
Photo-Selective Nets and Pest Control: Searching Behavior of the Codling Moth Parasitoid Mastrus ridens (Hymenoptera: Ichneumonidae) under Varying Light Quantity and Quality Conditions

 ,
,
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
2.1. Insects
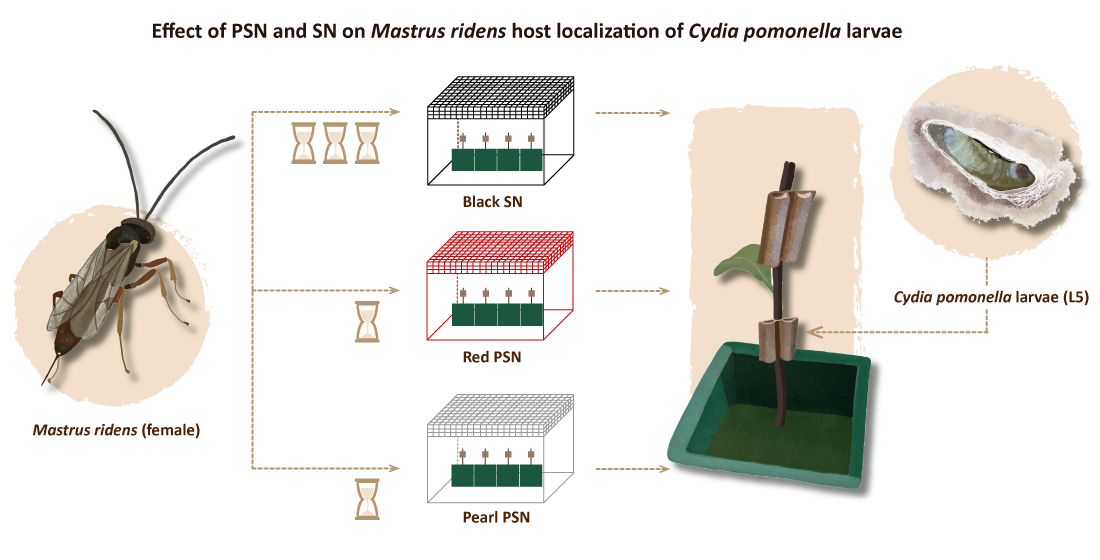
2.2. Host Localization
2.3. Parasitism
2.4. Parasitoid Behavior
2.5. Data Analysis
3. Results
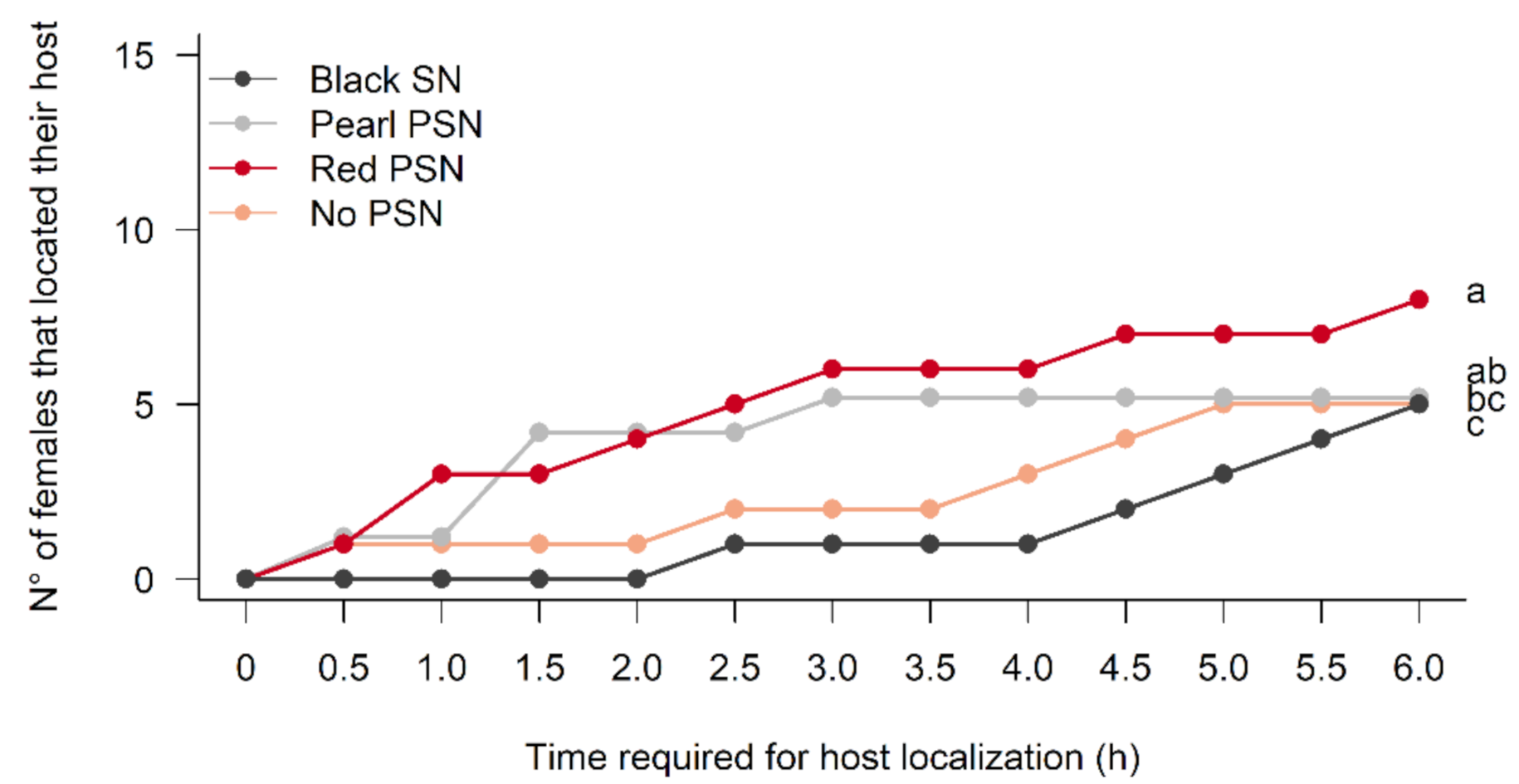
3.1. Effects of the PSN and Black SN on the Host Localization Capability of M. ridens
3.2. Effects of the PSN and Black SN on the Parasitism Level of M. ridens
3.3. Effect of the PSN and the Black SN on the Behavior of M. ridens Females
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moretti, C.L.; Mattos, L.M.; Calbo, A.G.; Sargent, S.A. Climate changes and potential impacts on postharvest quality of fruit and vegetable crops: A review. Food Res. Int. 2010, 43, 1824–1832. [Google Scholar] [CrossRef]
- Wheeler, T.; von Braun, J. Climate change impacts on global food security. Science 2013, 341, 508–513. [Google Scholar] [CrossRef]
- Kalcsits, L.; Musacchi, S.; Layne, D.R.; Schmidt, T.; Mupambi, G.; Serra, S.; Mendoza, M.; Asteggiano, L.; Jarolmasjed, S.; Sankaran, S.; et al. Above and below-ground environmental changes associated with the use of photoselective protective netting to reduce sunburn in apple. Agric. For. Meteorol. 2017, 237–238, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Mupambi, G.; Anthony, B.M.; Layne, D.R.; Musacchi, S.; Serra, S.; Schmidt, T.; Kalcsits, L.A. The influence of protective netting on tree physiology and fruit quality of apple: A review. Sci. Hortic. 2018, 236, 60–72. [Google Scholar] [CrossRef]
- Iglesias, I.; Alegre, S. The effect of anti-hail nets on fruit protection, radiation, temperature, quality and profitability of “Mondial Gala” apples. J. Appl. Hortic. 2006, 8, 91–100. [Google Scholar] [CrossRef]
- Lobos, G.A.; Retamales, J.B.; Hancock, J.F.; Flore, J.A.; Cobo, N.; del Pozo, A. Spectral irradiance, gas exchange characteristics and leaf traits of Vaccinium corymbosum L. “Elliott” grown under photo-selective nets. Environ. Exp. Bot. 2012, 75, 142–149. [Google Scholar] [CrossRef]
- Shahak, Y.; Ratner, K.; Giller, Y.E.; Zur, N.; Or, E.; Gussakovsky, E.E.; Stern, R.; Sarig, P.; Raban, E.; Harcavi, E.; et al. Improving solar energy utilization, productivity and fruit quality in orchards and vineyards by photoselective netting. Acta Hortic. 2008, 772, 65–72. [Google Scholar] [CrossRef]
- Tinyane, P.P.; Sivakumar, D.; Soundy, P. Influence of photo-selective netting on fruit quality parameters and bioactive compounds in selected tomato cultivars. Sci. Hortic. 2013, 161, 340–349. [Google Scholar] [CrossRef]
- Ayala-Tafoya, F.; Yañez Juárez, M.; López Orona, C.; Medina López, R.; de Jesus Velásquez Alcaraz, T.; Díaz Valdés, T. Sunlight transmitted by colored shade nets on photosynthesis and yield of cucumber. Cienc. Rural 2018, 48, 1–9. [Google Scholar] [CrossRef]
- Shahak, Y. Photo-selective netting for improved performance of horticultural crops. A review of ornamental and vegetable studies carried out in Israel. Acta Hortic. 2008, 161–168. [Google Scholar] [CrossRef]
- Ben-Yakir, D.; Antignus, Y.; Offir, Y.; Shahak, Y. Colored shading nets impede insect invasion and decrease the incidences of insect-transmitted viral diseases in vegetable crops. Entomol. Exp. Appl. 2012, 144, 249–257. [Google Scholar] [CrossRef]
- Elad, Y.; Messika, Y.; Brand, M.; David, D.R.; Sztejnberg, A. Effect of colored shade nets on pepper powdery mildew (Leveillula taurica). Phytoparasitica 2007, 35, 285–299. [Google Scholar] [CrossRef]
- Shahak, Y.; Gussakovsky, E.E.; Gal, E.; Ganelevin, R. ColorNets: Crop protection and light-quality manipulation in one technology. Acta Hortic. 2004, 659, 143–151. [Google Scholar] [CrossRef]
- Bastías, R.M.; Corelli-Grappadelli, L. Light quality management in fruit orchards: Physiological and technological aspects. Chil. J. Agric. Res. 2012, 72, 574–581. [Google Scholar] [CrossRef]
- Ilić, Z.S.; Milenković, L.; Šunić, L.; Barać, S.; Mastilović, J.; Kevrešan, Ž.; Fallik, E. Effect of shading by coloured nets on yield and fruit quality of sweet pepper. Zemdirb. Agric. 2017, 104, 53–62. [Google Scholar] [CrossRef]
- Arthurs, S.P.; Stamps, R.H.; Giglia, F.F. Environmental modification inside photoselective shadehouses. HortScience 2013, 48, 975–979. [Google Scholar] [CrossRef]
- Hemming, S.; Dueck, T.; Janse, J.; van Noort, F. The effect of diffuse light on crops. Acta Hortic. 2008, 801, 1293–1300. [Google Scholar] [CrossRef] [Green Version]
- Selahle, K.M.; Sivakumar, D.; Jifon, J.; Soundy, P. Postharvest responses of red and yellow sweet peppers grown under photo-selective nets. Food Chem. 2015, 173, 951–956. [Google Scholar] [CrossRef]
- Antignus, Y.; Lapidot, M.; Hadar, D.; Messika, Y.; Cohen, S. Ultraviolet-absorbing screens serve as optical barriers to protect crops from virus and insect pests. J. Econ. Entomol. 1998, 91, 1401–1405. [Google Scholar] [CrossRef]
- Chyzik, R.; Dobrinin, S.; Antignus, Y. Effect of a UV-deficient environment on the biology and flight activity of Myzus persicae and its hymenopterous parasite Aphidius matricariae. Phytoparasitica 2003, 31, 467–477. [Google Scholar] [CrossRef]
- Dáder, B.; Gwynn-Jones, D.; Moreno, A.; Winters, A.; Fereres, A. Impact of UV-A radiation on the performance of aphids and whiteflies and on the leaf chemistry of their host plants. J. Photochem. Photobiol. B Biol. 2014, 138, 307–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antignus, Y.; Nestel, D.; Cohen, S.; Lapidot, M. Ultraviolet-deficient greenhouse environment affects whitefly attraction and flight-behavior. Environ. Entomol. 2001, 30, 394–399. [Google Scholar] [CrossRef]
- Legarrea, S.; Betancourt, M.; Plaza, M.; Fraile, A.; García-Arenal, F.; Fereres, A. Dynamics of nonpersistent aphid-borne viruses in lettuce crops covered with UV-absorbing nets. Virus Res. 2012, 165, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiel, E.; Messika, Y.; Steinberg, S.; Antignus, Y. The effect of UV-absorbing plastic sheet on the attraction and host location ability of three parasitoids: Aphidius colemani, Diglyphus isaea and Eretmocerus mundus. BioControl 2006, 51, 65–78. [Google Scholar] [CrossRef]
- Umanzor, C.; Bastías, R.; Wilckens, R.; Quezada, C. Influencia del uso de malla perla y roja en las condiciones micro-climáticas, daño por sol y desarrollo de color de frutos en huertos de manzanos cvs. gala y fuji. Rev. Bras. Frutic. 2016, 39, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Welter, S.C. Codling moth. In Encyclopedia of Insects; Resh, V.H., Cardé, R.T., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2009; pp. 174–175. ISBN 9780123741448. [Google Scholar]
- Wearing, C.H.; Hansen, J.D.; Whyte, C.; Miller, C.E.; Brown, J. The potential for spread of codling moth (Lepidoptera: Tortricidae) via commercial sweet cherry fruit: A critical review and risk assessment. Crop Prot. 2001, 20, 465–488. [Google Scholar] [CrossRef] [Green Version]
- Bosch, D.; Avilla, J.; Musleh, S.; Rodríguez, M.A. Target-site mutations (AChE and kdr), and PSMO activity in codling moth (Cydia pomonella (L.) (Lepidoptera: Tortricidae)) populations from Spain. Pestic. Biochem. Physiol. 2018, 146, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, M.A.; Bosch, D.; Avilla, J. Resistance of Spanish codling moth (Cydia pomonella) populations to insecticides and activity of detoxifying enzymatic systems. Entomol. Exp. Appl. 2011, 138, 184–192. [Google Scholar] [CrossRef] [Green Version]
- Sauphanor, B.; Bouvier, J.C.; Brosse, V. Effect of an ecdysteroid agonist, tebufenozide, on the completion of diapause in susceptible and resistant strains of the codling moth, Cydia pomonella. Entomol. Exp. Appl. 1999, 90, 157–165. [Google Scholar] [CrossRef]
- Lacey, L.A.; Unruh, T.R. Biological control of codling moth (Cydia Pomonella, Lepidoptera: Tortricidae) and its role in integrated pest management, with emphasis on entomopathogens. Vedalia 2005, 12, 33–60. [Google Scholar]
- Bezemer, T.M.; Harvey, J.A.; Mills, N.J. Influence of adult nutrition on the relationship between body size and reproductive parameters in a parasitoid wasp. Ecol. Entomol. 2005, 30, 571–580. [Google Scholar] [CrossRef]
- Devotto, L.; del Valle, C.; Ceballos, R.; Gerding, M. Biology of Mastrus ridibundus (Gravenhorst), a potential biological control agent for area-wide management of Cydia pomonella (Linneaus) (Lepidoptera: Tortricidae). J. Appl. Entomol. 2010, 134, 243–250. [Google Scholar] [CrossRef]
- Mills, N. Selecting effective parasitoids for biological control introductions: Codling moth as a case study. Biol. Control 2005, 34, 274–282. [Google Scholar] [CrossRef]
- Retamal, R.; Zaviezo, T.; Malausa, T.; Fauvergue, X.; Le Goff, I.; Toleubayev, K. Genetic analyses and occurrence of diploid males in field and laboratory populations of Mastrus ridens (Hymenoptera: Ichneumonidae), a parasitoid of the codling moth. Biol. Control 2016, 101, 69–77. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Xu, R.; Kuang, R.; Sun, R. Phototactic behaviour of the parasitoid Encarsia formosa (Hymenoptera: Aphelinidae). Biocontrol Sci. Technol. 2016, 26, 250–262. [Google Scholar] [CrossRef]
- Schirmer, S.T.; Sengonca, C.E.; Blaeser, P.E. Influence of abiotic factors on some biological and ecological characteristics of the aphid parasitoid Aphelinus asychis (Hymenoptera: Aphelinidae) parasitizing Aphis gossypii (Sternorrhyncha: Aphididae). Eur. J. Entomol. 2008, 105, 121–129. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Liu, X.; Zhou, J.; Dou, H.; Kuang, R. Phototactic behaviour of Pachyneuron aphidis (Hymenoptera: Pteromalidae)–hyperparasitoid of Myzus persicae (Hemiptera: Aphidiae). Biocontrol Sci. Technol. 2014, 24, 1469–1480. [Google Scholar] [CrossRef]
- Peitsch, D.; Fietz, A.; Hertel, H.; de Souza, J.; Ventura, D.F.; Menzel, R. The spectral input systems of hymenopteran insects and their receptor-based colour vision. J. Comp. Physiol. A 1992, 170, 23–40. [Google Scholar] [CrossRef]
- Jumean, Z.; Unruh, T.; Gries, R.; Gries, G. Mastrus ridibundus parasitoids eavesdrop on cocoon-spinning codling moth, Cydia pomonella, larvae. Naturwissenschaften 2005, 92, 20–25. [Google Scholar] [CrossRef]
- Sandanayaka, W.R.M.; Chhagan, A.; Page-Weir, N.E.M.; Charles, J.G. Colony optimisation of Mastrus ridens (Hymenoptera: Ichneumonidae), a potential biological control agent of codling moth in New Zealand. N. Z. Plant Prot. 2011, 64, 227–234. [Google Scholar] [CrossRef]
- Charles, J.G.; Sandanayaka, W.R.M.; Chhagan, A.; Page-Weir, N.E.M. Host selection behaviour in Mastrus ridens, a gregarious ectoparasitoid of codling moth, Cydia pomonella. BioControl 2013, 58, 493–503. [Google Scholar] [CrossRef]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Mächler, M.; Bolker, B.M. glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. R Found. Stat. Comput.: Vienna, Austria, 2019. Available online: https//www.r-project.org (accessed on 30 June 2020).
- Salt, G. Experimental studies in insect parasitism. III. Host selection. Proc. R. Soc. B Biol. Sci. 1935, 117, 413–435. [Google Scholar] [CrossRef]
- Paine, T.D. Application of chemical cues in arthropod pest management for forest trees. In Chemical Ecology of Insect Parasitoids; Wajnberg, E., Colazza, S., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2013; pp. 282–295. ISBN 9781118409527. [Google Scholar]
- Koul, O.; Dhaliwal, G.S. Predators and parasitoids: An introduction. In Predators and Parasitoids; Koul, O., Dhaliwal, G.S., Eds.; Taylor & Francis: London, UK, 2003; pp. 1–15. ISBN 0203302567. [Google Scholar]
- Vinson, S.B. Host selection by insect parasitoids. Annu. Rev. Entomol. 1976, 21, 109–133. [Google Scholar] [CrossRef]
- Flores-Prado, L.; Niemeyer, H.M. Host Location by ichneumonid parasitoids is associated with nest dimensions of the host bee species. Neotrop. Entomol. 2012, 41, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Sandlan, K. Host location by Coccygomimus turionellae (Hymenoptera: Ichneumonidae). Entomol. Exp. Appl. 1980, 27, 233–245. [Google Scholar] [CrossRef]
- Bueno, E.; Romero, A.; Osorio, I.; Zaviezo, T. Laboratory rearing methods that promote inbreeding have a negative impact on the fitness of Mastrus ridens Horstmann (Hymenoptera: Ichneumonidae), a parasitoid used to control the codling moth. Chil. J. Agric. Res. 2017, 77, 413–419. [Google Scholar] [CrossRef]
- Hougardy, E.; Bezemer, T.M.; Mills, N.J. Effects of host deprivation and egg expenditure on the reproductive capacity of Mastrus ridibundus, an introduced parasitoid for the biological control of codling moth in California. Biol. Control 2005, 33, 96–106. [Google Scholar] [CrossRef]
- Bastías, R.M.; Manfrini, L.; Corelli Grappadelli, L. Exploring the potential use of photo-selective nets for fruit growth regulation in apple. Chil. J. Agric. Res. 2012, 72, 224–231. [Google Scholar] [CrossRef]
- Ilić, Z.S.; Milenković, L.; Šunić, L.; Fallik, E. Effect of coloured shade-nets on plant leaf parameters and tomato fruit quality. J. Sci. Food Agric. 2015, 95, 2660–2667. [Google Scholar] [CrossRef] [PubMed]
- Lobos, G.A.; Retamales, J.B.; Hancock, J.F.; Flore, J.A.; Romero-Bravo, S.; Del Pozo, A. Productivity and fruit quality of Vaccinium corymbosum cv. Elliott under photo-selective shading nets. Sci. Hortic. 2013, 153, 143–149. [Google Scholar] [CrossRef]
- Legarrea, S.; Weintraub, P.G.; Plaza, M.; Viñuela, E.; Fereres, A. Dispersal of aphids, whiteflies and their natural enemies under photoselective nets. BioControl 2012, 57, 523–532. [Google Scholar] [CrossRef] [Green Version]
- Dáder, B.; Plaza, M.; Fereres, A.; Moreno, A. Flight behaviour of vegetable pests and their natural enemies under different ultraviolet-blocking enclosures. Ann. Appl. Biol. 2015, 167, 116–126. [Google Scholar] [CrossRef]
- Cochard, P.; Galstian, T.; Cloutier, C. Light environments differently affect parasitoid wasps and their hosts’ locomotor activity. J. Insect Behav. 2017, 30, 595–611. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PSN Color | Supplier | Specifications | PAR (µmol m−2 s−1) | Shading Factor (%) |
|---|---|---|---|---|
| No PSN (control) | - | - | 7 | 0 |
| Pearl PSN | ChromatiNet®, Polysack Industries, Negev, Israel | HDPE monofilament, mesh size 5 × 2 mm | 6 | 20 |
| Red PSN | ChromatiNet®, Polysack Industries, Negev, Israel | HDPE monofilament, mesh size 5 × 2 mm | 5 | 20 |
| Black SN | ChromatiNet®, Polysack Industries, Negev, Israel | Polyethylene, mesh size 3 × 1 mm | 2 | 80 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yáñez-Díaz, M.-J.; Rodríguez, M.; Musleh, S.; Devotto, L.; Silva, G.; Lucas, E. Photo-Selective Nets and Pest Control: Searching Behavior of the Codling Moth Parasitoid Mastrus ridens (Hymenoptera: Ichneumonidae) under Varying Light Quantity and Quality Conditions. Insects 2021, 12, 582. https://doi.org/10.3390/insects12070582
Yáñez-Díaz M-J, Rodríguez M, Musleh S, Devotto L, Silva G, Lucas E. Photo-Selective Nets and Pest Control: Searching Behavior of the Codling Moth Parasitoid Mastrus ridens (Hymenoptera: Ichneumonidae) under Varying Light Quantity and Quality Conditions. Insects. 2021; 12(7):582. https://doi.org/10.3390/insects12070582
Chicago/Turabian StyleYáñez-Díaz, María-José, Marcela Rodríguez, Selim Musleh, Luis Devotto, Gonzalo Silva, and Eric Lucas. 2021. "Photo-Selective Nets and Pest Control: Searching Behavior of the Codling Moth Parasitoid Mastrus ridens (Hymenoptera: Ichneumonidae) under Varying Light Quantity and Quality Conditions" Insects 12, no. 7: 582. https://doi.org/10.3390/insects12070582
APA StyleYáñez-Díaz, M. -J., Rodríguez, M., Musleh, S., Devotto, L., Silva, G., & Lucas, E. (2021). Photo-Selective Nets and Pest Control: Searching Behavior of the Codling Moth Parasitoid Mastrus ridens (Hymenoptera: Ichneumonidae) under Varying Light Quantity and Quality Conditions. Insects, 12(7), 582. https://doi.org/10.3390/insects12070582