
Towards a Knowledge-Based Decision Support System for Integrated Control of Woolly Apple Aphid, Eriosoma lanigerum, with Maximal Biological Suppression by the Parasitoid Aphelinus mali


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Monitoring
2.2. Modelling
2.3. Field Study in Apple Orchard
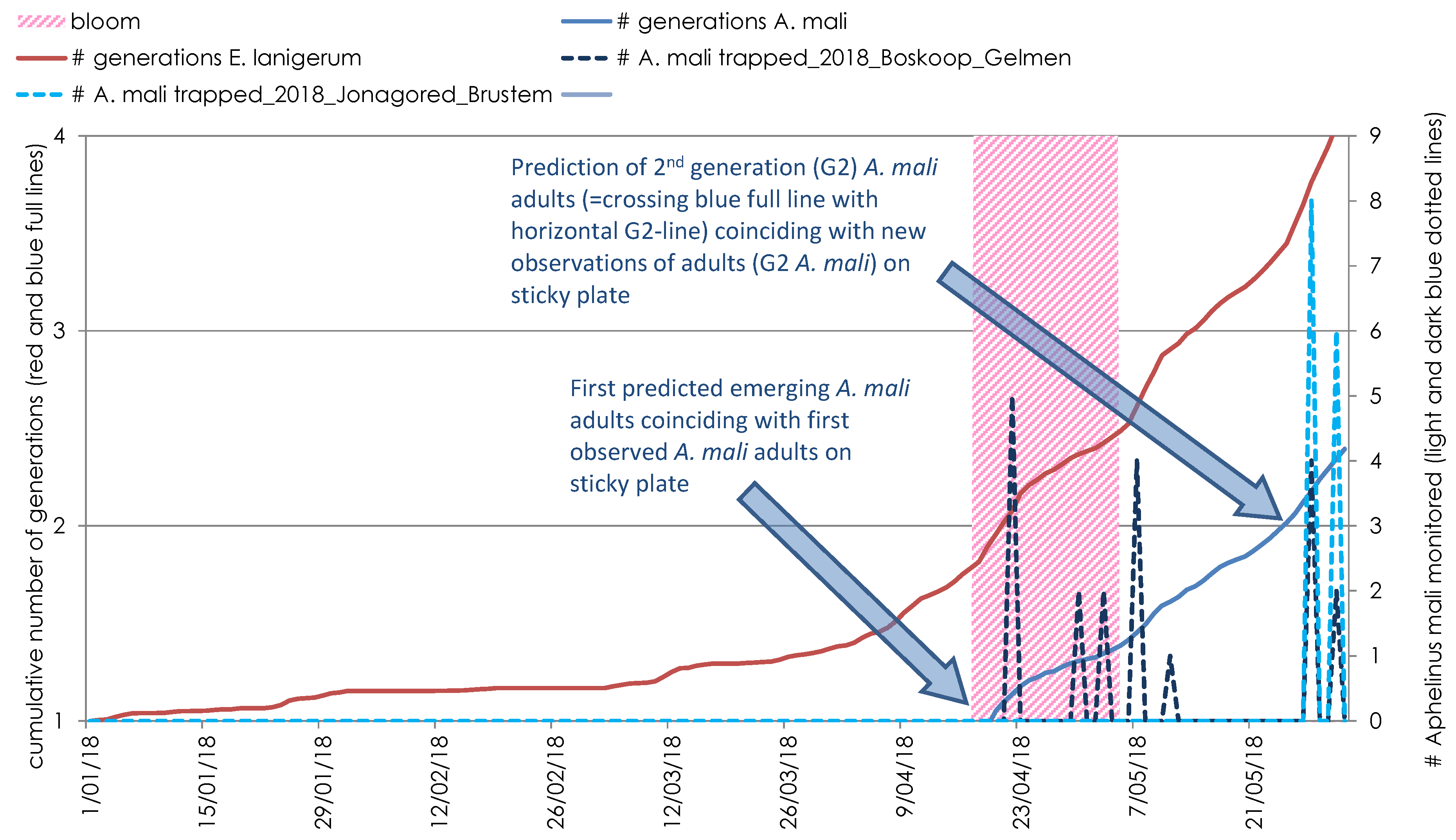
3. Results
3.1. Polynomial Regression Models for A. mali and E. lanigerum
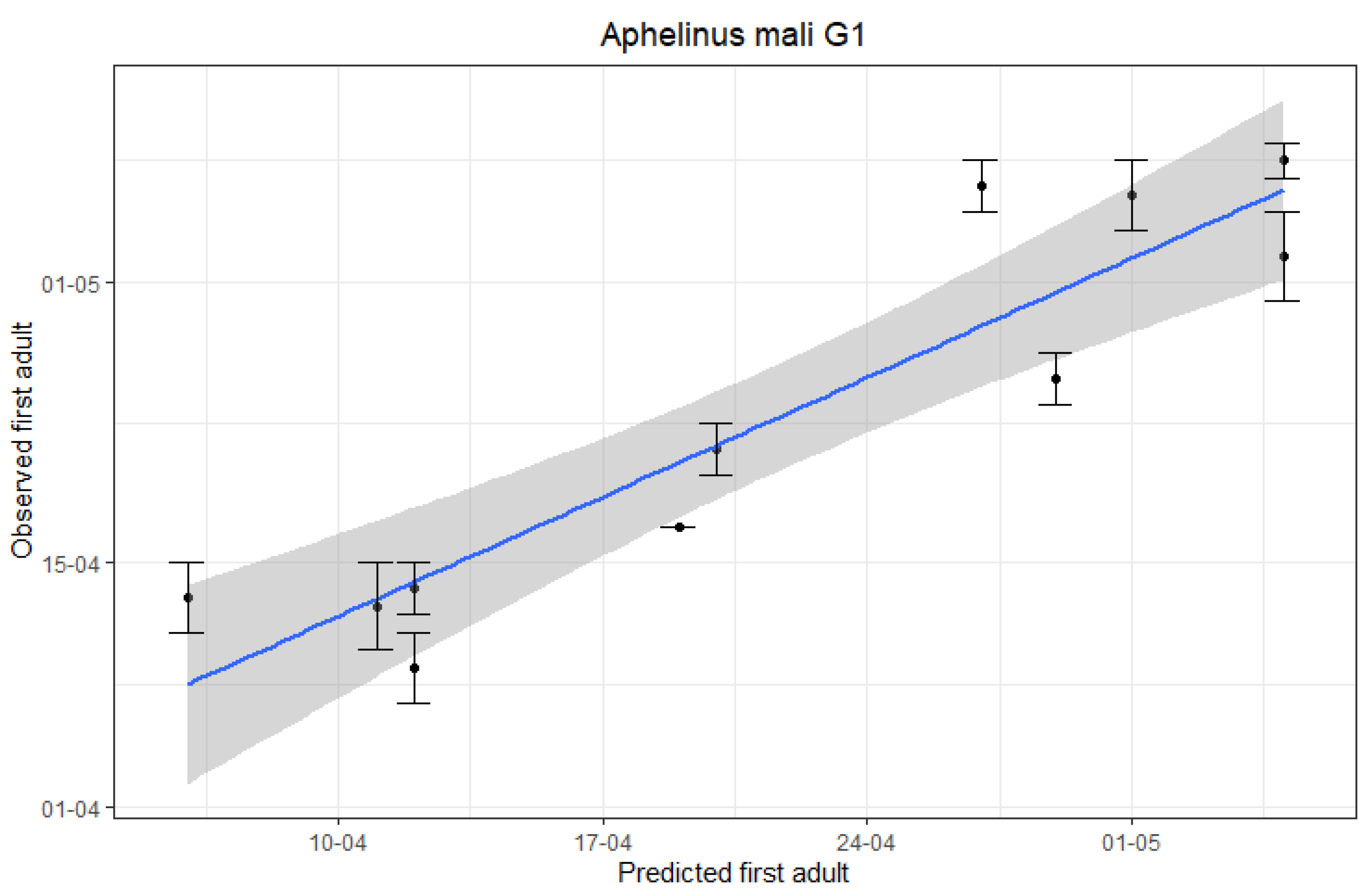
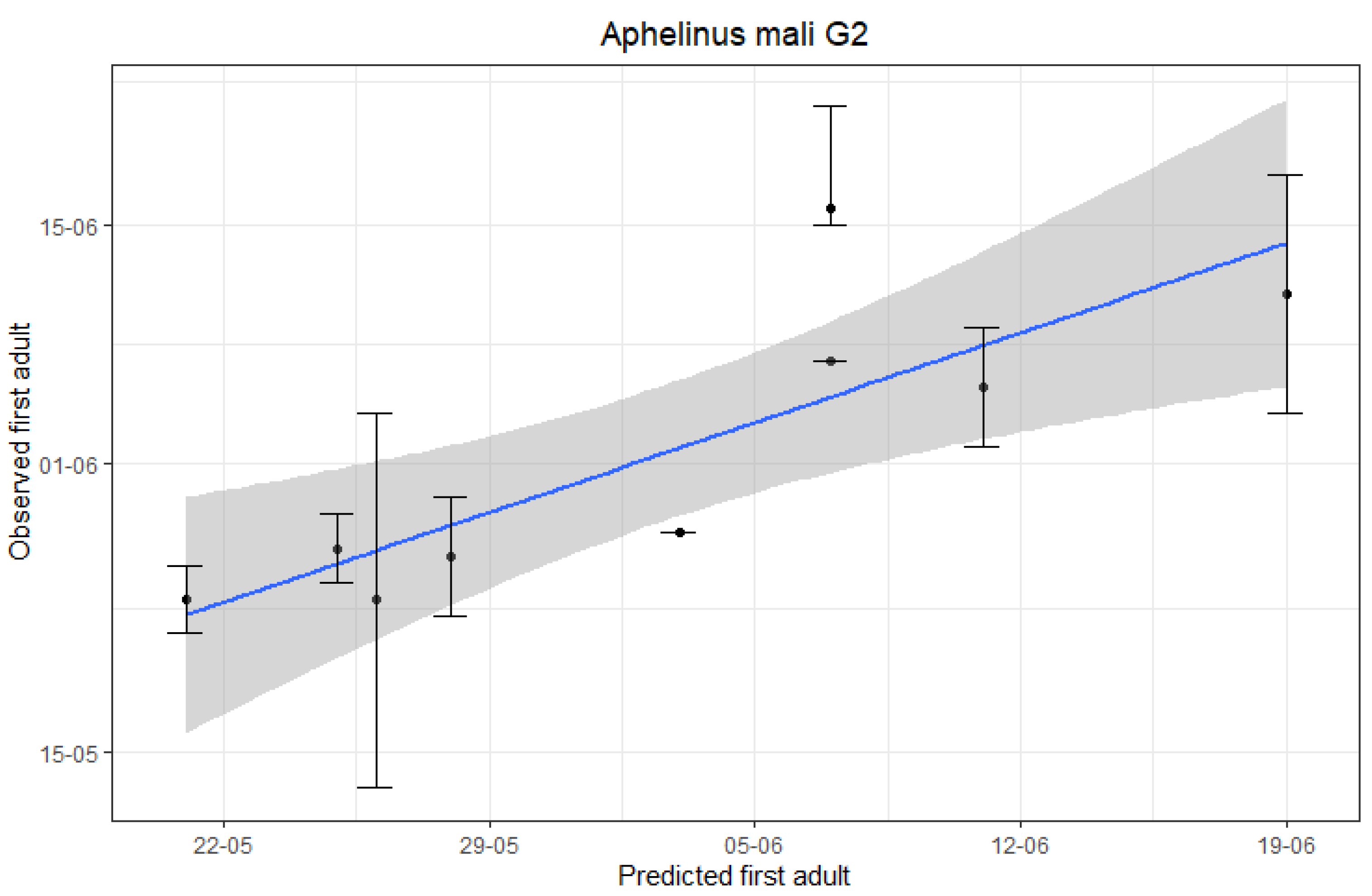
3.2. Accuracy of A. mali First and Second Generation Adults Emergence Predictions
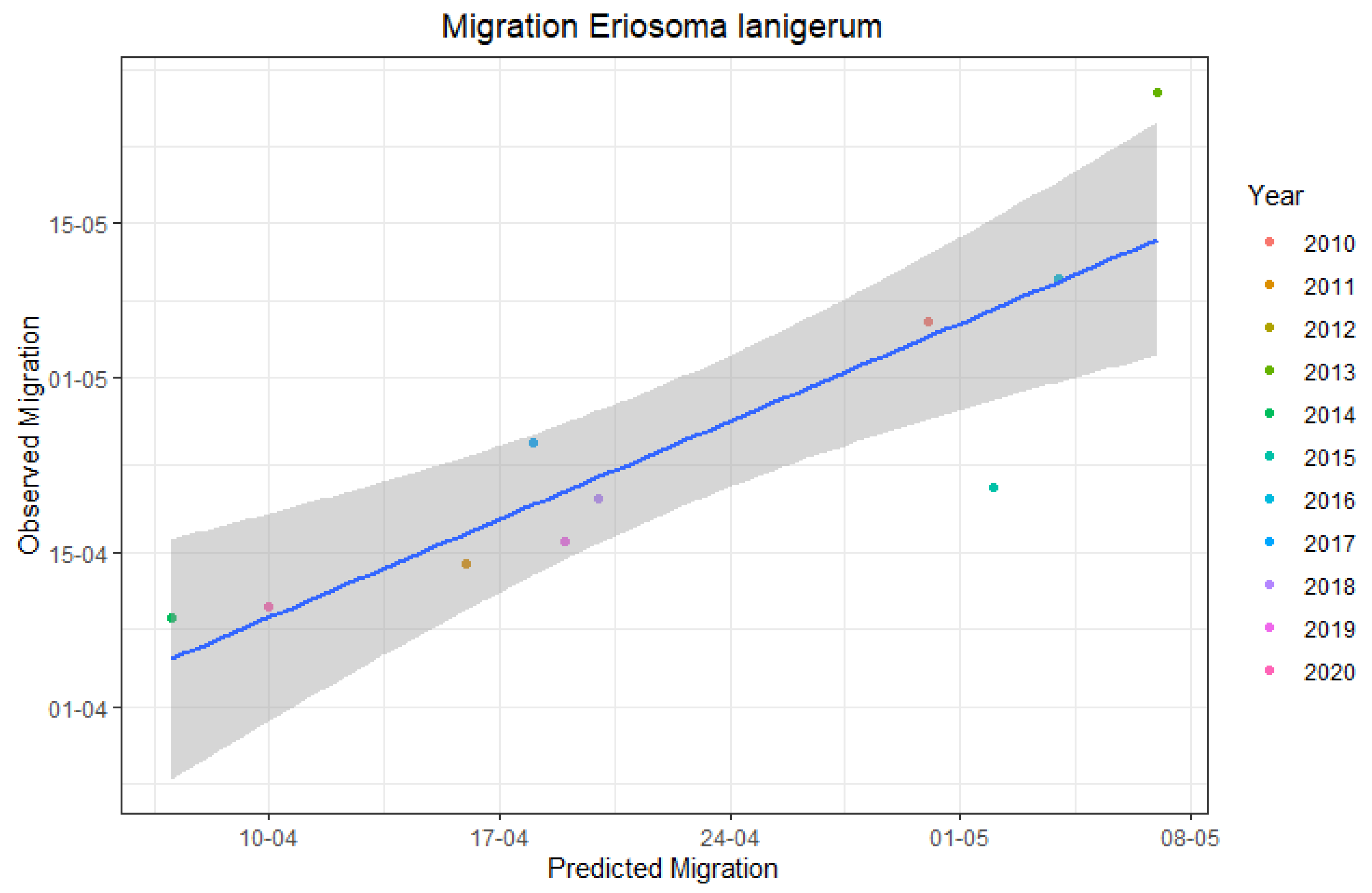
3.3. Accuracy of E. lanigerum Migration Predictions
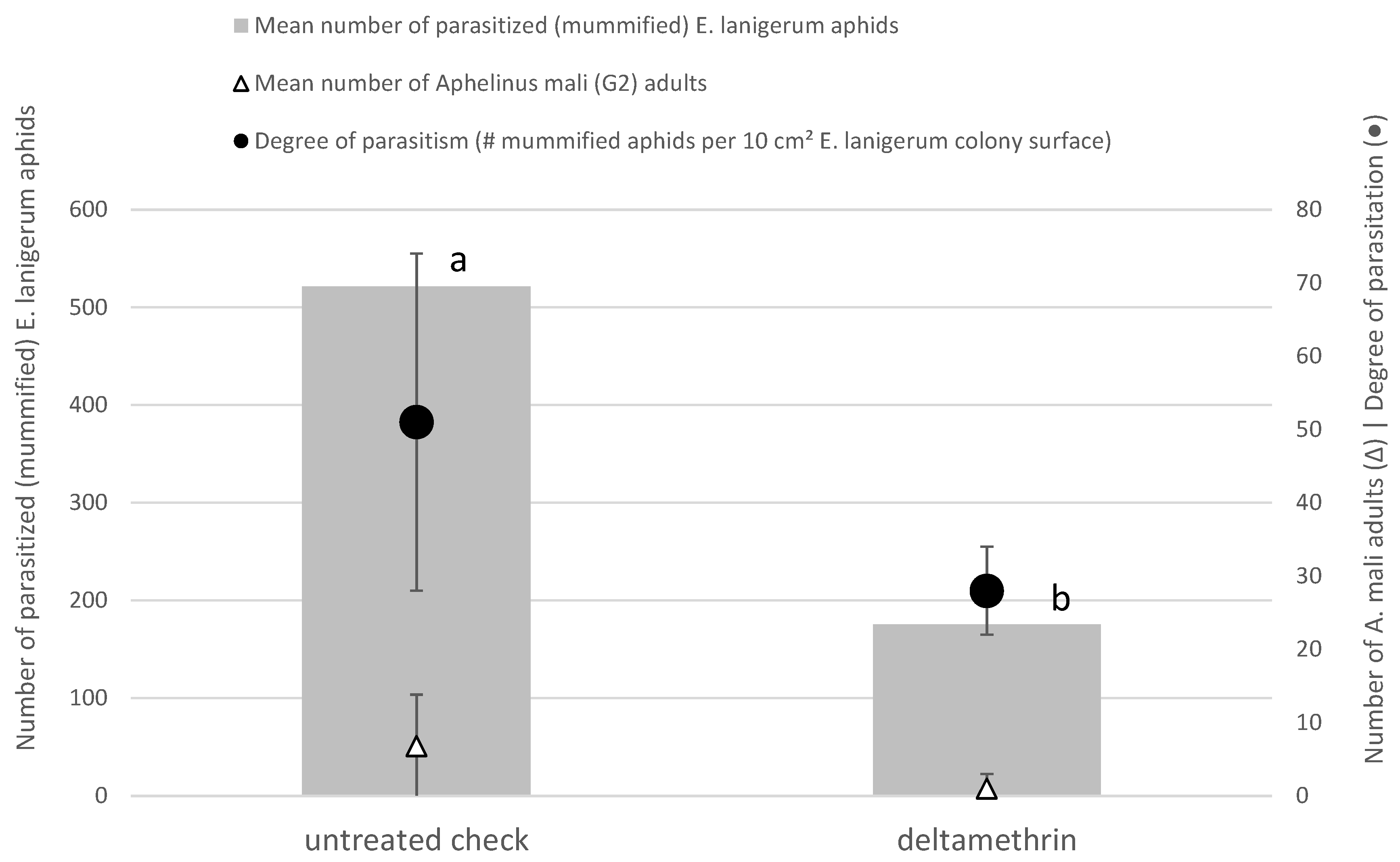
3.4. Field Study on Side-Effects on A. mali and Impact on Biologicol Control of E. lanigerum
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beers, E.H.; Cockfield, S.D.; Gontijo, L.M. Seasonal phenology of woolly apple aphid (Hemiptera: Aphididae) in Central Washington. Environ. Entomol. 2010, 39, 286–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lordan, J.; Alegre, S.; Gatius, F.; Sarasúa, M.J.; Alins, G. Woolly apple aphid Eriosoma lanigerum Hausmann ecology and its relationship with climatic variables and natural enemies in Mediterranean areas. Bull. Entomol. Res. 2015, 105, 60–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goossens, D.; Bangels, E.; Belien, T.; Schoevaerts, C.; De Maeyer, L. Optimal profit of the parasitation by Aphelinus mali in an IPM complementary strategy for the control of Eriosoma lanigerum. Commun. Agric. Appl. Biol. Sci. 2011, 76, 457–465. [Google Scholar] [PubMed]
- Stokwe, N.F.; Malan, A.P. Woolly apple aphid, Eriosoma lanigerum (Hausmann), in South Africa: Biology and management practices, with focus on the potential use of entomopathogenic nematodes and fungi. Afr. Entomol. 2016, 24, 267–278. [Google Scholar] [CrossRef]
- Beliën, T.; Bangels, E.; Peusens, G.; Goossens, D.; Berkvens, N.; Viaene, N. Towards improved control of woolly apple aphid (Eriosoma lanigerum) in integrated fruit production. Acta Hortic. 2011, 917, 15–22. [Google Scholar] [CrossRef]
- Beers, E.H.; Cockfield, S.D.; Fazio, G. Biology and management of woolly apple aphid, Eriosoma lanigerum (Hausmann), in Washington state. IOBC-WPRS Bull. 2007, 30, 37–42. [Google Scholar]
- Evenhuis, H.H. Een oecologisch onderzoek over de appelbloedluis, Eriosoma lanigerum (Hausm.), en haar parasiet Aphelinus mali (Hald.) in Nederland. Tijdschr. Over Plantenziekten 1958, 64, 1–103. [Google Scholar] [CrossRef]
- Cross, J.V.; Solomon, M.G.; Babandreier, D.; Blommers, L.; Easterbrook, M.A.; Jay, C.N.; Jenser, G.; Jolly, R.L.; Kuhlmann, U.; Lilley, R.; et al. Biocontrol of pests of apples and pears in northern and central Europe: 2. Parasitoids. Biocontrol Sci. Technol. 1999, 9, 277–314. [Google Scholar] [CrossRef]
- Sandanayaka, W.R.; Bus, V.G. Evidence of sexual reproduction of woolly apple aphid, Eriosoma lanigerum, in New Zealand. J. Insect Sci. 2005, 5, 27. [Google Scholar] [CrossRef] [Green Version]
- Gontijo, L.M.; Cockfield, S.D.; Beers, E.H. Natural enemies of woolly apple aphid (Hemiptera: Aphididae) in Washington State. Environ. Entomol. 2012, 41, 1364–1371. [Google Scholar] [CrossRef] [Green Version]
- Orpet, R.J.; Crowder, D.W.; Jones, V.P. Woolly Apple Aphid Generalist Predator Feeding Behavior Assessed through Video Observation in an Apple Orchard. J. Insect Behav. 2019, 32, 153–163. [Google Scholar] [CrossRef]
- Nicholas, A.H.; Spooner-Hart, R.N.; Vickers, R.A. Abundance and natural control of the woolly aphid Eriosoma lanigerum in an Australian apple orchardIPM program. BioControl 2005, 50, 271–291. [Google Scholar] [CrossRef]
- Penalver-Cruz, A.; Alvarez, D.; Lavandero, B. Do hedgerows influence the natural biological control of woolly apple aphids in orchards? J. Pest Sci. 2020, 93, 219–234. [Google Scholar] [CrossRef]
- Short, B.D.; Bergh, J.C. Feeding and Egg Distribution Studies of Heringia calcarata (Diptera: Syrphidae), a Specialized Predator of Woolly Apple Aphid (Homoptera: Eriosomatidae) in Virginia Apple Orchards. J. Econ. Entomol. 2004, 97, 813–819, 817. [Google Scholar] [CrossRef]
- Quarrell, S.R.; Corkrey, R.; Allen, G.R. Predictive thresholds for forecasting the compatibility of Forficula auricularia and Aphelinus mali as biological control agents against woolly apple aphid in apple orchards. Biocontrol 2017, 62, 243–256. [Google Scholar] [CrossRef]
- Bergh, J.C.; Stallings, J.W. Field evaluations of the contribution of predators and the parasitoid, Aphelinus mali, to biological control of woolly apple aphid, Eriosoma lanigerum, in Virginia, USA. Biocontrol 2016, 61, 155–165. [Google Scholar] [CrossRef]
- Asante, S.K.; Danthanarayana, W. Development of Aphelinus mali an endoparasitoid of woolly apple aphid, Eriosoma lanigerum at different temperatures. Entomol. Exp. Appl. 1992, 65, 31–37. [Google Scholar] [CrossRef]
- Mueller, T.F.; Blommers, L.H.M.; Mols, P.J.M. Woolly apple aphid (Eriosoma lanigerum Hausm., Hom., Aphidae) parasitism by Aphelinus mali Hal. (Hym., Aphelinidae) in relation to host stage and host colony size, shape and location. J. Appl. Entomol. 1992, 114, 143–154. [Google Scholar] [CrossRef]
- Mols, P.J.; Boers, J.M. Comparison of a Canadian and a Dutch strain of the parasitoid Aphelinus mali (Hald) (Hym., Aphelinidae) for control of woolly apple aphid Eriosoma lanigerum (Haussmann) (Hom., Aphididae) in the Netherlands: A simulation approach. J. Appl. Entomol. 2001, 125, 255–262. [Google Scholar] [CrossRef]
- Heunis, J.M.; Pringle, K.L. The susceptibility of Aphelinus mali (Haldeman), a parasitoid of Eriosoma lanigerum (Hausmann), to pesticides used in apple orchards in the Elgin area, Western Cape Province, South Africa. Afr. Entomol. 2003, 11, 91–95. [Google Scholar]
- Cohen, H.; Horowitz, A.R.; Nestel, D.; Rosen, D. Susceptibility of the woolly apple aphid parasitoid, Aphelinus mali (Hym.: Aphelinidae), to common pesticides used in apple orchards in Israel. Entomophaga 1996, 41, 225–233. [Google Scholar] [CrossRef]
- Asante, S.K.; Danthanarayana, W.; Heatwole, H. Bionomics and population growth statistics of apterous virginoparae of woolly apple aphid, Eriosoma lanigerum, at constant temperatures. Entomol. Exp. Appl. 1991, 60, 261–270. [Google Scholar] [CrossRef]
- Wallis, D.; Shaw, P.W. Evaluation of coloured sticky traps for monitoring beneficial insects in apple orchards. N. Z. Plant Prot. 2008, 61, 328–332. [Google Scholar] [CrossRef]
- Rebaudo, F.; Struelens, Q.; Dangles, O. Modelling temperature-dependent development rate and phenology in arthropods: The devRate package for r. Methods Ecol. Evol. 2018, 9, 1144–1150. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.R.; Anil, S.; Verma, S.P. Overwintering, emergence pattern, sex ratio and mating strategy of Aphelinus mali (Haldeman), an endoparasitoid of the woolly apple aphid. J. Biol. Control 2007, 21, 113–118. [Google Scholar]
- Piñeiro, G.; Perelman, S.; Guerschman, J.P.; Paruelo, J.M. How to evaluate models: Observed vs. predicted or predicted vs. observed? Ecol. Model. 2008, 216, 316–322. [Google Scholar] [CrossRef]
- Tedeschi, L.O. Assessment of the adequacy of mathematical models. Agric. Syst. 2006, 89, 225–247. [Google Scholar] [CrossRef]
- Candolfi, M.P.; Blümel, S.; Forster, R.; Bakker, F.M.; Grimm, C.; Hassan, S.A.; Heimbach, U.; Mead-Briggs, M.A.; Reber, B.; Schmuck, R.; et al. (Eds.) Guidelines to Evaluate Side-Effects of Plant Protection Products to Non-Target Arthropods; IOBC/WPRS: Gent, Belgium, 2000; Volume IX, p. 158. ISBN 92-9067-129-7. [Google Scholar]
- EPPO. PP1/254(1)—Eriosoma Lanigerum on Apple. OEPP/EPPO Bull. 2007, 37, 92–94. [Google Scholar] [CrossRef]
- EPPO. PP1/239(3)—Dose Expression for Plant Protection Products. OEPP/EPPO Bull. 2021, 51, 10–33. [Google Scholar] [CrossRef]
- Asante, S.K.; Danthanarayana, W.; Cairns, S.C. Spatial and Temporal Distribution Patterns of Eriosoma lanigerum (Homoptera: Aphididae) on Apple. Environ. Entomol. 1993, 22, 1060–1065. [Google Scholar] [CrossRef]
- Damos, P.; Colomar, L.A.; Ioriatti, C. Integrated Fruit Production and Pest Management in Europe: The Apple Case Study and How Far We Are From the Original Concept? Insects 2015, 6, 626–657. [Google Scholar] [CrossRef]
- Wearing, C.H.; Attfield, B.A.; Colhoun, K. Biological control of woolly apple aphid, Eriosoma lanigerum (Hausmann), during transition to integrated fruit production for pipfruit in Central Otago, New Zealand. N. Z. J. Crop Hortic. Sci. 2010, 38, 255–273. [Google Scholar] [CrossRef]
- Beers, E.H.; Horton, D.R.; Miliczky, E. Pesticides used against Cydia pomonella disrupt biological control of secondary pests of apple. Biol. Control 2016, 102, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Beers, E.H. Effect of Trap Color and Orientation on the Capture of Aphelinus mali (Hymenoptera: Aphelinidae), a Parasitoid of Woolly Apple Aphid (Hemiptera: Aphididae). J. Econ. Entomol. 2012, 105, 1342–1349. [Google Scholar] [CrossRef] [Green Version]
- Bradley, S.J.; Murrell, V.C.; Shaw, P.W.; Walker, J.T.S. Effect of Orchard Pesticides on Aphelinus mali, the Woolly Apple Aphid Parasitoid. In Proceedings of the 50th Conference of the Plant Protection Society of New Zealand, Lincoln University, Lincoln, New Zealand, 18–21 August 1997; New Zealand Plant Protection Society Inc.: Rotorua, New Zealand, 1997; pp. 218–222. [Google Scholar] [CrossRef] [Green Version]
- Su, M.; Tan, X.; Yang, Q.; Wan, F.; Zhou, H. Temperature adaptability of two clades of Aphelinus mali (Hymenoptera: Aphelinidae) in China. Egypt. J. Biol. Pest Control 2018, 28, 16. [Google Scholar] [CrossRef] [Green Version]
- Schoevaerts, C.; Goossens, D.; D’Haemer, K.; Van Dyck, H.; De Maeyer, L. The Multitarget Use of Spirotetramat (Movento (R) 100 SC): Simultaneous Control of Key Pests in Apples. In XXVIII International Horticultural Congress on Science and Horticulture for People; Hale, C., Ed.; Acta Horticulturae; International Society for Horticultural Science: Leuven, Belgium, 2011; pp. 69–76. [Google Scholar]
- Fernandez, D.E.; Beers, E.H.; Brunner, J.F.; Doerr, M.D.; Dunley, J.E. Effects of seasonal mineral oil applications on the pest and natural enemy complexes of apple. J. Econ. Entomol. 2005, 98, 1630–1640. [Google Scholar] [CrossRef] [PubMed]
- Ikbal, C.; Pavela, R. Essential oils as active ingredients of botanical insecticides against aphids. J. Pest Sci. 2019, 92, 971–986. [Google Scholar] [CrossRef]
- Ateyyat, M.; Al-Antary, T. Susceptibility of nine apple cultivars to woolly apple aphid, Eriosoma lanigerum (Homoptera: Aphididae) in Jordan. Int. J. Pest Manag. 2009, 55, 79–84. [Google Scholar] [CrossRef]
- Tobutt, K.R.; Boskovic, R.; Roche, P. Incompatibility and resistance to woolly apple aphid in apple. Plant Breed. 2000, 119, 65–69. [Google Scholar] [CrossRef]
- Rousselin, A.; Bevacqua, D.; Sauge, M.H.; Lescourret, F.; Mody, K.; Jordan, M.O. Harnessing the aphid life cycle to reduce insecticide reliance in apple and peach orchards. A review. Agron. Sustain. Dev. 2017, 37, 38. [Google Scholar] [CrossRef] [Green Version]
- Tonnang, H.E.Z.; Hervé, B.D.B.; Biber-Freudenberger, L.; Salifu, D.; Subramanian, S.; Ngowi, V.B.; Guimapi, R.Y.A.; Anani, B.; Kakmeni, F.M.M.; Affognon, H.; et al. Advances in crop insect modelling methods—Towards a whole system approach. Ecol. Model. 2017, 354, 88–103. [Google Scholar] [CrossRef] [Green Version]
- Neta, A.; Gafni, R.; Elias, H.; Bar-Shmuel, N.; Shaltiel-Harpaz, L.; Morin, E.; Morin, S. Decision support for pest management: Using field data for optimizing temperature-dependent population dynamics models. Ecol. Model. 2021, 440, 109402. [Google Scholar] [CrossRef]
- Belien, T.; Lux, S.; De Ketelaere, B.; Bylemans, D. Improving monitoring and forecasting in integrated management of fruit arthropod pests. In Integrated Management of Diseases and Insect Pests of Tree Fruit; Xu, X., Fountain, M., Eds.; Burleigh Dodds Series in Agricultural Science; Burleigh Dodds Science Publishing: Cambridge, UK, 2019; pp. 1–44. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Predicted First G1 Adults | Observed First G1 Adults 1 | Predicted First G2 Adults | Observed First G2 Adults 2 |
|---|---|---|---|---|
| 2010 | 28/04 | 24/04 | 11/06 | NA 3 |
| 2011 | 11/04 | 04/04 | 20/05 | NA 3 |
| 2012 | 30/04 | 05/05 | 06/06 | 15/06 |
| 2013 | 04/05 | 07/05 | 18/06 | 10/06 |
| 2014 | 05/04 | 12/04 | 25/05 | 23/05 |
| 2015 | 26/04 | 05/05 | 10/06 | 04/06 |
| 2016 | 04/05 | 01/05 | 06/06 | 06/06 |
| 2017 | 11/04 | 08/04 | 27/05 | 25/05 |
| 2018 | 19/04 | 20/04 | 24/05 | 26/05 |
| 2019 | 18/04 | 16/04 | 02/06 | 27/05 |
| 2020 | 10/04 | 11/04 | 20/05 | 23/05 |
| Year | Predicted First Migration | Observed First Migration |
|---|---|---|
| 2010 | 30/04 | 06/05 |
| 2011 | 16/04 | 14/04 |
| 2012 | 29/04 | NA 1 |
| 2013 | 07/05 | 27/05 |
| 2014 | 07/04 | 09/04 |
| 2015 | 02/05 | 21/04 |
| 2016 | 03/05 | 09/05 |
| 2017 | 18/04 | 25/04 |
| 2018 | 20/04 | 20/04 |
| 2019 | 19/04 | 16/04 |
| 2020 | 09/04 | 09/04 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bangels, E.; Alhmedi, A.; Akkermans, W.; Bylemans, D.; Belien, T. Towards a Knowledge-Based Decision Support System for Integrated Control of Woolly Apple Aphid, Eriosoma lanigerum, with Maximal Biological Suppression by the Parasitoid Aphelinus mali. Insects 2021, 12, 479. https://doi.org/10.3390/insects12060479
Bangels E, Alhmedi A, Akkermans W, Bylemans D, Belien T. Towards a Knowledge-Based Decision Support System for Integrated Control of Woolly Apple Aphid, Eriosoma lanigerum, with Maximal Biological Suppression by the Parasitoid Aphelinus mali. Insects. 2021; 12(6):479. https://doi.org/10.3390/insects12060479
Chicago/Turabian StyleBangels, Eva, Ammar Alhmedi, Wannes Akkermans, Dany Bylemans, and Tim Belien. 2021. "Towards a Knowledge-Based Decision Support System for Integrated Control of Woolly Apple Aphid, Eriosoma lanigerum, with Maximal Biological Suppression by the Parasitoid Aphelinus mali" Insects 12, no. 6: 479. https://doi.org/10.3390/insects12060479
APA StyleBangels, E., Alhmedi, A., Akkermans, W., Bylemans, D., & Belien, T. (2021). Towards a Knowledge-Based Decision Support System for Integrated Control of Woolly Apple Aphid, Eriosoma lanigerum, with Maximal Biological Suppression by the Parasitoid Aphelinus mali. Insects, 12(6), 479. https://doi.org/10.3390/insects12060479