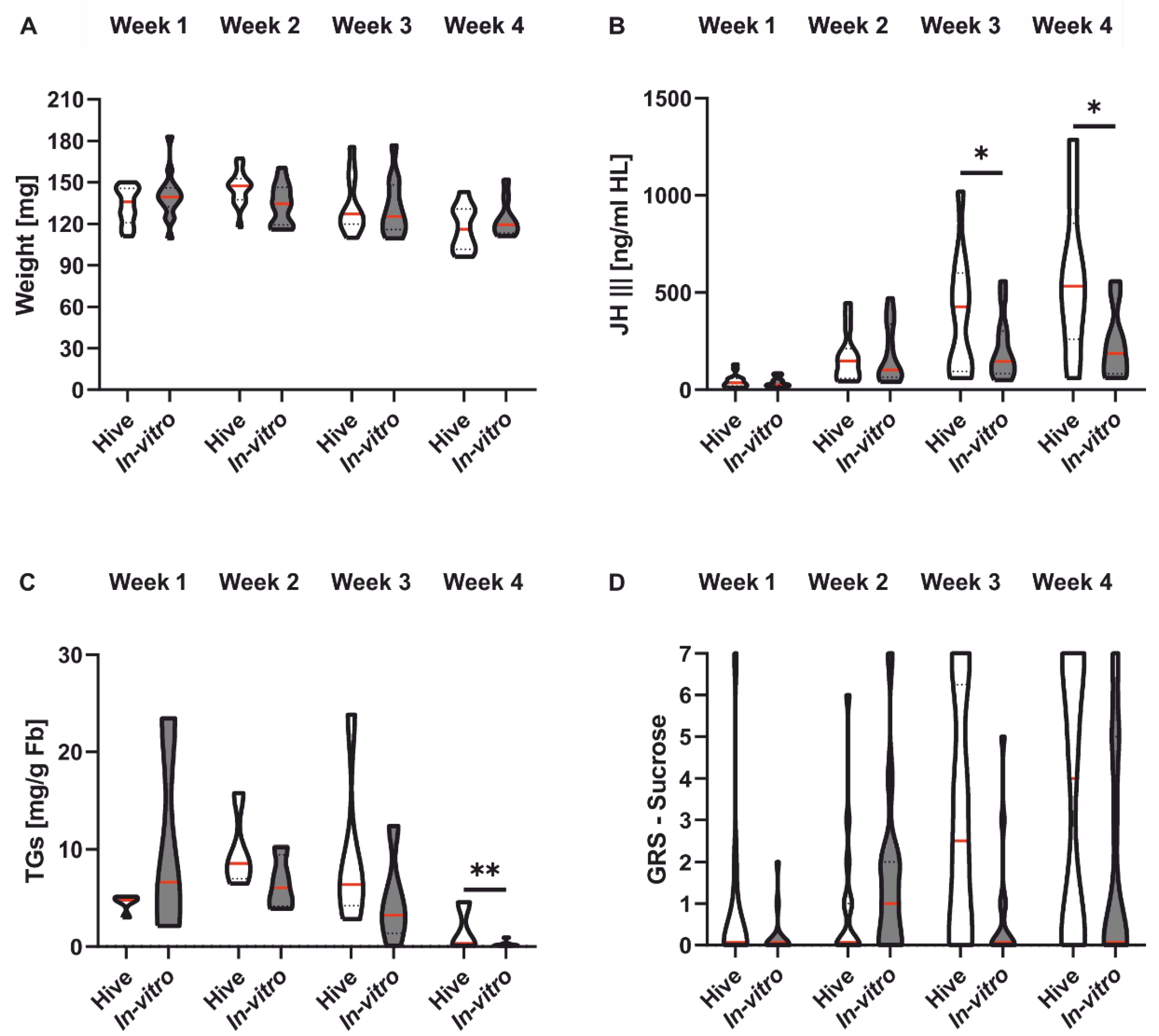
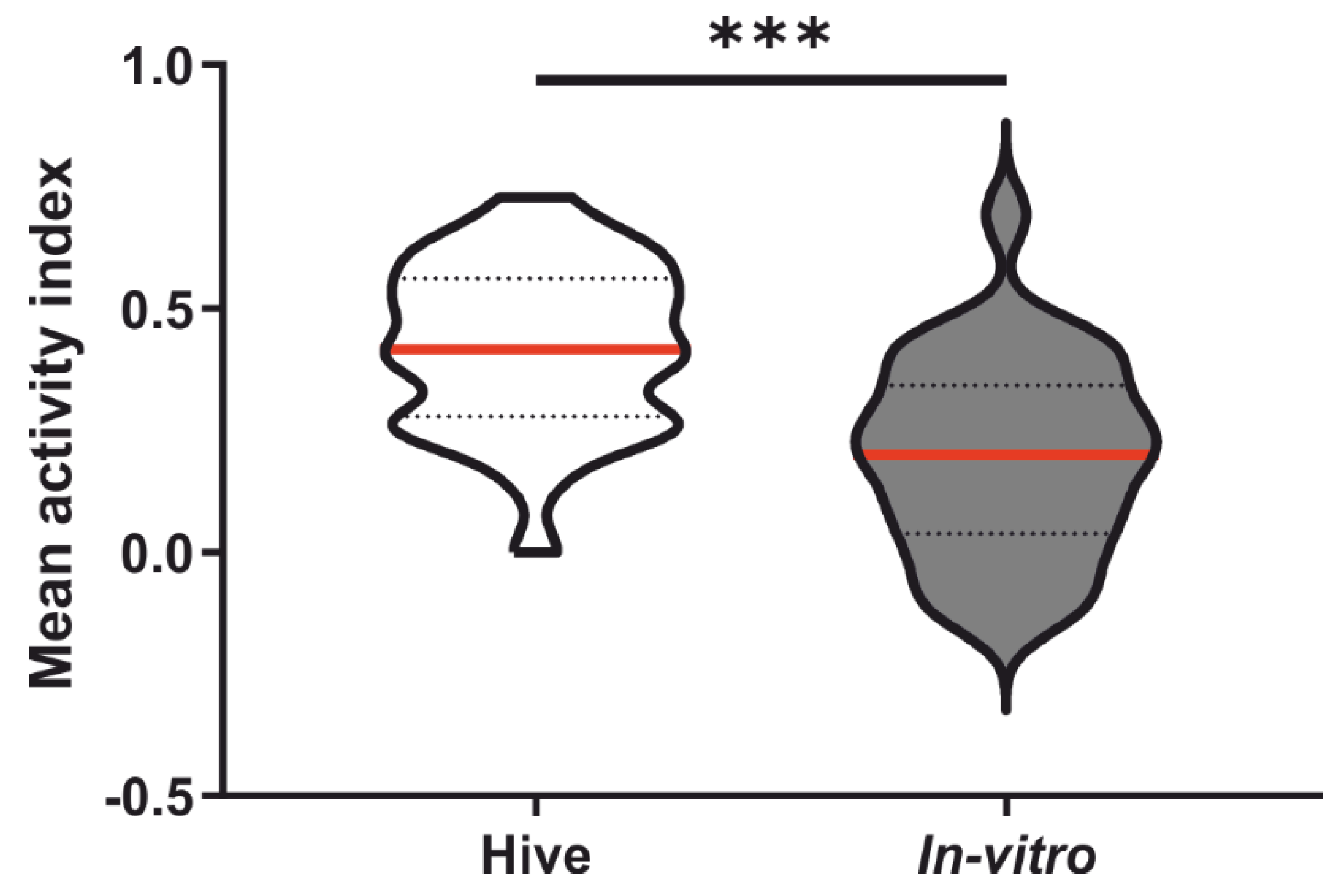
3.3. Honeybee Morphology, Physiology and Sucrose Responsiveness
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Seeley, T.D. The Wisdom of the Hive: The Social Physiology of Honey Bee Colonies; Harvard University Press: Cambridge, MA, USA, 1996; ISBN 978-0-674-95376-5. [Google Scholar]
- Winston, M.L. The Biology of the Honey Bee; Harvard University Press: Cambridge, MA, USA, 1987; ISBN 978-0-674-07409-5. [Google Scholar]
- Huang, Z.-Y.; Robinson, G.E. Regulation of honey bee division of labor by colony age demography. Behav. Ecol. Sociobiol. 1996, 39, 147–158. [Google Scholar] [CrossRef]
- Toth, A.L.; Kantarovich, S.; Meisel, A.F.; Robinson, G.E. Nutritional status influences socially regulated foraging ontogeny in honey bees. J. Exp. Biol. 2005, 208, 4641–4649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amdam, G.V. Social context, stress, and plasticity of aging. Aging Cell 2011, 10, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Bordier, C.; Suchail, S.; Pioz, M.; Devaud, J.M.; Collet, C.; Charreton, M.; Le Conte, Y.; Alaux, C. Stress response in honeybees is associated with changes in task-related physiology and energetic metabolism. J. Insect Physiol. 2017, 98, 47–54. [Google Scholar] [CrossRef]
- Zanni, V.; Değirmenci, L.; Annoscia, D.; Scheiner, R.; Nazzi, F. The reduced brood nursing by mite-infested honey bees depends on their accelerated behavioral maturation. J. Insect Physiol. 2018, 109, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Hendriksma, H.P.; Härtel, S.; Steffan-Dewenter, I. Honey bee risk assessment: New approaches for in vitro larvae rearing and data analyses. Methods Ecol. Evol. 2011, 2, 509–517. [Google Scholar] [CrossRef]
- Steijven, K.; Spaethe, J.; Steffan-Dewenter, I.; Härtel, S. Learning performance and brain structure of artificially-reared honey bees fed with different quantities of food. PeerJ 2017, 5, e3858. [Google Scholar] [CrossRef] [Green Version]
- Foley, K.; Fazio, G.; Jensen, A.B.; Hughes, W.O.H. Nutritional limitation and resistance to opportunistic Aspergillus parasites in honey bee larvae. J. Invertebr. Pathol. 2012, 111, 68–73. [Google Scholar] [CrossRef]
- Peng, C.Y.-S.; Mussen, E.; Fong, A.; Cheng, P.; Wong, G.; Montague, M.A. Laboratory and Field Studies on the Effects of the Antibiotic Tylosin on Honey Bee Apis mellifera L. (Hymenoptera: Apidae) Development and Prevention of American Foulbrood Disease. J. Invertebr. Pathol. 1996, 67, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Charpentier, G.; Vidau, C.; Ferdy, J.B.; Tabart, J.; Vetillard, A. Lethal and sub-lethal effects of thymol on honeybee (Apis mellifera) larvae reared in vitro. Pest Manag. Sci. 2014, 70, 140–147. [Google Scholar] [CrossRef]
- Mortensen, A.N.; Ellis, J.D. The effects of artificial rearing environment on the behavior of adult honey bees, Apis mellifera L. Behav. Ecol. Sociobiol. 2018, 72, 92. [Google Scholar] [CrossRef]
- Schmehl, D.R.; Tomé, H.V.V.; Mortensen, A.N.; Martins, G.F.; Ellis, J.D. Protocol for the in vitro rearing of honey bee (Apis mellifera L.) workers. J. Apic. Res. 2016, 55, 113–129. [Google Scholar] [CrossRef] [Green Version]
- Aupinel, P.; Fortini, D.; Dufour, H.; Taśei, J.N.; Michaud, B.; Odoux, J.-F.; Pham-Delègue, M.-H. Improvement of artificial feeding in a standard in vitro method for rearing Apis mellifera larvae. Bull. Insectology 2005, 58, 107. [Google Scholar]
- Aupinel, P.; Fortini, D.; Michaud, B.; Marolleau, F.; Tasei, J.-N.; Odoux, J.-F. Toxicity of dimethoate and fenoxycarb to honey bee brood (Apis mellifera), using a newin vitro standardized feeding method. Pest Manag. Sci. 2007, 63, 1090–1094. [Google Scholar] [CrossRef]
- Kaftanoglu, O.; Linksvayer, T.A.; Page, R.E. Rearing Honey Bees, Apis mellifera, in vitro 1: Effects of Sugar Concentrations on Survival and Development. J. Insect Sci. 2011, 11, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aupinel, P.; Fortini, D.; Michaud, B.; Medrzycki, P.; Padovani, E.; Przygoda, D.; Maus, C.; Charriere, J.-D.; Kilchenmann, V.; Riessberger-Galle, U.; et al. Honey bee brood ring-test: Method for testing pesticide toxicity on honeybee brood in laboratory conditions. Julius-Kühn-Archiv 2009, 423, 96–102. [Google Scholar]
- Schmickl, T.; Crailsheim, K. How honeybees (Apis mellifera L.) change their broodcare behaviour in response to non-foraging conditions and poor pollen conditions. Behav. Ecol. Sociobiol. 2002, 51, 415–425. [Google Scholar] [CrossRef]
- Technau, G.M. Fiber number in the mushroom bodies of adult drosophila melanogaster depends on age, sex and experience. J. Neurogenet. 1984, 1, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Campbell, J.B.; Kaftanoglu, O.; Page, R.E.; Amdam, G.V.; Harrison, J.F. Larval starvation improves metabolic response to adult starvation in honey bees (Apis mellifera L.). J. Exp. Biol. 2016, 219, 960–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scofield, H.N.; Mattila, H.R. Honey bee workers that are pollen stressed as larvae become poor foragers and waggle dancers as adults. PLoS ONE 2015, 10, e0121731. [Google Scholar] [CrossRef] [PubMed]
- Hoover, S.E.R.; Higo, H.A.; Winston, M.L. Worker honey bee ovary development: Seasonal variation and the influence of larval and adult nutrition. J. Comp. Physiol. B Biochem. Syst. Environ. Physiol. 2006, 176, 55–63. [Google Scholar] [CrossRef]
- Brodschneider, R.; Riessberger-Gallé, U.; Crailsheim, K. Flight performance of artificially reared honeybees (Apis mellifera). Apidologie 2009, 40, 441–449. [Google Scholar] [CrossRef] [Green Version]
- Huang, Z.Y.; Robinson, G.E.; Borst, D.W. Physiological correlates of division of labor among similarly aged honey bees. J. Comp. Physiol. A 1994, 174, 731–739. [Google Scholar] [CrossRef]
- Huang, Z.Y.; Robinson, G.E. Seasonal changes in juvenile hormone titers and rates of biosynthesis in honey bees. J. Comp. Physiol. B 1995, 165, 18–28. [Google Scholar] [CrossRef] [PubMed]
- Ament, S.A.; Wang, Y.; Robinson, G.E. Nutritional regulation of division of labor in honey bees: Toward a systems biology perspective. Wiley Interdiscip. Rev. Syst. Biol. Med. 2010, 2, 566–576. [Google Scholar] [CrossRef] [PubMed]
- Amdam, G.V.; Omholt, S.W. The hive bee to forager transition in honeybee colonies: The double repressor hypothesis. J. Theor. Biol. 2003, 223, 451–464. [Google Scholar] [CrossRef]
- Amdam, G.V.; Norberg, K.; Page, R.E.; Erber, J.; Scheiner, R. Downregulation of vitellogenin gene activity increases the gustatory responsiveness of honey bee workers (Apis mellifera). Behav. Brain Res. 2006, 169, 201–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scheiner, R.; Entler, B.V.; Barron, A.B.; Scholl, C.; Thamm, M. The Effects of Fat Body Tyramine Level on Gustatory Responsiveness of Honeybees (Apis mellifera) Differ between Behavioral Castes. Front. Syst. Neurosci. 2017, 11, 55. [Google Scholar] [CrossRef] [Green Version]
- Scheiner, R.; Reim, T.; Søvik, E.; Entler, B.V.; Barron, A.B.; Thamm, M. Learning, gustatory responsiveness and tyramine differences across nurse and forager honeybees. J. Exp. Biol. 2017, 220, 1443–1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hesselbach, H.; Seeger, J.; Schilcher, F.; Ankenbrand, M.; Scheiner, R. Chronic exposure to the pesticide flupyradifurone can lead to premature onset of foraging in honeybees Apis mellifera. J. Appl. Ecol. 2020, 57, 609–618. [Google Scholar] [CrossRef] [Green Version]
- Scheiner, R.; Abramson, C.I.; Brodschneider, R.; Crailsheim, K.; Farina, W.M.; Fuchs, S.; Grünewald, B.; Hahshold, S.; Karrer, M.; Koeniger, G.; et al. Standard methods for behavioural studies of Apis mellifera. J. Apic. Res. 2013, 52, 1–58. [Google Scholar] [CrossRef]
- Scholl, C.; Wang, Y.; Krischke, M.; Mueller, M.J.; Amdam, G.V.; Rössler, W. Light exposure leads to reorganization of microglomeruli in the mushroom bodies and influences juvenile hormone levels in the honeybee. Dev. Neurobiol. 2014, 74, 1141–1153. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.P.; Krause, D.M.; Mueller, M.J.; Fekete, A. Accumulation of extra-chloroplastic triacylglycerols in Arabidopsis seedlings during heat acclimation. J. Exp. Bot. 2015, 66, 4517–4526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Mächler, M.; Bolker, B.M. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Mächler, M.; Bolker, B.M.; Walker, S.C. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Hartig, F. Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models; R Package DHARMa Version 0.4.4; Hartig, F.: Regensburg, Germany, 2021. [Google Scholar]
- Lenth, R.V. Least-squares means: The R package lsmeans. J. Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Sagili, R.R.; Pankiw, T. Effects of Brood Pheromone Modulated Brood Rearing Behaviors on Honey Bee (Apis mellifera L.) Colony Growth. J. Insect Behav. 2009, 22, 339–349. [Google Scholar] [CrossRef] [Green Version]
- Kaftanoglu, O.; Linksvayer, T.A.; Page, R.E. Rearing honey bees (Apis mellifera L.) in vitro: Effects of feeding intervals on survival and development. J. Apic. Res. 2010, 49, 311–317. [Google Scholar] [CrossRef]
- Di Pasquale, G.; Salignon, M.; Le Conte, Y.; Belzunces, L.P.; Decourtye, A.; Kretzschmar, A.; Suchail, S.; Brunet, J.-L.; Alaux, C. Influence of Pollen Nutrition on Honey Bee Health: Do Pollen Quality and Diversity Matter? PLoS ONE 2013, 8, e72016. [Google Scholar] [CrossRef] [Green Version]
- Castle, D.; Alkassab, A.T.; Bischoff, G.; Steffan-Dewenter, I.; Pistorius, J. High nutritional status promotes vitality of honey bees and mitigates negative effects of pesticides. Sci. Total Environ. 2021, 806, 151280. [Google Scholar] [CrossRef] [PubMed]
- Ruedenauer, F.A.; Raubenheimer, D.; Kessner-Beierlein, D.; Grund-Mueller, N.; Noack, L.; Spaethe, J.; Leonhardt, S.D. Best be(e) on low fat: Linking nutrient perception, regulation and fitness. Ecol. Lett. 2020, 23, 545–554. [Google Scholar] [CrossRef] [Green Version]
- Tosi, S.; Burgio, G.; Nieh, J.C. A common neonicotinoid pesticide, thiamethoxam, impairs honey bee flight ability. Sci. Rep. 2017, 7, 1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, Y.; Jeong, S.; Lee, D.; Kim, S.; Park, R.J.; Gibson, L.; Zheng, C.; Park, C. Foraging trip duration of honeybee increases during a poor air quality episode and the increase persists thereafter. Ecol. Evol. 2021, 11, 1492–1500. [Google Scholar] [CrossRef]
- Rodney, S.; Kramer, V.J. Probabilistic assessment of nectar requirements for nectar-foraging honey bees. Apidologie 2020, 51, 180–200. [Google Scholar] [CrossRef] [Green Version]
- De Souza, D.A.; Kaftanoglu, O.; De Jong, D.; Page, R.E.; Amdam, G.V.; Wang, Y. Differences in the morphology, physiology and gene expression of honey bee queens and workers reared in vitro versus in situ. Biol. Open 2018, 7, bio036616. [Google Scholar] [CrossRef] [Green Version]
- Vázquez, D.E.; Farina, W.M. Differences in pre-imaginal development of the honey bee Apis mellifera between in vitro and in-hive contexts. Apidologie 2020, 51, 861–875. [Google Scholar] [CrossRef]
- Robinson, G.E. Regulation of honey bee age polyethism by juvenile hormone. Behav. Ecol. Sociobiol. 1987, 20, 329–338. [Google Scholar] [CrossRef]
- Rutz, W.; Gerig, L.; Wille, H.; Lüscher, M. The function of juvenile hormone in adult worker honeybees, Apis mellifera. J. Insect Physiol. 1976, 22, 1485–1491. [Google Scholar] [CrossRef]
- Hagenguth, H.; Rembold, H. Identification of juvenile horm one 3 as the only JH hom olog in all developmental stages of the honey bee. Z. Naturforsch. Sect. C J. Biosci. 1978, 33, 847–850. [Google Scholar] [CrossRef]
- Amdam, G.V.; Aase, A.L.T.O.; Seehuus, S.-C.C.; Kim Fondrk, M.; Norberg, K.; Hartfelder, K. Social reversal of immunosenescence in honey bee workers. Exp. Gerontol. 2005, 40, 939–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toth, A.L.; Robinson, G.E. Worker nutrition and division of labour in honeybees. Anim. Behav. 2005, 69, 427–435. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet A | Royal Jelly | Fructose | Glucose | Yeast | Water |
| [%] | 50 | 6 | 6 | 1 | 37 |
| [g] | 20 | 2.4 | 2.4 | 0.4 | 14.8 |
| Diet B | Royal Jelly | Fructose | Glucose | Yeast | Water |
| [%] | 50 | 7.5 | 7.5 | 1.5 | 33.5 |
| [g] | 20 | 3 | 3 | 0.6 | 13.4 |
| Diet C | Royal Jelly | Fructose | Glucose | Yeast | Water |
| [%] | 50 | 9 | 9 | 2 | 30 |
| [g] | 20 | 3.6 | 3.6 | 0.8 | 12 |
| Analysis | Figure | Treatment | Sample Size | Test | Test-Value | p |
|---|---|---|---|---|---|---|
| Nursing proportion | 1A | Colony | 199 | Chi-square | χ = 8.16 | <0.01 |
| In vitro | 192 | |||||
| Onset of nursing | 1B | Colony | 104 | Mann-Whitney | U = 3055 | 0.56 |
| In vitro | 62 | |||||
| Termination of nursing | 1C | Colony | 104 | Mann-Whitney | U = 3098 | 0.67 |
| In vitro | 62 | |||||
| Nursing span | 1D | Colony | 104 | Mann-Whitney | U = 3223 | 0.99 |
| In vitro | 62 | |||||
| Foraging proportion | 2A | Colony | 1016 | Chi-square | χ = 21.04 | <0.001 |
| In vitro | 1005 | |||||
| Onset of foraging | 2B | Colony | 472 | GLMM | χ = 15.58 | <0.001 |
| Factor treatment | In vitro | 164 | ||||
| Termination of foraging | 2C | Colony | 472 | GLMM | χ = 34.14 | <0.001 |
| Factor treatment | In vitro | 164 | ||||
| Foraging span | 2D | Colony | 472 | GLMM | χ = 18.99 | <0.001 |
| Factor treatment | In vitro | 164 | ||||
| Duration per foraging trip | 2E | Colony | 472 | GLMM | χ = 0.03 | 0.8834 |
| Factor treatment | In vitro | 164 | ||||
| Foraging trips per day | 2F | Colony | 472 | GLMM | χ = 54.31 | <0.001 |
| Factor treatment | In vitro | 164 |
| Analysis | Figure | Treatment | Sample Size | Test | Test Value | p | |
|---|---|---|---|---|---|---|---|
| Weight | 3A | Colony | 63 | GLMM | F = 0.06 | 0.804 | |
| Factor Treatment | In vitro | 66 | |||||
| Weight | 3A | Colony | 63 | GLMM | F = 43.27 | <0.001 | |
| Factor Week | In vitro | 66 | |||||
| Weight | 3A | Colony | 63 | GLMM | F = 6.86 | 0.076 | |
| Interaction Treatment and Week | In vitro | 66 | |||||
| JH III | 3B | Colony | 50 | GLMM | χ = 5.36 | <0.05 | |
| Factor Treatment | In vitro | 50 | |||||
| JH III | 3B | Colony | 50 | GLMM | χ = 146.70 | <0.001 | |
| Factor Week | In vitro | 50 | |||||
| JH III | 3B | Colony | 50 | GLMM | χ = 6.04 | 0.11 | |
| Interaction Treatment and Week | In vitro | 50 | |||||
| JH III | 3B | Week 1 | Colony | 15 | Tukey | t = 0.76 | 0.45 |
| Pairwise Tukey Test | In vitro | 15 | |||||
| 3B | Week 2 | Colony | 15 | Tukey | t = −0.30 | 0.77 | |
| In vitro | 14 | ||||||
| 3B | Week 3 | Colony | 11 | Tukey | t = 1.99 | <0.05 | |
| In vitro | 12 | ||||||
| 3B | Week 4 | Colony | 9 | Tukey | t = 2.60 | <0.05 | |
| In vitro | 9 | ||||||
| TGs | 3C | Colony | 17 | GLMM | χ = 0.04 | <0.05 | |
| Factor Treatment | In vitro | 21 | |||||
| TGs | 3C | Colony | 17 | GLMM | χ = 81.12 | <0.001 | |
| Factor Week | In vitro | 21 | |||||
| TGs | 3C | Colony | 17 | GLMM | χ = 10.92 | <0.05 | |
| Interaction Treatment and Week | In vitro | 21 | |||||
| TGs | 3C | Week 1 | Colony | 5 | Tukey | t = −1.43 | 0.16 |
| Pairwise Tukey Test | In vitro | 3 | |||||
| 3C | Week 2 | Colony | 4 | Tukey | t = 0.70 | 0.49 | |
| In vitro | 4 | ||||||
| 3C | Week 3 | Colony | 5 | Tukey | t = 1.28 | 0.21 | |
| In vitro | 5 | ||||||
| 3C | Week 4 | Colony | 3 | Tukey | t = 3.31 | <0.01 | |
| In vitro | 9 | ||||||
| GRS | 3D | Colony | 17 | GLMM | χ = 3.69 | 0.055 | |
| Factor Treatment | In vitro | 21 | |||||
| GRS | 3D | Colony | 17 | GLMM | χ = 18.43 | <0.001 | |
| Factor Week | In vitro | 21 | |||||
| GRS | 3D | Colony | 17 | GLMM | χ = 5.68 | 0.13 | |
| Interaction Treatment and Week | In vitro | 21 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schilcher, F.; Hilsmann, L.; Rauscher, L.; Değirmenci, L.; Krischke, M.; Krischke, B.; Ankenbrand, M.; Rutschmann, B.; Mueller, M.J.; Steffan-Dewenter, I.; et al. In Vitro Rearing Changes Social Task Performance and Physiology in Honeybees. Insects 2022, 13, 4. https://doi.org/10.3390/insects13010004
Schilcher F, Hilsmann L, Rauscher L, Değirmenci L, Krischke M, Krischke B, Ankenbrand M, Rutschmann B, Mueller MJ, Steffan-Dewenter I, et al. In Vitro Rearing Changes Social Task Performance and Physiology in Honeybees. Insects. 2022; 13(1):4. https://doi.org/10.3390/insects13010004
Chicago/Turabian StyleSchilcher, Felix, Lioba Hilsmann, Lisa Rauscher, Laura Değirmenci, Markus Krischke, Beate Krischke, Markus Ankenbrand, Benjamin Rutschmann, Martin J. Mueller, Ingolf Steffan-Dewenter, and et al. 2022. "In Vitro Rearing Changes Social Task Performance and Physiology in Honeybees" Insects 13, no. 1: 4. https://doi.org/10.3390/insects13010004
APA StyleSchilcher, F., Hilsmann, L., Rauscher, L., Değirmenci, L., Krischke, M., Krischke, B., Ankenbrand, M., Rutschmann, B., Mueller, M. J., Steffan-Dewenter, I., & Scheiner, R. (2022). In Vitro Rearing Changes Social Task Performance and Physiology in Honeybees. Insects, 13(1), 4. https://doi.org/10.3390/insects13010004